lncRNA MIAT通过miR-203调控NPAS4促进胃癌细胞增殖、侵袭、迁移的机制研究
2022-02-11刘丹吴瑾周红凤刘东辉胡晓薇
刘丹,吴瑾,周红凤,刘东辉,胡晓薇
【提要】目的 本研究通过体外培养胃癌细胞HGC-27,探究lncRNA MIAT能否通过靶向miR-203调控神经元PAS结构域蛋白4(NPAS4)促进胃癌细胞增殖、侵袭、迁移。方法 体外培养HGC-27与GES-1细胞,qRT-PCR法检测各细胞中lncRNA MIAT、miR-203、NAPS4 mRNA表达。双荧光素酶报告基因实验验证lncRNA MIAT与miR-203、miR-203与NPAS4之间的靶向关系。将对数生长期HGC-27细胞分组并进行转染:对照组、si-NC组(转染si-NC)、si-MIAT组(转染si-MIAT)、miR-NC组(转染miR-NC)、miR-203组(转染miR-203)、anti-miR-NC组(转染anti-miR-NC)、anti-miR-203组(转染anti-miR-203)、si-MIAT+anti-miR-NC组(转染si-MIAT+anti-miR-NC)、si-MIAT+anti-miR-203组(转染si-MIAT+anti-miR-203)、pc-NC组(转染pc-NC)、pc-NAPS4组(转染pc-NAPS4)、pc-NAPS4+miR-NC组(转染pc-NAPS4+miR-NC)、pc-NAPS4+miR-203组(转染pc-NAPS4+miR-203),qRT-PCR法检测lncRNA MIAT、miR-203、NPAS4 mRNA表达;MTT法检测细胞增殖活力;Transwell实验检测细胞迁移与侵袭能力;Western blot法检测MMP-3、MMP-9、CyclinD1、NPAS4蛋白表达。结果 与GES-1细胞比较,胃癌细胞HGC-27中lncRNA MIAT、NAPS4 mRNA表达显著升高,miR-203表达显著降低(P<0.05)。双荧光素酶报告基因实验证实,lncRNA MIAT能够靶向miR-203调控NAPS4(P<0.05)。沉默lncRNA MIAT可显著降低细胞活力以及迁移与侵袭能力,下调MMP-3、MMP-9、CyclinD1蛋白表达,降低NAPS4 mRNA和蛋白表达(P<0.05);过表达miR-203可显著降低NAPS4 mRNA和蛋白表达,降低细胞活力以及迁移与侵袭能力,下调MMP-3、MMP-9、CyclinD1蛋白表达(P<0.05);而抑制miR-203表达则与miR-203过表达作用相反,且抑制miR-203表达可显著逆转沉默lncRNA MIAT对细胞活力以及迁移与侵袭能力的抑制作用(P<0.05);过表达NAPS4可显著促进HGC-27细胞增殖、迁移与侵袭,上调MMP-3、MMP-9、CyclinD1蛋白表达(P<0.05),同时其对细胞恶性行为的促进作用可被miR-203过表达削弱(P<0.05)。结论 沉默lncRNA MIAT可通过靶向miR-203调控NAPS4表达,减弱胃癌细胞增殖、侵袭、迁移等细胞生物学行为过程。
