促进油茶花粉萌发的营养物质配比研究
2022-02-11冯都煌刘会云吴新蕾丁佐楠张文元翁小婷汪加魏
冯都煌 刘会云 张 莉 吴新蕾 丁佐楠 张文元 谭 澍 翁小婷 汪加魏
(江西农业大学林学院,江西 南昌 330045)
油茶(Camellia oleifera)是山茶科山茶属的常绿灌木或小乔木,别名茶子树、茶油树、白花茶等。油茶与油橄榄(Oleae uropaea)、油棕(Elaeis guineensis)、椰子(Cocos nucifera)并称为世界四大木本油料植物,是我国南方特有的重要木本粮油树种[1],随着油茶种植面积不断增大,其经济和生态价值也不断突显[2],而目前“落花落果”、“花而不实”已成为限制油茶产量提高的最主要原因之一[3]。而花粉活力、花粉管生长均可能会影响油茶的坐果[4]。
蔗糖在花粉萌发中有提供能源和调节渗透压的作用[5],对花粉萌发率和花粉管长度影响较大[6]。硼(B)能促进花粉管萌发,缺硼会使酸性果胶质在顶端大量富集,导致花粉管破裂而无法正常授粉[7-8],花期喷施适宜浓度的硼酸可改善果实品质和坐果率[9-11]。尿素中氮素可参与合成蛋白质、核酸、磷脂及其他必需有机氮,盛花期喷施适宜浓度的硼酸和尿素可能提高果树的坐果率[11-13]。钙(Ca)在细胞骨架的组装[14]、分泌小泡的运输和融合[15]、花粉管顶端的脉冲式生长[16-17]、花粉管生长调控[18-19]等方面发挥着重要作用。磷(P)和钾(K)可提高酶及功能蛋白活性[18],同时参与植物体内糖类的运输,因此花期喷施磷酸二氢钾(KH2PO4)可作为油茶重要的培育措施[20]。PEG-3350 可改变花粉内膜结构和通透性,进而可促进花粉萌发和花粉管伸长[21-22]。维生素C 是一种还原剂,维生素C 有可能促进油茶花粉萌发[23-25]。维生素B1是植物生长发育中的重要调节成分之一,可能具有促进离体花粉萌发的作用[26]。本文采用的离体花粉固体培养基法与柱头上花粉萌发的条件较接近,为实验室促进油茶花粉萌发的养分筛选提供了技术保障[27]。因此,本研究旨在探讨蔗糖、硼酸、尿素、氯化钙(CaCl2)、KH2PO4、PEG-3350、维生素C、维生素B1等对花粉萌发率和花粉管萌发长度的影响,筛选出对油茶花粉萌发具有重要促进作用的营养成分,为促进油茶花粉萌发的营养混合液配比提供参考依据。
1 材料与方法
1.1 试验材料
2019 年11 月中旬于江西省林木育种中心(115°39′16.97″E,18°57′11.64″N),选取生长良好、进入盛花期的‘赣无2’(国S-SC-CO-026-2008)、‘赣70’(国S-SC-CO-025-2010)、‘长林166’(国R-SC-CO-008-2011)油茶为试验对象。分别剪取含苞待放的花枝,带回实验室进行水培,待花完全自然开放后,用硫酸纸盒收集花药并置于硅胶干燥器中干燥4~8 h,收集花粉置于离心管中干燥低温(-20 ℃)保存[28-29]。
