小檗属植物叶绿体基因组序列结构及系统发育分析
2022-02-14李述成郭生虎贝盏临
李述成,郭生虎,贝盏临
• 药材与资源 •
小檗属植物叶绿体基因组序列结构及系统发育分析
李述成1,郭生虎2,贝盏临3*
1. 宁夏农林科学院 宁夏 银川 750002 2. 宁夏农林科学院 农业生物技术研究中心 宁夏 银川 750002 3. 北方民族大学 生物科学与工程学院 宁夏 银川 750021
了解小檗属植物叶绿体基因组成及结构特征,为小檗属的系统发育及基因组进化研究提供参考。获取小檗属9个物种的叶绿体基因组,利用生物信息学方法比较叶绿体基因组间的结构特征与变异程度,并以小檗科的桃儿七属3个物种的叶绿体基因组为外类群分析了小檗属的系统发育关系。小檗属9个种的叶绿体基因组均为双链环形结构,均包含1个大单拷贝区(large single-copy,LSC)、1个小单拷贝区(small single-copy,SSC)以及2个反向重复区(IRa和IRb)。叶绿体基因组大小差异较小,最大差异为1984 bp。基因排列顺序基本一致,各基因数量相对保守,其中核糖体RNA(rRNA)数量最为保守,均为8个。此外,小檗属叶绿体基因组在序列长度、基因组成以及GC含量等方面相对保守,但4个边界存在明显的多样性。小檗属叶绿体基因组中非基因编码区存在较大差异,变异程度较高,而基因编码区差异较小,具有较高的保守性。在叶绿体基因组4个部分中,SSC区的变异程度最高,IRa区的变异程度最低。小檗属叶绿体基因组中包括、、、、、、、、、、和12个含内含子的蛋白编码基因。大部分基因的编码区长度较为保守,但在内含子长度或编码区长度均存在少量改变。十大功劳组是最先从小檗属中分离出来,是小檗属的原始组。在十大功劳组中阿里山十大功劳和安坪十大功劳与其他小檗属物种存在较远的亲缘关系。而其余小檗属物种可分为2支,威宁小檗为1支;黄芦木和朝鲜小檗为1支。小檗属叶绿体基因组具有较高的保守性,其基因排列顺序基本一致,未检测到大规模的倒位或基因重排,但其IR区与单拷贝区(single-copy,SC)边界存在明显的多样性。基于叶绿体基因组构建的系统发育树在一定程度上可以揭示小檗属物种的系统发育关系。
小檗属;叶绿体基因组;变异;进化;系统发育
叶绿体(chloroplast)作为质体(plastid)中的一种类型,其基因组大小多为150~200 kb,且结构较为保守[1]。叶绿体基因组大多可划分为4个边界区:一个大单拷贝(large single-copy,LSC)和一个小单拷贝(small single-copy,SSC)区域以及通过基因组中的连接位点(junction sites,JS)连成的2个相同的反向重复序列区IR(inverted repeat,IRa和IRb)[2],其编码约有120~130个基因,在基因组中只有一小部分由非编码基因区间组成,使得功能基因具有相当大的进化信号,作为高纯化来抵消不具关键进化重要性的部分基因的压力[3]。叶绿体细胞器通过有氧呼吸和光合作用确保了在真核细胞中发挥的关键作用。而在细胞起源于独立的复杂性的内共生事件中,叶绿体细胞器一直都能保持自己的基因组不变化,是一个非常重要的特性[4-5]。然而,最近的研究表明,叶绿体基因组的基因可能不像以前认为的那样紧密相连,也可能会经受不同的进化力量影响[6]。使得叶绿体基因作为种分类、系统发育、种起源以及避难所等研究中的依据,而被广泛应用。