胃癌是一种常见的消化系统恶性肿瘤,其发病率与致死率均较高,严重威胁患者生命健康,增加家庭与社会负担[1]。我国胃癌的发病率远高于其他国家,尽管胃癌的治疗取得了快速进展,但由于放、化疗等治疗手段的副作用,导致患者预后极差,生存率仍然很低[2-3]。研究发现,胃癌细胞发生恶性增殖、侵袭与转移是造成治疗失败、患者死亡的主要原因之一[4],但其潜在机制尚未发现。因此,探究胃癌细胞恶性行为学的发生机制对于胃癌患者治疗与预后具有非常重要的意义。长链非编码RNA(long non-coding RNA, lncRNA)可在多种肿瘤细胞中表达异常,并参与调控肿瘤细胞增殖、凋亡、侵袭与迁移等恶性生物学行为,与肿瘤的发生与发展存在密切联系[5-6]。据研究报道,多种lncRNA在胃癌中异常表达,调控细胞生物学过程[7-8],可能与胃癌的发生与发展有关。研究发现,血清外泌体来源lncRNA MIAT在胃癌患者体内高表达,可作为胃癌诊断与预后生物标志物[9]。另外,lncRNA MIAT在非小细胞肺癌细胞中呈高表达,敲低其表达能显著减少细胞增殖,抑制上皮间质转化等细胞过程[10]。近年来,微小RNA(microRNA, miRNA)在肿瘤中异常表达,参与肿瘤进展,且多种miRNA参与胃癌的发生、发展[11],其中miR-203在胃癌中低表达,影响胃癌细胞分化、转移,是胃癌患者预后的独立危险因素之一[12]。神经元PAS结构域蛋白4(neuronal Per-ArntSim domain protein 4,NPAS4)是一种与神经元兴奋相关的转录因子,几乎仅表达于神经系统,参与调节学习行为与记忆形成[13]。有研究称,NPAS4过表达对LPS诱导的细胞凋亡与炎症反应具有密切联系[14],但其在胃癌细胞表达情况以及对胃癌细胞行为学的影响尚不清楚。本研究通过体外培养胃癌细胞HGC-27,探究lncRNA MIAT能否通过靶向miR-203调控NPAS4促进胃癌细胞增殖、侵袭、迁移。
1 材料
1.1 细胞株与主要试剂
胃癌细胞HGC-27、正常胃黏膜上皮细胞GES-1均购自中国科学院细胞库;胎牛血清(abs9*)购自爱必信(上海)生物科技有限公司;青链双抗溶液(03-031-5B)购自以色列Bioind公司;RPMI1640培养基(PM150110)购自武汉普诺赛生命科技有限公司;TRIzol试剂(SH-2366)购自北京凯诗源生物科技有限公司;si-MIAT、miR-203模拟物(miR-203)、anti-miR-203、pc-NAPS4及其阴性对照物均购自上海吉玛制药有限公司;MTT细胞增殖检测试剂盒(M8180-1)购自北京索莱宝科技有限公司;Transwell小室(3413)购自美国Corning公司;Matrigel基底胶(SJ-JC15426)购自上海机纯实业有限公司;MMP-3抗体(ab52915)、MMP-9抗体(ab76003)、CyclinD1抗体(ab16663)、NPAS4抗体(ab242003)、HRP标记羊抗兔二抗(ab205718)均购自英国Abcam公司。