1.2 花粉固体培养法
在凹槽载玻片的凹槽内均匀而平坦的滴入培养基,待其凝固后,用毛笔蘸取少量花粉均匀的播撒在培养基上,培养皿中垫2 层湿滤纸,将载玻片放入(有花粉粒的一面朝上)。在20~25 ℃、40%湿度及黑暗的恒温培养箱中培养[30]。用光学显微镜观察每个凹槽内花粉的萌发情况,每个凹槽随机选取3 个视野(取其平均值视为1 次重复),进行3 次重复。其中每个视野花粉数目不少于30 粒,以花粉管萌发长度大于或等于花粉粒直径视为萌发标准,测量花粉管萌发长度(PGL),按公式(1)计算花粉萌发率(PGR)。

1.3 试验设计
1.3.1 基础培养基筛选试验
100 g/L 的蔗糖和0.1 g/L 的硼酸作为促进普通油茶花粉萌发的依据[29],本试验采用完全正交试验(表1),2、4、8 h 时分别统计花粉萌发数和花粉管萌发长度,筛选出最佳蔗糖-硼酸组合作为单因素试验的基础培养基。
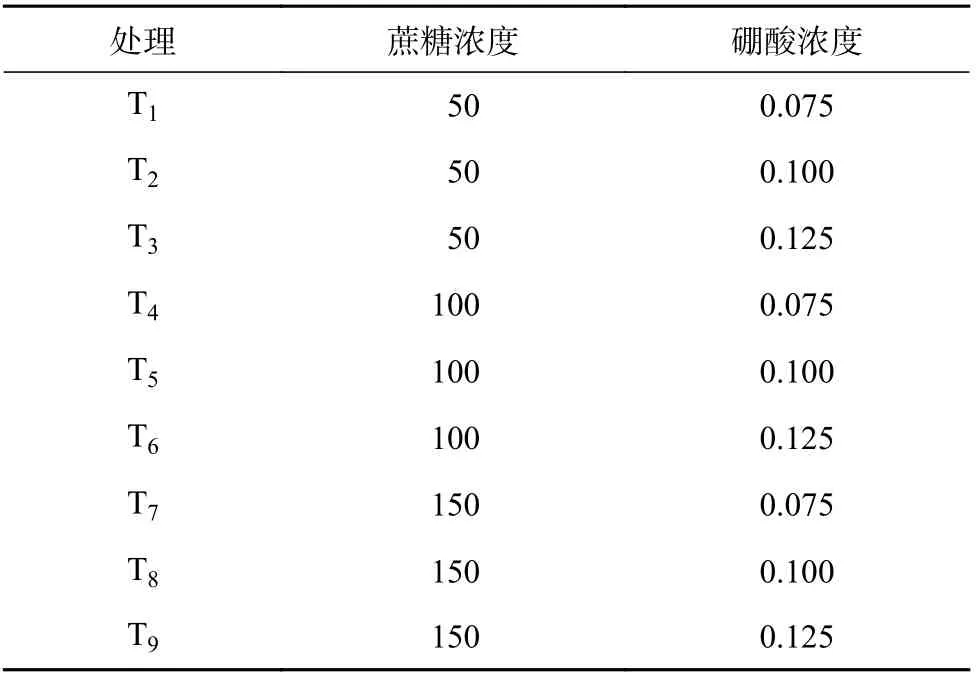
表1 油茶花粉基础培养基的试验处理Table 1 The treatments of base medium for the pollen germination of C. oleiferag/L
1.3.2 最佳培养基筛选试验
1)单因素试验以筛选出的最适蔗糖-硼酸组合(5%琼脂)为基础培养基,分别对6 种重要的营养物质进行单因素添加试验,培养8 h 后测定花粉萌发率和萌发长度,筛选出具有促进作用的营养物质及其浓度(表2)。2)正交试验在最适蔗糖-硼酸(5%琼脂)基础培养基上,添加单因素试验筛选出的具有显著促进作用的营养物质,采用正交试验,培养8 h 后测定花粉萌发率和萌发长度,筛选出最佳养分组合。
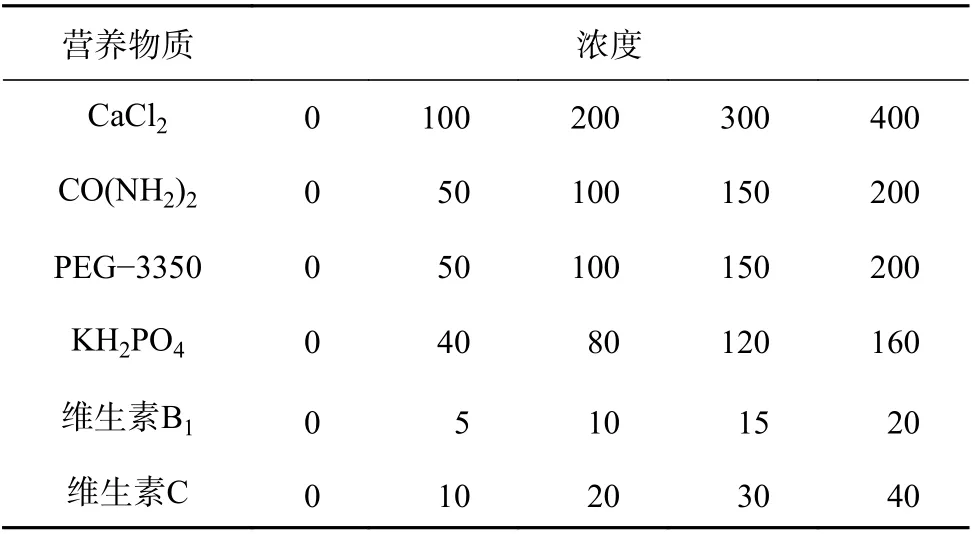
表2 培养基成分的浓度处理Table 2 The concentration treatments of nutrients in the mediummg/L
1.4 数据处理
统计显微镜视野中的花粉总数、花粉萌发数、花粉管长度,运用Microsoft Excel 2010 整理原始数据及初步分析,利用SPSS 21.0 对各试验进行一般线性方差分析和LSD 法多重比较。
2 结果与分析
2.1 基础培养基中蔗糖-硼酸对花粉萌发的影响