小檗属L.是小檗科(Leguminosae)中最大的一个属[7],该属大约由500种植物组成[8]。依据植物地理分布,Schneider将小檗属植物分为2组,中隔小檗属和澳大利亚小檗属[9]。中隔小檗属(旧大陆)由约300种组成,主要分布在欧亚大陆,但延伸至北美(2种)和北非(4种)。澳大利亚类群(新大陆)包含约200种物种,其中大部分分布在南美洲,少数分布在中美洲[10]。在中亚大多数物种则多集中在喜马拉雅地区,从巴基斯坦延伸到中国西部,再延伸到中国中部和南部,其中在中国区域的约有250种物种。以上这些地理分组都是基于物种形态特征进行的分组[9]。小檗属植物大多含有生物碱,其中最主要的生物碱是“小檗碱”[11],这些生物碱对霍乱、腹泻、阿米巴病、疟疾和利什曼病具有活性[12],具有很高的药用价值。此外,一些小檗属植物能被当作高价值的木材染料,另一些则可被当作食用的浆果。在中国民间医药中小檗属植物也常被使用,但该属物种在形态学特征识别较为困难,因此在使用时需要通过多种方法弄清小檗属物种类别。比如近期有研究报道小檗属植物中国特有中药威宁小檗L.[13]是通过叶绿体全基因组的分析方法,对该物种进行了分类。此外,来自阿根廷图库马地区的小檗属一新种L.,也是通过其DNA序列进行了系统发育位置的评估,最后被证实为南美洲小檗属其他物种的一个外类群[14]。评估物种系统发育位置也可以通过以其叶绿体基因为研究对象进行界定与分析。因此,形态学特征识别较为困难的小檗属物种,也可以利用其绿体基因组序列分析系统发育和DNA条形码作为依据鉴定该属植物。
本项研究对小檗属9个物种的叶绿体基因组结构和组成进行了界定与分析,并通过该属9个种的叶绿体基因组进行系统进化树的构建,为小檗属物种的鉴定以及进一步研究小檗属的系统发育进化提供一定的参考。
1 材料与方法
1.1 数据收集
通过美国国家生物信息中心(The National Center for Biotechnology Information,NCBI)数据库(https://www.ncbi.nlm.nih.gov/)检索小檗科的小檗属物种和桃儿七属物种的叶绿体全基因组序列信息。共检索到小檗属9个种信息,桃儿七属植物3个种信息。下载检索到的物种的叶绿体基因组序列、名称、基因组登录号,见表1。
表1 小檗属和桃儿七属物种的叶绿体基因组Genbank登记号
Table 1 Chloroplast Genbank accession numbers of species in Berberis and Sinopodophyllum
物种GenBank登记号文献 阿里山十大功劳B. oiwakensis HayataMN73522115 安坪十大功劳B. ganpinensis H.Lév.MN41730716 阔叶十大功劳B. bealei Fortune [Mahonia bealei (Fort.) Carr.]MH79530817 长柱十大功劳B. duclouxiana Gagnep. (Mahonia duclouxiana) isolate RC602MZ08677018 十大功劳B. fortunei (Lindl.) FeddeMH79530719 黄芦木B. amurensis Rupr var. quelpaertensis Nakai KM05737620 朝鲜小檗B. koreana PalibKM05737521 威宁小檗B. weiningensis T. S. YingMW01836322 具芒小檗B. aristata DC.MN74630823 桃儿七Sinopodophyllum hexandrum Roylevoucher SHEXA20200817MT98239624 桃儿七S. hexandrum RoyleMG59304824 桃儿七S. hexandrum Roylevoucher SHEX20150512KR77999424
1.2 方法