1.2 主要仪器
正置智能型显微镜DM4B购自徕卡显微系统(上海)贸易有限公司;MyGo Pro二代全光谱便携式实时荧光定量PCR仪购自英国IT-IS公司;ELx808酶标仪购自美国Lonza公司;IX70荧光倒置显微镜购自日本奥林巴斯公司;Corning®Axygen®凝胶成像系统购自美国Corning公司。
2 方法
2.1 细胞培养
将HGC-27细胞与GES-1细胞培养于含有10%胎牛血清、1%青链双抗的RPMI1640培养基中,置于37 ℃、5%CO2的饱和湿度培养箱中培养,每2天更换一次培养基,观察细胞生长状态,待细胞融合至80%以上时,进行传代培养,取对数生长期细胞进行后续实验。
2.2 qRT-PCR法检测lncRNA MIAT、miR-203、NPAS4 mRNA的表达
收集各组对数生长期HGC-27、GES-1细胞,采用TRIzol试剂提取总RNA并逆转录得到cDNA,以cDNA为模板进行qRT-PCR反应,反应条件为95 ℃ 10 min;95 ℃ 10 s,60 ℃ 30 s,72 ℃ 30 s,共45个循环。采用2-ΔΔCt法计算lncRNA MIAT、miR-203、NPAS4 mRNA的相对表达量。qRT-PCR反应所需引物序列为:lncRNA MIAT上游引物5′-CATCTTACAACAGACCAGAA-3′,下游引物5′-ACAGTCC-ACAGAACATCCAT-3′;miR-203上游引物5′-GCATGCAGT-ATGTCTATCAG-3′,下游引物5′-ACTAGACTAGATAGTCAG-3′;NPAS4上游引物5′-CAGGATGACTCACACTGACAGT-ATTTTTAG-3′,下游引物5′-GTGGGAGAAGAGCTATTTAT-ATCACCAG-3′;GAPDH上游引物5′-AGAAGGCTGGGGC-TATTG-3′;下游引物5′-GCAGGAGGCATTGCTGATGAT-3′。所需引物均购自上海生工生物工程技术服务有限公司。
2.3 细胞转染与实验分组
取对数生长期HGC-27细胞,稀释细胞密度为2×105个/孔,接种于6孔板,待细胞融合至80%以上时,将细胞分组:对照组、si-NC组、si-MIAT组、miR-NC组、miR-203组、anti-miR-NC组、anti-miR-203组、si-MIAT+anti-miR-NC组、si-MIAT+anti-miR-203组、pc-NC组、pc-NAPS4组、pc-NAPS4+miR-NC组、pc-NAPS4+miR-203组。采用Lipofectamine 2000转染试剂盒将si-NC、si-MIAT、miR-NC、miR-203、anti-miR-NC、anti-miR-203、pc-NC、pc-NAPS4按照分组进行转染。转染48 h后,先采用qRT-PCR法进行转染效果的检测,再进行后续实验。