由表3 可知,培养2、4、8 h 后,均为T4的萌发率最高,分别达30.58%、31.09%、31.49%,比T8培养2 h(13.23%)后、T7培养4 h(18.22%)和8 h(15.41%)后的最低花粉萌发率显著提高了131.14%、70.64% 和104.35%(P<0.05)。最高萌发率均显著高于T7~T9的萌发率(P<0.05)。培养2 h 后,T6的花粉管萌发长度最长,达626.75 μm,比T8的最短花粉萌发长度显著长298.43 μm(P<0.05);培养4 h 和8 h 后,均为T2的花粉萌发长度最长,分别达946.70 μm 和922.12 μm,4 h 后最长花粉萌发长度是T8的最短花粉萌发长度的2.55 倍,8 h 时,最长花粉管长度是T7的最短花粉萌发长度的1.98 倍(P<0.05)。最长花粉萌发长度均显著高于T7~T9的花粉萌发长度(P<0.05)。

表3 基础培养基中油茶花粉萌发率及花粉管长度Table 3 The PGR and PGL of C. oleifera in the basal medium
油茶‘赣无2’、‘赣70’、‘长林166’3 个品种的花粉通过2、4、8 h 培养后,其花粉萌发率及花粉管萌发长度分析结果见表3,T4对油茶花粉萌发率的促进效果最佳,T7~T9对油茶花粉管萌发长度的促进效果较差。本研究筛选出最适油茶花粉萌发的基础培养基为100 g/L 蔗糖+0.075 g/L 硼酸+5 g/L 琼脂。
2.2 培养基养分的筛选
2.2.1 尿素对油茶花粉萌发的影响
由表4 可知,尿素对3 种油茶的花粉管萌发和花粉管的伸长均有一定的促进作用但不明显。其中,仅50 mg/L 尿素对‘赣无2’花粉、‘赣70’花粉,100 mg/L 尿素对‘长林166’花粉的萌发具有促进作用,萌发率分别较CK 提高了30.78% 、7.31%、4.93%;100~200 mg/L 尿素对其萌发存在抑制作用。50 mg/L 尿素对‘赣无2’,100 mg/L尿素对‘长林166’花粉管伸长促进作用较优,分别比CK 长233.13、187.31 μm。

表4 添加尿素的培养基中油茶花粉萌发率及花粉管长度Table 4 The PGR and PGL of C. oleifera in the medium with CO(NH2)2
2.2.2 CaCl2对油茶花粉萌发的影响
由表5 可知,一定浓度的CaCl2对花粉萌发和花粉管伸长均具有一定促进作用,随着CaCl2的浓度增加,基本都呈现先增后减趋势,较高浓度CaCl2表现为一定的抑制作用。200 mg/L 和400 mg/L 的CaCl2对‘赣无2’花粉萌发具一定的促进作用,萌发率分别较CK 提高了18.43%和24.21%;100~400 mg/L 的CaCl2对‘赣70’和‘长林166’花粉萌发均有一定促进作用且以100 mg/L 促进效果最佳,分别较CK 提高了49.04%和34.32%。100 ~200 mg/L 的CaCl2对‘赣无2’,100、300 mg/L 的CaCl2对‘赣70’,100 mg/L 的CaCl2对‘长林166’花粉的管伸长具一定的促进效果,均以100 mg/L 促进效果最优,分别比CK长120.79、89.58、13.22 μm。