采用Geneious10.2.2软件[25]统计了小檗属9个物种的叶绿体基因组序列的4个边界(SSC、LSC、IRa和IRb区域)长度和基因数目类型、GC含量等信息。采用R语言4.1.1和Chloroplot软件[26]生成小檗属9个物种叶绿体基因组图谱,并进行了手动检查。采用R语言4.1.1和IRscope软件[27]对小檗属9个物种4个边界的收缩与扩张进行可视化分析。通过Python3.10.1进行文件注释后,采用mVISTA软件[28],并选用检测基因重排和倒位的全局比对模式(Shuffle-LAGAN),以十大功劳作为参考基因组,对小檗属另外8个物种叶绿体全基因组序列同源性进行了比较研究。采用Geneious10.2.2软件中Mauve多重基因组比对法对小檗属9个物种的叶绿体基因组进行共线性比较。
1.3 系统发育分析
本研究采用BioEdit 7.2.5软件[29]比对了9种小檗属物种,并基于IQ-tree中RaxML法(random axelerated maximum likelikhood)[30],以桃儿七属3个物种为外类群,利用自助抽样法(bootstrap method)进行重复抽样检验,并重复检验1000次,构建出系统发育树。构建系统发育树所选物种的详细信息见表1。
2 结果与分析
2.1 小檗属叶绿体基因组结构的基本特征
小檗属9个物种叶绿体基因组图谱(图1)。小檗属9个物种叶绿体基因组均为双链环形结构,均具有4个边界区(1个SSC、1个LSC、1个IRa和1个IRb),9个物种的叶绿体基因组范围在164 774~166 758 bp,其中最大的是朝鲜小檗,最小的是具芒小檗,最大与最小差为1984 bp。SSC长为18 563~18 773 bp,LSC长为73 286~73 774 bp,2个相同的反向重复序列IRa和IRb长为36 375~37 152 bp。小檗属9个物种叶绿体基因组的GC含量均为38%,无差异(图2)。
小檗属9个物种叶绿体基因组比较结果(图2)。小檗属9个物种叶绿体基因组较为保守,总基因数为95~104个,安坪十大功劳、长柱十大功劳具芒小檗的总基因数最多,黄芦木和朝鲜小檗的总基因数最少。rRNA(核糖体RNA)的数量均为8个,最为恒定保守。tRNA(转运RNA)的数量为34~38个,其中安坪十大功劳为34个。蛋白编码基因数为49~62个,其中安坪十大功劳的蛋白编码基因为62个,黄芦木和朝鲜小檗的蛋白编码基因均为49个。
2.2 小檗属叶绿体基因组IR与SC边界
小檗属叶绿体基因组虽然在序列长度、基因组成以及GC含量等方面相对稳定保守,但4个边界区的过渡区域却存在有多样性(图3)。小檗属9个物种的IRb-LSC边界,其中十大功劳、长柱十大功劳、阔叶十大功劳、安坪十大功劳、具芒小檗、威宁小檗均位于B的编码区,而黄芦木和朝鲜小檗则基因缺失。除了威宁小檗、黄芦木和朝鲜小檗外,小檗属其他种IRb-SSC边界均在基因间区内或的编码区。小檗属9个种的IRa-SSC边界,除了黄芦木、朝鲜小檗和威宁小檗外,其他种均位于基因内。阿里山十大功劳安坪十大功劳阔叶十大功劳、长柱十大功劳十大功劳、具芒小檗的IRa区均有R基因。此外,小檗属9个物种的IRa-LSC边界具有较高的保守性,其边界均位于B与tRNA-His(H)基因的基因间区。P的编码均在LSC区,F的编码均在SSC区,N的编码均在IRa区。
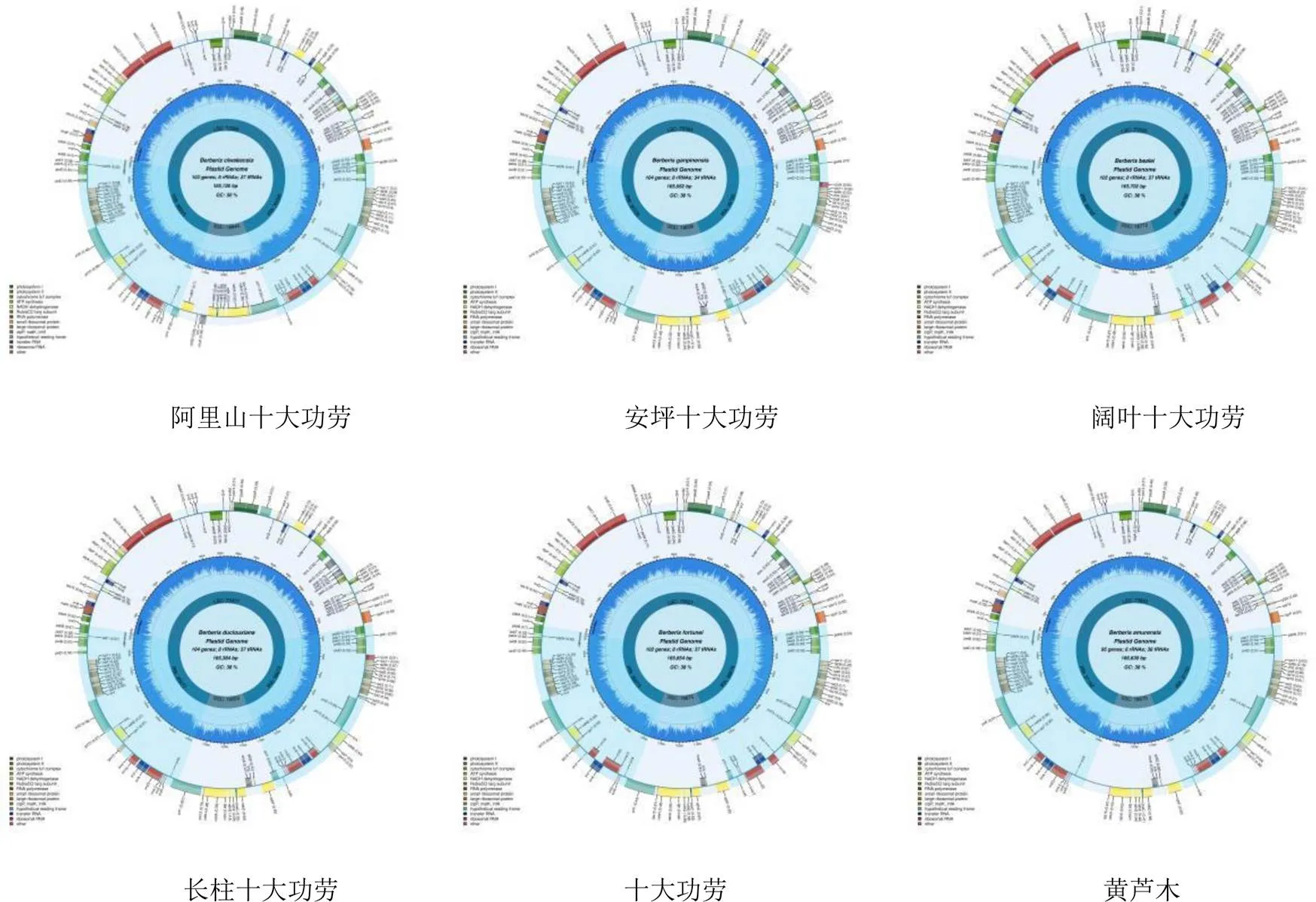

图2 小檗属9个物种的叶绿体基因组的基本特征

图3 小檗属9个物种叶绿体基因组的IR与SC边界比较
2.3 小檗属叶绿体基因组序列变异分析