2.4 MTT法检测细胞增殖
收集转染成功的各组HGC-27细胞,制备细胞悬液,调整细胞密度为1×104个/mL,按照每孔100 μL细胞悬液接种于96孔板中,每组设3个复孔,置于常规培养箱中继续培养48 h后,每孔中加入20 μL的MTT溶液,置于常规培养箱中继续孵育4 h,采用酶标仪测定各孔490 nm处OD值。重复测定三次,计算平均值作为细胞活力。
2.5 Transwell实验检测细胞迁移与侵袭
迁移实验:收集各组HGC-27细胞,制备细胞悬液,调整细胞密度为1×105个/mL,取100 μL细胞悬液加入Transwell小室的上室,下室中加入常规培养基,每组设置3个复孔,继续培养24 h后,采用多聚甲醛固定,结晶紫染色,置于倒置显微镜下观察并计数。
侵袭实验:于Transwell小室的上室铺上Matrigel基底胶,加入100 μL细胞悬液,下室加入常规培养基,其余操作同迁移实验。
2.6 双荧光素酶报告基因实验
通过生物信息学网站预测可知,lncRNA MIAT与miR-203存在靶向关系,miR-203与NPAS4存在靶向关系。将lncRNA MIAT野生型报告基因载体(MIAT-WT)、突变型载体(MIAT-MUT)以及NPAS4野生型载体(NPAS4-WT)、突变型载体(NPAS4-MUT)分别与miR-NC、miR-203共转染至对数期生长的HGC-27细胞中,6 h后收集细胞,采用荧光素酶活性检测试剂盒检测萤火虫荧光素酶活性与海肾荧光素酶活性,以两者比值作为目的基因荧光素酶活性。
2.7 Western blot法检测细胞中MMP-3、MMP-9、CyclinD1、NPAS4蛋白表达
收集各组HGC-27细胞提取总蛋白,BCA法定量,取适量蛋白样品进行电泳、转膜,于5%脱脂奶粉中封闭2 h,加入MMP-3、MMP-9、CyclinD1、NPAS4一抗,4 ℃孵育过夜,加入二抗,37 ℃孵育2 h,DAB显色,凝胶成像系统分析条带灰度值,以GAPDH为内参蛋白,计算目的蛋白的相对表达量。
2.8 统计学方法

3 结果
3.1 lncRNA MIAT、miR-203、NAPS4在正常胃黏膜上皮细胞和胃癌细胞中的表达
qRT-PCR结果显示,与正常胃黏膜上皮细胞GES-1比较,胃癌细胞HGC-27中lncRNA MIAT表达显著升高(P<0.05),miR-203表达显著降低(P<0.05),NAPS4 mRNA表达显著上调(P<0.05),见表1。
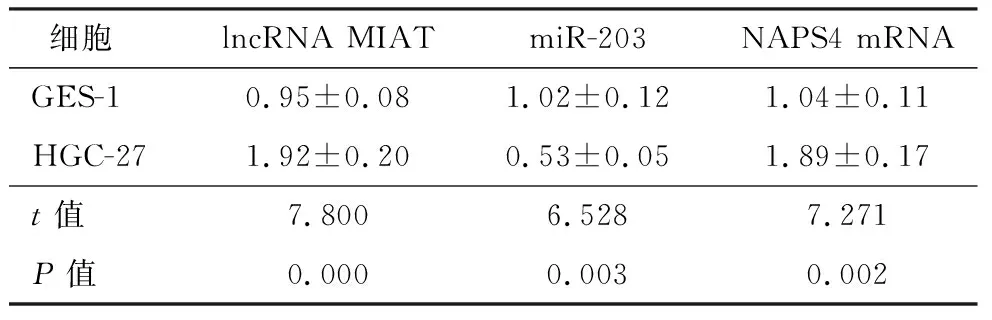
表1 lncRNA MIAT、miR-203、NAPS4在正常胃黏膜上皮细胞和胃癌细胞中的表达
3.2 沉默lncRNA MIAT对胃癌细胞增殖、迁移和侵袭的影响