表5 添加CaCl2 的培养基中油茶花粉萌发率及花粉管长度Table 5 The PGR and PGL of C. oleifera in the medium with CaCl2
2.2.3 KH2PO4对油茶花粉萌发的影响
由表6 可知,花粉萌发与花粉管萌发长度随着KH2PO4的浓度增加,基本呈现先增后减的趋势。40 mg/L 的KH2PO4处理下,‘赣无2’花粉的萌发率(50.73%)显著高于CK(P<0.05),是CK 的2.2 倍;‘赣70’花粉的萌发率在80 mg/L的KH2PO4浓度下达到36.87%,较CK 提高了57.63%,具有显著促进作用。80 mg/L 的KH2PO4处理下,‘赣无2’花粉萌发长度(1 062.64 μm)显著比CK 长324.62 μm(P<0.05);40 mg/L 的KH2PO4对‘赣70’花粉(720.53 μm)和‘长林166’(938.07 μm)花粉萌发长度的促进效果显著(P<0.05),分别比CK 长145.87 μm、475.7 μm。
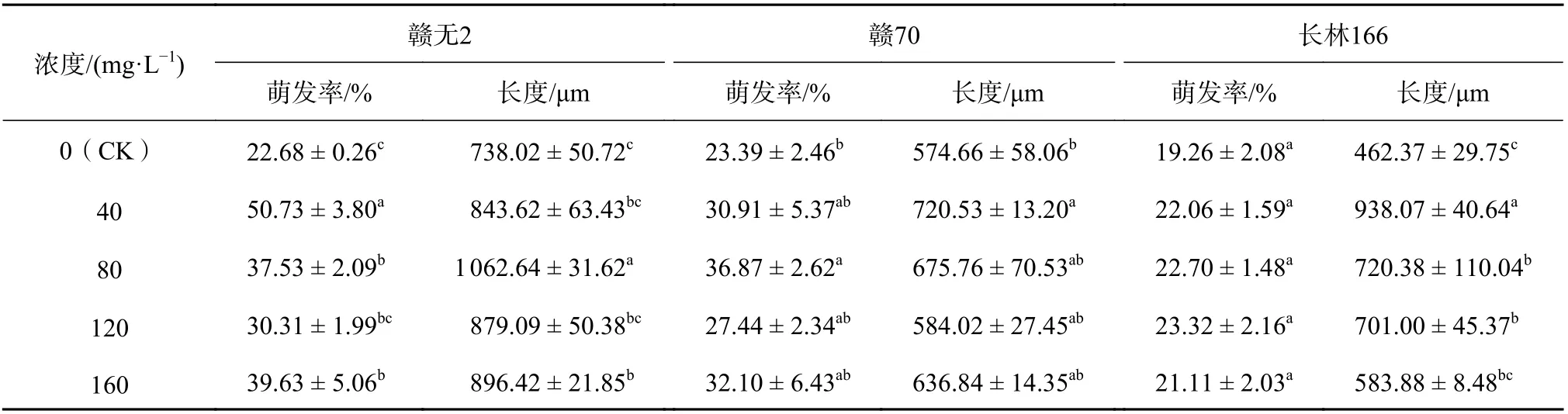
表6 添加KH2PO4 的培养基中油茶花粉萌发率及花粉管长度Table 6 The PGR and PGL of C. oleifera in the medium with KH2PO4
2.2.4 PEG-3350 对油茶花粉萌发的影响
由表7 可知,一定浓度的PEG-3350 对‘赣无2’、‘赣70’、‘长林166’花粉萌发和花粉管的伸长均具有促进作用,对‘赣无2’花粉萌发的促进作用随着浓度升高而上升;对‘赣70’、‘长林166’花粉萌发在200 mg/L 的PEG-3350时有所下降。对花粉管生长则基本呈现先增后减趋势。100~200 mg/L 的PEG-3350 处理对‘长林166’花粉萌发促进作用显著(P<0.05)。‘赣无2’和‘长林166’花粉分别在200 mg/L和150 mg/L 的PEG-3350 浓度条件下萌发率达最高分别为38.58%、71.53%,显著高于CK(P<0.05),是CK 的1.7 倍和3.7 倍。150 mg/L 的PEG-3350 处理下,‘赣无2’(1 571.17 μm)、‘赣70’(846.44 μm)、‘长林166’(1 575.93 μm)花粉萌发长度均达最长,比CK 长822.06、271.78、1 113.56 μm,均具有显著促进作用(P<0.05)。

表7 添加PEG-3350 的培养基中油茶花粉萌发率及花粉管长度Table 7 The PGR and PGL of C. oleifera in the medium with PEG-3350