本研究采用mVISTA软件[28],并选用检测基因重排和倒位的全局比对模式(Shuffle-LAGAN),以阿里山十大功劳作为参考基因组,对小檗属另外8个物种叶绿体全基因组序列同源性进行了比较研究(图4)。结果表明,小檗属9个物种叶绿体基因组4个部分排列顺序较为一致、保守性较高,在非基因编码区存在有较明显的差异,变异程度较高,其中LSC区的变异程度最高。而在基因编码区差异不明显,在IRa区的变异程度最低,具有较高的保守性。小檗属叶绿体基因组中在、、J、D、、、、、、、、、以及B等基因的编码区存在显著差异。
2.4 小檗属叶绿体基因组内含子分析
小檗属9个物种的叶绿体基因组中内含子编码序列(coding sequences,CDS)长度和基因(gene length)全长见表2。小檗属9个物种的叶绿体基因组中共检测到12个含内含子的蛋白编码基因:、、、、、、、、、、和。在这12个含内含子的蛋白编码基因中,内含子长度或编码序列长度等因素均会直接影响到整个基因长度的变化。在小檗属9个物种中基因的编码序列长度一致,而基因长度存在少量改变。基因的编码序列在安坪十大功劳、黄芦木朝鲜小檗中没有重复,而在阿里山十大功劳、阔叶十大功劳、长柱十大功劳、十大功劳、威宁小檗、具芒小檗等其它6个种内则有重复。此外5种基因的编码也没有重复,而在4种基因的编码则均有重复。除了基因外,其它各基因在内含子长度或编码序列长度均存在少量改变。
2.5 小檗属的叶绿体基因组共线性分析
采用Mauve多重基因组比对法检测小檗属9个种的叶绿体基因组的重排和共线性(图5),通过多重基因组比对法检测出9个种的叶绿体基因组之间有3个局部共线块(locally collinear block,LCB),这表明小檗属9个种之间的基因组具有高度的相似性。小檗属9个种的叶绿体全基因组序列的比对显示,小檗属9个种的叶绿体基因组之间没有重排或倒置,然在区域(73 000~110 000;130 000~165 000)中观察到常见的突变,其特征是在对齐的叶绿体基因组中基因序列的高度变异。

图4 小檗属9种叶绿体基因序列比对分析
2.6 系统发育分析
根据本研究中9个小檗属物种的叶绿体基因组,并另选择了来自小檗科桃儿七属3个物种的叶绿体基因组为外类群构建系统发育树(图6)。在系统发育树中,可确定为不包括外类群物种的2个主要分支:十大功劳组与具芒小檗组。十大功劳组为一个单系,有很好的支持率(100%),支系中的支持率大于90%;而具芒小檗组为另外一个单系,有很好的支持率(100%),支系中的支持率不低于100%,具芒小檗与威宁小檗、朝鲜小檗、黄芦木具有密切的关系。黄芦木和朝鲜小檗的亲缘关系最密切。结果表明,叶绿体基因组构建的系统发育树可用于小檗属9个物种植物的鉴定。
3 讨论
质体基因组(plastome)是重建绿色植物系统发育[31]最重要的数据来源,最近的核基因系统基因组分析普遍也支持以前基于质体的假设[32-33]。作为质体类型之一的叶绿体,其结构与序列具有高度保守性[34-35]。反向重复结构IR区在高等植物叶绿体基因组较为常见的区,IR区的长短常会影响到叶绿体基因组内同源序列线性顺序的重排[36]。因此,IR区的长度在维持叶绿体基因组的稳定性中起到重要作用[37-38]。
表2 小檗属9种植物的叶绿体基因组中内含子的编码序列长度和基因全长
Table 2 CDS length and gene length of intron of chloroplast genomes in nine species of Berberis
