登上高处的生命:适应高原低氧的生理机制与过程*
2022-02-10黄大明何牧张宇明JayStorz
黄大明 何牧 张宇明 Jay F.Storz
(1.中国清华大学生命学院生态学实验室,北京 100084;2.美国内布拉斯加大学生物学院,内布拉斯加 68588)
0. 引言
在高海拔地区,脊椎动物由于O2分压(PO2)降低限制有氧代谢速率,影响生存和繁殖等生理功能。为了更好地合成ATP(三磷酸腺苷)来维持足够的能量转化率,O2转移系统需要将供给组织细胞线粒体的O2与代谢中电子传递链中氧化磷酸化的化学利用O2需求匹配起来。因此,调整呼吸和心血管减少对吸入的低氧空气的氧需求,尽量减少组织细胞未利用O2的流失[43,62,88,111,106,108]和/或调整代谢以减少O2需求[84]。对生活在高海拔地区的恒温动物,由于需要应对慢性低氧条件下维持体温、繁殖与生活的能量需求,包括雌性哺乳+觅食+挖洞+领域性+逃避捕食+其它日常的应急等,通过抑制代谢减少系统O2需求并不是可行的长期对策。由于高海拔的环境特点是低温,尽管O2的可利用率降低,但恒温动物还要被迫增加代谢产热。因此,代谢需氧量在高海拔地区不减反增。因此,提高O2利用率的生理调整,有助于保持循环系统O2流量。与低海拔地区相比,高海拔地区的哺乳动物和鸟类在呼吸、心血管和代谢方面表现出许多独特差异[43,62,63,88,91,107,108,111]。
1. 表型弹性与遗传适应
一般耐低氧的生理调节可以表现为环境引起的可逆或不可逆的遗传弹性变化或演变,如果对特定环境变化的表型与环境选择压力一致,就是适应弹性,适应弹性有时会掩盖遗传性状的变异。物种迁移到新环境时,如果新环境条件在原生适应范围内,那么会弹性适应,如果超出适应范围,就会不适应,一些低海拔生命在高海拔会对低氧表现出遗传不适应。
2. 高原适应生理基础
2.1 身体有氧运动与代谢
所有动物有氧运动的研究都集中在高海拔适应[62,108],恒温动物的许多生理生态指标,如持续运动能力和在寒冷中的活动能力,都与氧代谢率直接相关,可以用耗氧量(VO2)来衡量。最大代谢率(MMR)(最大耗氧速率,VO2max)可通极限运动或极限低温的耗氧量来获得。对于低温,恒温动物会增加耗氧代谢产热来维持恒温,如果足够冷,会产生最大产热率,达到最大耗O2速率(也称为顶峰代谢)[35,36,108]。
2.2 O2的运输和利用途径
有机体的有氧代谢主要是组织细胞中线粒体的氧化磷酸化过程中消耗O2,O2需要经过肺、心脏、血管、血液循环送到线粒体(图1)。包括5个步骤:(1)肺通气,吸入空气向肺表面运动;(2)肺氧扩散,氧在空气-血液界面上的扩散;(3)循环氧运输,动脉血液中氧向组织毛细血管运动[几乎所有的氧都结合在红细胞的血红蛋白(Hb)上];(4)组织氧的扩散,氧从毛细血管向组织细胞的线粒体扩散;(5)线粒体的氧利用,通过氧化磷酸化消耗氧产生ATP。O2分压是氧扩散的动力。图1b所示外部空气的PO2和线粒体中的PO2的压力梯度。而CO2沿同一途径反向运动。原则上,以上所有因素都与低氧适应弹性有关[43,62,111]。
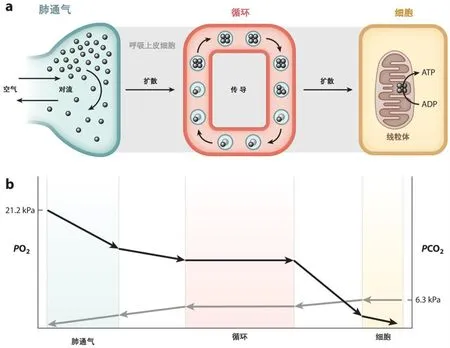
图1 O2和CO2的传输途径。(a)哺乳动物心肺系统中气体传输图示。氧气通过几个扩散和对流步骤从大气输送到组织线粒体:肺通气、肺氧扩散、循环氧输送、组织氧扩散和线粒体氧利用。Hb被示意性地画为四聚体(4个亚单位)蛋白质,4个氧分子可逆地结合到Hb上。(b)扩散的氧气流量由PO2的梯度驱动,导致PO2从一个阶段下降到下一个阶段。当Hb将O2释放到组织时,血液PO2从组织毛细血管(动脉)入口下降到传出(静脉)出口。PO2也随着离毛细血管的距离而下降,因此细胞PO2的梯度应反映毛细血管PO2和扩散距离的变化。CO2的反向传输存在类似途径。缩写:ADP,二磷酸腺苷;ATP,三磷酸腺苷;Hb,血红蛋白;PO2,O2的分压。