2.2.5 维生素C 对油茶花粉萌发的影响
由表8 可知,花粉萌发随着维生素C 的浓度增加,基本呈现先增后减趋势,而花粉管长度则无明显增减趋势。维生素C 对‘赣无2’、花粉萌发具有显著抑制作用(P<0.05);对‘赣70’花粉仅在20 mg/L 条件下萌发率(24.09%)较CK 提高了2.99%,其余均表现出一定的抑制作用;对‘长林166’花粉萌发具一定的抑制作用。维生素C 对‘赣70’花粉管伸长有一定抑制作用,在10 mg/L 的维生素C 条件下抑制作用显著(P<0.05);20~30 mg/L 的维生素C 对‘长林166’花粉管伸长促进效果显著(P<0.05),20 mg/L 时花粉萌发长度达最长,比CK 长124.76 μm。
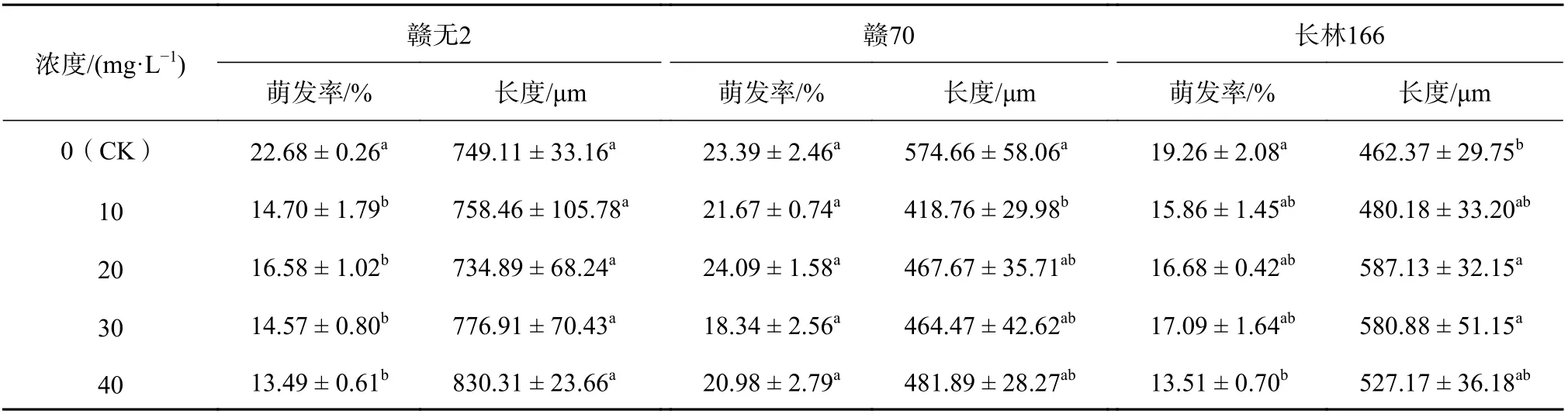
表8 添加维生素C 的培养基中油茶花粉萌发率及花粉管长度Table 8 The PGR and PGL of C. oleifera in the medium with Vitamin C
2.2.6 维生素B1对油茶花粉萌发的影响
由表9 可知,花粉萌发和花粉管伸长随维生素B1的浓度增加,基本呈现先增后减趋势。维生素B1对‘赣无2’、‘赣70’、‘长林166’花粉管伸长均具有一定的抑制作用,对‘赣无2’的花粉管伸长抑制作用显著(P<0.05)。维生素B1对‘赣无2’花粉管伸长具有显著促进作用(P<0.05),在20 mg/L 的维生素B1处理下萌发长度较CK 长737.5 μm,是CK 的1.98 倍;‘长林166’在15 mg/L 的维生素B1处理下花粉管长度长于CK520.26 μm,是CK 的2.13 倍,促进作用显著(P<0.05)。

表9 添加维生素B1 的培养基中油茶花粉萌发率及花粉管长度Table 9 The PGR and PGL of C. oleifera in the medium with Vitamin B1
2.3 KH2PO4 与PEG-3350 对油茶花粉萌发的影响
由表10 可知,不同浓度的KH2PO4与PEG-3350 处理组合对油茶花粉萌发及花粉管伸长均有一定的促进作用。通过分析花粉萌发率,‘赣无2’、‘赣70’的花粉萌发率在各处理间的差异不显著,T13达最高,比最低萌发率提高了40.83%,T10、T11、T12、T17的萌发率均稍低于最高萌发率;‘赣70’花粉在处理2 的萌发率最高,较最低萌发率提高了23.68%,T15的花粉萌发率略低于最高萌发率;‘长林166’花粉在T11的萌发率高达46.16%,与其他处理间差异较显著,比最低萌发率显著(P<0.05)提高了33.64%。