内含子基因阿里山十大功劳安坪十大功劳阔叶十大功劳长柱十大功劳十大功劳黄芦木朝鲜小檗威宁小檗具芒小檗 rps12CDS length372/372372372/372372/372372/372372372372/372372/372 Gene length908/908372980/980980/980980/980908908980/980980/980 rps16CDS length237237237237237237237237237 Gene length113211321134113311311133113311321133 rpl2CDS length837/993816/816852/855816/456852/855816/816816/816810/810852/855 Gene length1493/14931475/14751475/14751475/11151475/14751475/14751475/14751469/14691475/1475 rpl16CDS length408/408420/408408/408408/408408/408408/408408/408411/408408/408 Gene length1239/12391239/12391238/12381237/12371237/12371234/12341234/1234411/12381239/1239 rpoC1CDS length205520522046204320462064206420762046 Gene length279927992799279927992824282428242804 petBCDS length648/660654/648660/660648/648660/660660/660660/660648/660660/660 Gene length1417/14171417/14171417/14171417/14171424/14241424/14241424/14241424/14241424/1424 petDCDS length546/54660/504504/504546/504504/504504/504504/504546/504507/507 Gene length546/546693/11871187/1187546/11871187/11871187/11871187/1187546/11871190/1190 ndhBCDS length1536/16381536/16381626/16291536/15361626/16291638/15361638/15361536/15361626/1629 Gene length2228/22282228/22282228/22282228/22282228/22282227/22272227/22272227/22272227/2227 ndhACDS length110710951098109510981095109510951098 Gene length213921362136214121412135213321432143 atpFCDS length570555558555558570570555558 Gene length128012801279128012791280128012801277 clpPCDS length642612606642606606606606606 Gene length209620762074210720792080207620702065 ycf3CDS length510507510510510510510510510 Gene length198119801979197919801980198119821980

图5 小檗属9个物种的叶绿体全基因组比对

图6 小檗属叶绿体基因组序列构建的系统发育树