Western blot法与Transwell实验检测发现,与对照组比较,si-NC组HGC-27细胞活力与侵袭、迁移能力以及MMP-3、MMP-9、CyclinD1蛋白表达均无显著差异(P>0.05);与si-NC组比较,si-MIAT组lncRNA MIAT表达显著降低(P<0.05),提示沉默lncRNA MIAT表达的HGC-27细胞构建成功,HGC-27细胞活力显著降低(P<0.05),细胞迁移与侵袭能力下降(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著降低(P<0.05)。结果见图1、2和表2、3。

图1 各组HGC-27细胞侵袭与迁移情况(×400)
表2 各组HGC-27细胞lncRNA MIAT表达以及细胞活力、侵袭与迁移个数的比较

图2 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达
表3 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达的比较
3.3 lncRNA MIAT靶向调控miR-203的表达
生物信息学网站StarBase(https://starbase.sysu.edu.cn/)预测可知,lncRNA MIAT与miR-203之间均存在连续的互补结合位点,见图3。双荧光素酶报告基因实验结果显示,MIAT-WT和miR-203共转染与MIAT-WT和miR-NC共转染比较,荧光素酶活性显著降低(P<0.05);MIAT-MUT和miR-203共转染与MIAT-MUT和miR-NC共转染比较,荧光素酶活性无显著差异(P>0.05),结果见表4。qRT-PCR结果显示,si-MIAT组(1.75±0.17)HGC-27细胞中miR-203表达显著高于si-NC组(0.95±0.07)(P<0.05)。

图3 lncRNA MIAT与miR-203的靶向结合位点
表4 双荧光素酶报告基因实验结果
3.4 lncRNA MIAT靶向miR-203调控NPAS4的表达
生物信息学网站TargetScan(https://www.targetscan.org/vert_71/)预测可知,NPAS4与miR-203之间存在连续的互补结合位点,见图4。双荧光素酶报告基因实验结果显示,NPAS4-WT和miR-203共转染与NPAS4-WT和miR-NC共转染比较,荧光素酶活性显著降低(P<0.05),NPAS4-MUT和miR-203共转染与NPAS4-MUT和miR-NC共转染比较,荧光素酶活性无显著差异(P>0.05),结果见表5。qRT-PCR与Western blot检测结果显示,miR-203组HGC-27细胞中NPAS4 mRNA及蛋白表达显著低于miR-NC组(P<0.05),anti-miR-203组HGC-27细胞中NPAS4 mRNA及蛋白表达显著高于anti-miR-NC组(P<0.05),另外si-MIAT组HGC-27细胞中NPAS4 mRNA及蛋白表达显著低于si-NC组(P<0.05),si-MIAT+anti-miR-203组HGC-27细胞中NPAS4 mRNA及蛋白表达显著高于si-MIAT+anti-miR-NC组(P<0.05),结果见图5、表6。