通过分析花粉管萌发长度,3 种油茶的花粉萌发长度在各处理间存在显著差异(P<0.05)。‘赣无2’在T18的花粉管萌发最长为1 988.47 μm,显著长于T10、T12、T13,且比最短花粉管显著长522.69 μm(P<0.05);‘赣70’花粉在T17的花粉管长度最长为1 743.85 μm,显著长于T10~T15的花粉管长度,花粉管长度在T15最短为1 211.91 μm(P<0.05);‘长林166’花粉在T11、T12、T14的花粉管长度长于处理T13、T15、T16、T17、T18且差异显著,T12的最长花粉管长度比T16的最短花粉管长度提高了90.56%。综合分析萌发率与萌发长度,在基础培养基条件下,60 mg/L 的KH2PO4+150 mg/L 的PEG-3350 对油茶花粉萌发的促进效果最佳。

表10 添加KH2PO4 与PEG-3350 的培养基中油茶花粉萌发率及花粉管长度Table 10 The PGR and PGL of C. oleifera in the medium with KH2PO4 and PEG-3350
3 结论与讨论
花粉萌发和花粉管伸长是植物有性生殖的重要生理过程,除了受自身遗传学特性、湿度、温度、光照等环境因素外,还受N、P、K、Ca 等营养元素的影响,本试验研究了蔗糖、硼酸、尿素、CaCl2、KH2PO4、PEG-3350、维生素C、维生素B1对花粉萌发及花粉管萌发长度的影响。
本研究发现100 g/L 蔗糖+0.075 g/L 硼酸的基础培养基最适于油茶花粉萌发。高浓度蔗糖对花粉萌发和花粉管的伸长存在一定抑制作用,可能是高浓度蔗糖对花粉渗透压产生了负面效应,从而间接影响花粉萌发和花粉管伸长[7]。
在培养基养分的单因素试验中发现,50~100 mg/L 的尿素对花粉萌发和花粉管伸长有一定的促进作用,50 mg/L 的尿素对‘赣无2’花粉萌发率较CK 提高了30.78%,该浓度稍低于袁德义研究的100~150 mg/L 适宜尿素浓度[31]。100~200 mg/L 的CaCl2对花粉萌发和花粉管伸长具有一定的促进作用,此浓度的CaCl2处理‘赣无2’花粉的萌发率是CK 的1.24 倍,花粉管伸长量比CK 长120.79 μm,该结论与100 mg/L 钙能促进海棠(Malus spectabilis)[32](萌发率最高达76.43%)、思茅松(Pinus kesiyavar.langbianensis)[33](萌发率最高达42.16%)花粉萌发的结论基本相同;而也有研究表明CaCl2对油茶花粉萌发具有一定抑制作用[31],其作用机制尚不十分清楚。40~120 mg/L的KH2PO4对花粉萌发和花粉管伸长具有显著促进作用,其中40 mg/L 的KH2PO4处理‘赣无2’花粉的萌发率最高达50.73%(是CK 的2.24 倍);该结果与50~100 mg/L 的KH2PO4对油茶花粉萌发的促进作用的结果基本一致[31]。