本研究利用小檗属9个种的叶绿体基因组,对小檗属内种的叶绿体基因组进行比较分析,9个物种叶绿体基因组均为双链环形结构,均具有4个边界(1个SSC、1个LSC、1个IRa和1个IRb),9个物种的叶绿体基因组长度在164 kp左右,最大和最小差为1984 bp。SSC长度在18 kp左右,LSC长度在73 kp左右,而2个相同的反向重复序列IRa和IRb的长度范围在36~37 kp。因此小檗属种叶绿体基因组较为恒定,具有较高的保守性。虽然大多数高等被子植物叶绿体基因组IR区中的基因突变率常低于位于2个长短单拷贝区中的基因[39],但是叶绿体基因组中2个反向重复序列区与长、短单拷贝区的边界会存在扩张与收缩,也会像基因和内含子的缺失一样影响到叶绿体基因组从而显示出高的突变性。
小檗属植物叶绿体基因组具有较高的保守性,所研究的小檗属9个物种的叶绿体基因组排列顺序基本一致,未检测到大规模的倒位或基因重排,但根据研究结果发现小檗属9个物种的叶绿体基因组中的IR与SC边界显示也有存在明显的扩张与收缩。此外,小檗属叶绿体基因组中有内含子的蛋白编码基因12个,基因长度变异主要是因为内含子长度或编码序列长度的变化,编码区差异性较小基因保守性则会较高。小檗科自被建立为属以来,其分类以及系统发育研究备受关注。前人对小檗属的系统发育研究多集中在属间系统发育关系方面,对于小檗属内的系统发育分析仍有待进一步研究。本研究通过构建小檗属物种系统发育树,通过支持率可将小檗属种划分为2个主要分支:十大功劳组与具芒小檗组。并且2个主要分支及其直系分支的支持率均不低于92%。因此,叶绿体全基因组序列分析在一定程度上可以支持小檗属的系统发育关系,可为进一步的系统发育与进化研究提供依据。
利益冲突 所有作者均声明不存在利益冲突
[1] Neuhaus H E, Emes M J. Nonphotosynthetic metabolism in plastids [J]., 2000, 51: 111-140.
[2] Shinozaki K, Ohme M, Tanaka M,. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression [J]., 1986, 5(9): 2043-2049.
[3] Wu C S, Lai Y T, Lin C P,. Evolution of reduced and compact chloroplast genomes (cpDNAs) in gnetophytes: Selection toward a lower-cost strategy [J]., 2009, 52(1): 115-124.
[4] Ponce-Toledo R I, Deschamps P, López-García P,. An early-branching freshwater cyanobacterium at the origin of plastids [J]., 2017, 27(3): 386-391.
[5] Martijn J, Vosseberg J, Guy L,. Deep mitochondrial origin outside the sampled alphaproteobacteria [J]., 2018, 557(7703): 101-105.
[6] Gonçalves D J P, Simpson B B, Ortiz E M,. Incongruence between gene trees and species trees and phylogenetic signal variation in plastid genes [J]., 2019, 138: 219-232.