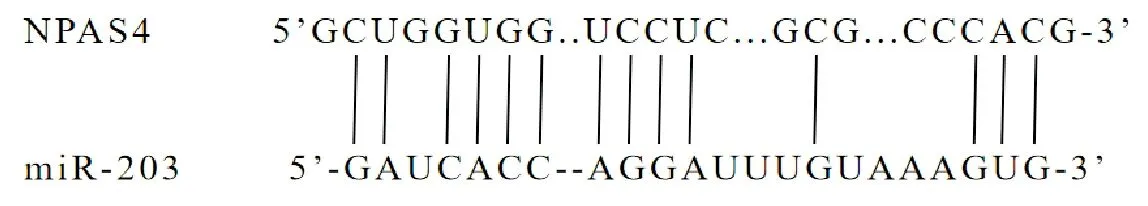
图4 NPAS4与miR-203的靶向结合位点
表5 双荧光素酶报告基因实验结果

图5 各组HGC-27细胞中NAPS4蛋白表达
3.5 过表达miR-203或抑制miR-203表达对胃癌细胞增殖、迁移与侵袭的影响
Western blot法与Transwell实验检测发现,与miR-NC组比较,miR-203组miR-203表达显著升高(P<0.05),细胞活力以及迁移与侵袭能力下降(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著降低(P<0.05);与anti-miR-NC组比较,anti-miR-NC组miR-203表达显著降低(P<0.05),细胞活力以及细胞迁移与侵袭能力显著升高(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著上调(P<0.05);此外,与si-MIAT+anti-miR-NC组比较,si-MIAT+anti-miR-203组miR-203表达显著降低(P<0.05),细胞活力以及细胞迁移与侵袭能力显著升高(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著上调(P<0.05)。结果见图6、7和表7、8。
表6 各组HGC-27细胞中NAPS4 mRNA、蛋白表达的比较
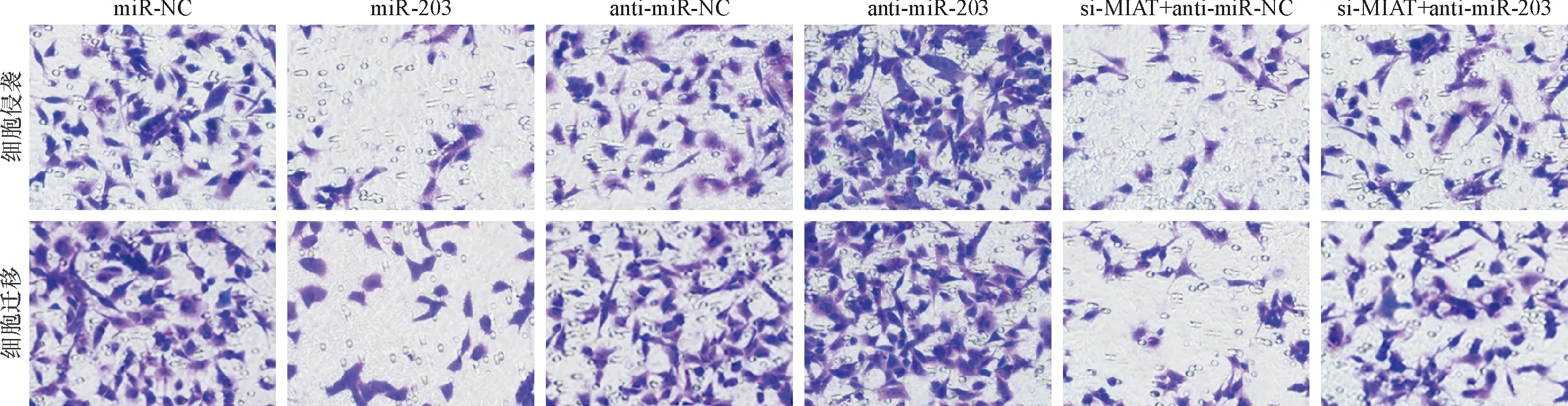
图6 各组HGC-27细胞侵袭与迁移情况(×400)
表7 各组HGC-27细胞中miR-203表达与细胞活力、侵袭与迁移个数的比较

图7 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达
3.6 过表达NAPS4对胃癌细胞增殖、迁移与侵袭的影响
Western blot法与Transwell实验检测发现,与pc-NC组比较,pc-NAPS4组NAPS4 mRNA和蛋白表达均显著上调(P<0.05),与细胞活力以及细胞迁移与侵袭能力显著升高(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著上调(P<0.05);此外,与pc-NAPS4+miR-NC组比较pc-NAPS4+miR-203组NAPS4 mRNA和蛋白表达均显著降低(P<0.05),细胞活力以及细胞迁移与侵袭能力显著降低(P<0.05),MMP-3、MMP-9、CyclinD1蛋白表达均显著下调(P<0.05)。结果见图8、9和表9、10。
表8 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达的比较

图8 各组HGC-27细胞侵袭与迁移情况(×400)