50~200 mg/L的PEG-3350 对花粉萌发和花粉管伸长有一定的促进作用,其中150 mg/L 的PEG-3350处理的‘长林166’花粉萌发率最高达71.53%,是CK的3.71 倍;该结果与150 mg/L的PEG-3350 可显著促进苹果花粉萌发率的结果相似[34]。维生素C 对油茶花粉萌发促进作用不显著或具一定抑制作用,维生素B1对油茶花粉萌发具有一定抑制作用,而对花粉管伸长具有一定促进作用,‘赣无2’花粉在20 mg/L 的维生素B1条件下萌发长度达到最大值1 486.61 μm,是CK的1.98 倍;这与20 mg/L 的维生素B1可显著促进文冠果花粉萌发的研究结果基本一致[35],然而目前维生素C 和维生素B1对油茶花粉萌发的作用机制还尚不清楚[25],有待进一步研究。不同油茶品种在同一养分浓度的培养基处理下,花粉萌发率和花粉管伸长存在差异。这可能是由于不同油茶品种的花粉自身所含营养差异、花粉活力高低,以及花粉对不同营养元素浓度的敏感性或耐受性不同所引起。有研究表明,离体培养基中蔗糖、H3BO3和Ca(NO3)2的浓度配比因柳树(Salix babylonica)个体不同而略有差异[36],不同柚(Citrus grandis)品种促进花粉萌发的最佳蔗糖和硼酸浓度配比不同[37],3 个李(Prunus salicina)品种花粉在相同蔗糖-硼酸培养基中的花粉萌发率不同[38];烟草(Nicotiana tabacum)K326 花粉在不同发育时期的可溶性糖、可溶性淀粉和ATP 酶等内含物含量均可能对花粉活力有一定影响[39]。此外,花粉在不同温度条件下对相同营养物质的敏感程度不同[40]。因此不同植株或不同品种(种源)的花粉活力、最适花粉萌发条件均可能存在差异[35,37,41]。而不同油茶品种花粉对不同营养元素浓度的敏感性的相关研究较少,其作用或影响机制有待进一步研究。
综上所述,促进油茶花粉萌发的基础培养基为100 g/L 蔗糖+0.075 g/L 硼酸+5%琼脂。对油茶花粉萌发而言,KH2PO4(40~120 mg/L)、PEG-3350(50~200 mg/L)具有显著促进作用;尿素(50~100 mg/L)和CaCl2(100~200 mg/L)具有一定的促进作用;维生素C、维生素B1均存在一定的抑制作用。促进油茶花粉萌发的最佳营养物质配比为基础培养基(100 g/L 蔗糖+0.075 g/L硼酸+5 g/L 琼脂)+60 mg/L 的KH2PO4+150 mg/L的PEG-3350。本研究将为促进油茶花粉萌发的营养混合液配比及人工喷雾授粉技术提供一定参考,将有利于油茶开花坐果调控及其增产培育。