[7] Kim Y D, Kim S H, Landrum L R. Taxonomic and phytogeographic implications from ITS phylogeny in(Berberidaceae) [J]., 2004, 117(3): 175-182.
[8] Ahrendt L W A.and Mahonia A taxonomic revision [J]., 1961, 57(369): 1-410.
[9] Schneider C. Die gattung Berberis (Euberberis): vorarbeiten für eine monographie [M].:, 1905: 33-48.
[10] Landrum L R. Revision of(Berberidaceae) in Chile and adjacent southern[J]., 1999, 86(4): 793.
[11] Bottini M C J, De Bustos A, Sanso A M,. Relationships in Patagonian species of(Berberidaceae) based on the characterization of rDNA internal transcribed spacer sequences [J]., 2007, 153(3): 321-328.
[12] Singh P B, Ambasta S S, Tripathi V N,. Blunt renal injury: An experience of 30 cases [J]., 1986, 17(4): 228-229.
[13] Feng T, Xiao Q Y, Li W J,. The complete chloroplast genome of(Berberidaceae), an endemic and traditional Chinese medicinal herb [J]., 2021, 6(3): 1175-1177.
[14] Gori M, Biricolti S, Pedrazzani S,.OR Dantur, S. Radice, E. Giordani, A. apini sp. nov. (Berberidaceae): A new species [J]., 2021, 68(5): 1799-1808.
[15] Icon P l.[M].():, 1916, 6: 1.
[16] Gard CFortune [J].:, 1850, 4: 212.
[17] Bull S.Gagnep [M].:, 1908, 55: 87.
[18] John L.Lindl [J].. 1846, 1: 231.
[19] Bull B.Rupr in Bull [J]., 1857, 15: 260.
[20] Trudy I S.Palib. [J]., 1899, 17: 22.
[21] Ying T S.[J]., 1999, 37(4): 326.
[22] Parmar C, Kaushai M K.DC. [J]., 1821, 2: 8.
[23] Royle J F.Royle in Illustr [J]., 1834, 2(1): 64.
[24] Kearse M, Moir R, Wilson A,. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data [J]., 2012, 28(12): 1647-1649.
[25] Zheng S Y, Poczai P, Hyvönen J,. Chloroplot: An online program for the versatile plotting of organelle genomes [J]., 2020, 11: 576124.
[26] Amiryousefi A, Hyvönen J, Poczai P. IRscope: An online program to visualize the junction sites of chloroplast genomes [J]., 2018, 34(17): 3030-3031.
[27] Frazer K A, Pachter L, Poliakov A,. VISTA: computational tools for comparative genomics [J]., 2004, 32 (Web Server issue): W273-W279.
[28] Hall T, Biosciences I, Carlsbad C. BioEdit: An important software for molecular biology [J]., 2011, 2(1): 60-61.
[29] Nguyen L T, Schmidt H A, von Haeseler A,. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies [J]., 2015, 32(1): 268-274.
[30] Davis C C, Xi Z X, Mathews S. Plastid phylogenomics and green plant phylogeny: Almost full circle but not quite there [J]., 2014, 12: 11.
[31] Zeng L P, Zhang Q, Sun R R,. Resolution of deep angiosperm phylogeny using conserved nuclear genes and estimates of early divergence times [J]., 2014, 5: 4956.
[32] Wickett N J, Mirarab S, Nguyen N,. Phylotranscriptomic analysis of the origin and early diversification of land plants [J]., 2014, 111(45): E4859-E4868.
[33] Chumley T W, Palmer J D, Mower J P,. The complete chloroplast genome sequence ofx: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants [J]., 2006, 23(11): 2175-2190.
[34] Serrano M, Wang B J, Aryal B,. Export of salicylic acid from the chloroplast requires the multidrug and toxin extrusion-like transporter EDS5 [J]., 2013, 162(4): 1815-1821.
[35] Palmer J D, Thompson W F. Rearrangements in the chloroplast genomes of mung bean and pea [J]., 1981, 78(9): 5533-5537.