表9 各组HGC-27细胞中NAPS4 mRNA表达与细胞活力、侵袭与迁移个数的比较
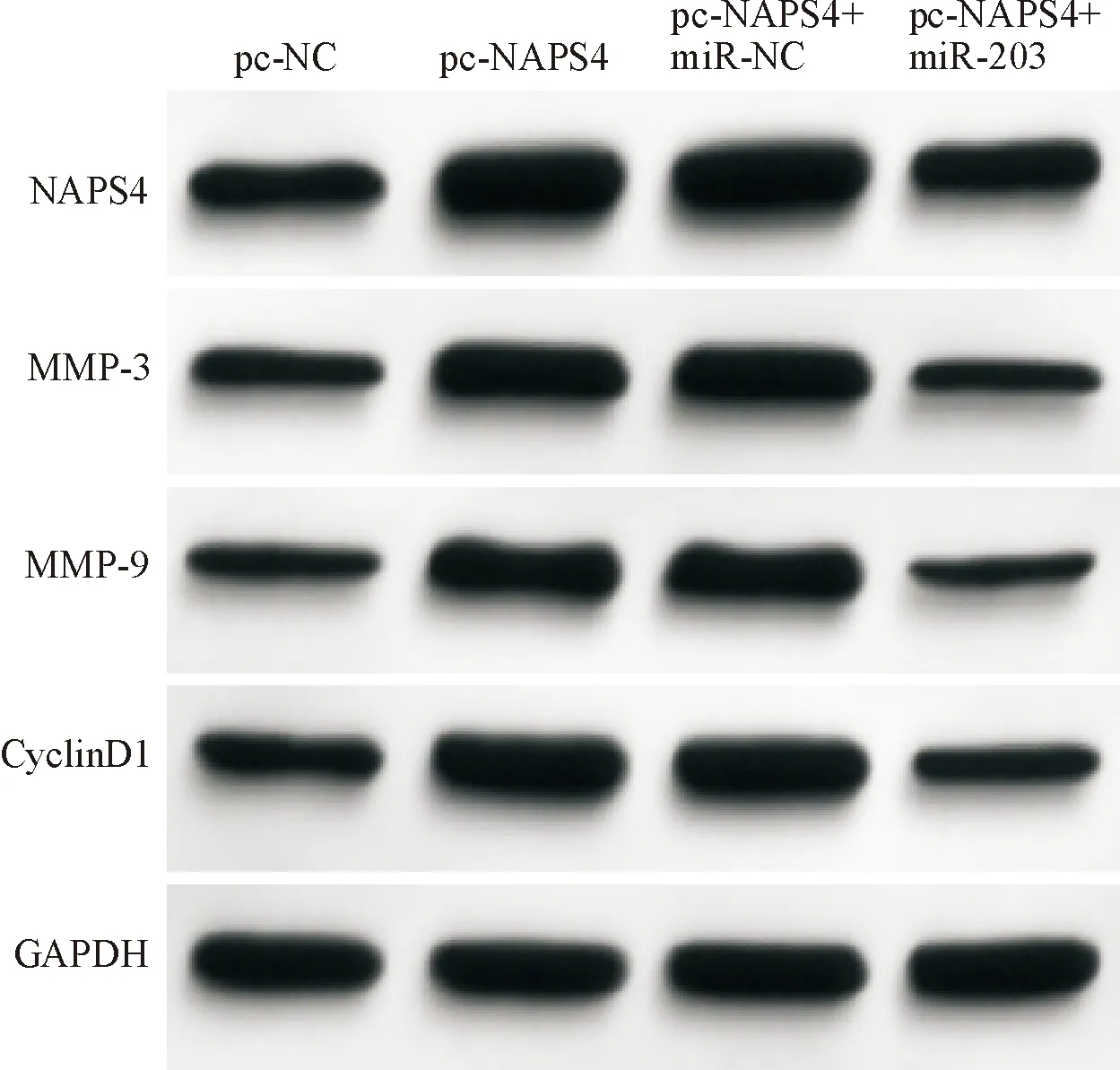
图9 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达
表10 各组HGC-27细胞中MMP-3、MMP-9、CyclinD1蛋白表达的比较
4 讨论
胃癌是我国多发的一种常见消化系统恶性肿瘤,死亡率占全国癌症相关死亡第二名[15]。目前胃癌的根治方法主要是彻底清除原发肿瘤以及转移浸润的癌旁组织,并减少复发,提高预后生存率,但由于胃癌发展过程涉及多种因素和机制,对于其诊断、治疗、预后都带来了极大的挑战[16]。lncRNAs在肿瘤中异常表达,参与肿瘤细胞增殖、迁移、侵袭与凋亡等恶性行为过程,可用作肿瘤治疗的新型靶点[17-18]。对于lncRNA MIAT在胃癌中的表达与作用,已有学者研究报道[19-20]。本研究结果显示,lncRNA MIAT在胃癌细胞中高表达,沉默lncRNA MIAT可显著降低HGC-27细胞活力与侵袭、迁移能力,下调MMP-3、MMP-9、CyclinD1蛋白表达。
前期我们采用miRNA芯片技术(Exipon mercury LNA microRNA 16.0)筛选胃癌组织中表达异常的miRNA,本研究结果显示,miR-203在胃癌细胞中表达显著下调,抑制miR-203的表达可促进胃癌细胞的增殖、迁移、侵袭等过程,而过表达miR-203可显著抑制以上细胞行为,此结果与前人研究结果一致[21]。但miR-203在胃癌中的调控机制尚未阐明,我们采用TargetScan等生物信息学数据库预测其靶基因,并通过双荧光素酶报告基因实验检测发现,miR-203与lncRNA MIAT和NAPS4均具有靶向关系,且抑制miR-203表达可显著逆转沉默lncRNA MIAT对NAPS4表达、细胞活力以及迁移与侵袭能力的抑制作用,表明lncRNA MIAT可通过靶向miR-203调控NAPS4表达,在胃癌中发挥促癌作用。
NAPS4是存在于神经系统中的一种转录因子,与学习行为建立和记忆形成存在密切联系,但也有研究证明,其作用并未局限于神经系统,在胰岛β细胞毒性中也可发挥作用[22]。本研究结果显示,胃癌细胞中NAPS4呈高表达,与lncRNA MIAT表达趋势一致;过表达NAPS4可促进胃癌细胞增殖、迁移与侵袭,同时上调MMP-3、MMP-9、CyclinD1蛋白表达;且miR-203过表达可显著削弱NAPS4过表达对胃癌细胞恶性行为学的促进作用。基于以上研究结果,表明lncRNA MIAT可通过miR-203调控NAPS4促进胃癌细胞增殖、侵袭、迁移。
综上所述,lncRNA MIAT可通过靶向miR-203调控NAPS4,促进胃癌细胞增殖、迁移与侵袭,这为胃癌诊断提供了新靶点,为胃癌治疗与预后提供了新的思路。但由于胃癌发生涉及基因表达多样,lncRNA MIAT作用机制复杂,关于lncRNA MIAT参与胃癌发生的机制还需深入探索。