[36] Guisinger M M, Kuehl J V, Boore J L,. Extreme reconfiguration of plastid genomes in the angiosperm family Geraniaceae: Rearrangements, repeats, andusage [J]., 2011, 28(1): 583-600.
[37] Hirao T, Watanabe A, Kurita M,. Complete nucleotide sequence of theD. Don. chloroplast genome and comparative chloroplast genomics: Diversified genomic structure of coniferous species [J]., 2008, 8: 70.
[38] Maier R M, Neckermann K, Igloi G L,. Complete sequence of the maize chloroplast genome: Gene content, hotspots of divergence and fine tuning of genetic information by transcript editing [J]., 1995, 251(5): 614-628.
[39] Dempewolf H, Kane N C, Ostevik K L,. Establishing genomic tools and resources for(L.f.) Cass.—the development of a library of expressed sequence tags, microsatellite loci, and the sequencing of its chloroplast genome [J]., 2010, 10(6): 1048-1058.
Chloroplast genome sequences and phylogenetic analysis ofgenus
LI Shu-cheng1, GUO Sheng-hu2, BEI Zhan-lin3
1. NingXia Academy of Agricultural and Forestry Sciences, Yinchuan 750002, China 2. Agri-Biotechnology Research Centre, NingXia Academy of Agricultural and Forestry Sciences, Yinchuan 750002, China 3. School of Biological Science and Engineering, North Minzu University, Yinchuan 750021, China
To understand the chloroplast gene composition and structural characteristics of, so as to provide reference for the phylogeny and genome evolution of.The chloroplast genomes of nine species ofwere obtained. The structural characteristics and variation degree of chloroplast genomes were compared by bioinformatics methods. The phylogenetic relationship ofwas analyzed by taking the chloroplast genomes of three species ofas an outgroup.The chloroplast genomes of nine species ofhad a double stranded ring structure, including a long single copy region (LSC), a short single copy region (SSC) and two reverse repeats (IRa and IRb). The difference of chloroplast genome size was small, and the maximum difference was 1984 bp. The sequence of genes was basically the same, and the number of genes was relatively conservative, among which the number of ribosomal RNA (rRNA) was the most conservative, with eight genes each. In addition, the chloroplast genome ofwas relatively conservative in sequence length, gene composition and GC content, but there was obvious diversity in the four boundaries. The non gene coding regions in the chloroplast genome ofhad great differences and high degree of variation, while the gene coding regions had small differences and high conservatism. Among the four parts of chloroplast genome, the variation degree of SSC region was the highest and that of IRA region was the lowest. The chloroplast genome ofincluded 12 protein coding genes containing intron such as,,,,,,, ndhB,,,and. The length of coding region of most genes was conservative, but there were a few changes in intron length or coding region length.group was the first isolated from, which was the original group of. In theformation,andwere far related to otherspecies. The other species ofcan be divided into two branches, andwas one branch;andwere one branch.The chloroplast genome ofis highly conservative, and its gene sequence is basically the same. No large-scale inversion or gene rearrangement is detected, but there is obvious diversity at the boundary between IR region and single copy region (SC). The phylogenetic tree based on chloroplast genome can reveal the phylogenetic relationship ofspecies to a certain extent.
L.;chloroplast genome;variation;evolution;phylogeny
R286.12
A
0253 - 2670(2022)03 - 0818 - 09
10.7501/j.issn.0253-2670.2022.03.022
2021-10-03
国家自然科学基金资助项目(31860086);宁夏自然科学基金资助项目(2021AAC03190);农业高质量发展和生态保护科技创新示范课题(NGSB-2021-9-01);自主科研项目(NKYZZ-J-19-07)
李述成(1972—),男,硕士研究生,研究方向为农业经济管理。Tel: 13037963679 E-mail: nxnlkxy1166@163.com
贝盏临(1975—),男,硕士,副研究员,研究方向为药用植物资源。Tel: (0951)2067508 E-mail: realpal0047@163.com
[责任编辑 时圣明]