3. 有氧运动和代谢弹性演化的生理基础
研究脊椎动物有氧运动对低氧耐受性和其它氧运输机制随海拔变化的工作很多[62]。
3.1 VO2max随海拔变化的模式
在低氧环境下,有几个高原种类的有氧运动能力比其低海拔同类更强。在人类[6,10],鹿鼠(Peromyscus maniculatus)[14,15,16,58,114]和叶耳鼠(Phyllotis xanthopygus)[85]中,高海拔类群的VO2max大于低海拔类群的VO2max,但在海平面上(常氧)PO2并不一定不同。在低PO2+低温环境中,有更高的VO2max支持有氧生热和/或运动是有益的。还无法确定这种高海拔表型是低氧适应表型还是遗传型(见图 2)[6,14,15,16,58,114]。
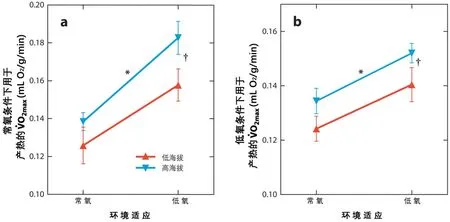
图2 高原小鼠(Peromyscus Mangulast)在低氧条件下比低海拔同类具有更高的有氧能力。低氧时的耗氧容量被计为在发热或锻炼期间O2消耗的最大速率(VO2max)。
随海拔升高,O2运输的表型弹性和适应演化与VO2max的动态。在常氧条件下,影响人与脊椎动物VO2max的因素很多。有人认为:O2循环中,常氧水平决定VO2max,因为常氧水平决定O2运输到代谢组织的速率[1]。其它认为:在O2运输中的所有步骤的容量有比例,使得每个步骤对VO2max有类似的调控[120]。增加O2运输途径中每个步骤的能力都可以增加VO2max,但强度不同。而从毛细血管到线粒体的O2扩散能力在常氧和低氧中的强度最大[92,94,117]。实验显示:大鼠跑步耐力增加的人工选择显示,7代后VO2max的增加完全可以解释为运动肌氧扩散和线粒体氧利用能力的增加,而肺功能或通气、循环氧运输没有任何变化[37,39]。在经过15代选择后,VO2max的继续增加是由于骨骼肌的进一步变化,以及心脏泵出的血量增加(即心脏输出量的增加),组织循环氧传递增加,肺通气和肺氧扩散能力增加[30,39,50]。总之,初始期,O2运输途径中单一步骤的改进,就能初步适度提高VO2max,但VO2max的持续增强需要其它步骤也有改进。
在高原类群中,VO2max的增加主要由于运动和/或产热过程中活跃的组织中O2扩散和线粒体O2利用能力的增加(即骨骼肌和棕色脂肪组织),也可能与循环和肺氧传输能力增加有关。不同步骤的相对影响在低氧与常氧条件下可以改变,在低氧条件下肺氧传输有更大的影响[92,117]。因此,高原低氧增加VO2max的生理机制可能与常氧条件下VO2max不同。在高海拔地区,氧气运输途径的每一个步骤的能力变化都可能导致O2运输的表型弹性和适应演化的变化。研究表明,环境诱导的变化和遗传固有变化都会引起更显著的变化,并在多个步骤中进一步变化。在高原适应中似乎没有单一限制因素,而是综合的生理变化,共同提升O2运输。
许多种类在高海拔进行强烈代谢活动时,其运动肌肉线粒体利用氧能力提高。例如,高海拔鹿鼠的腓肠肌线粒体进化出更大的呼吸能力[59],还有用于运动和颤抖性产热的后肢肌肉增大[33,76],表现为氧化功能的肌纤维(富含线粒体)比例增加[58,90]并且其中的线粒体也增加[59]。同样,斑头雁(Anser indicus)和湍鸭(Merganetta armata)运动肌纤维中的线粒体呼吸能力更强[20,89,95],鹿鼠腓肠肌线粒体呼吸强度在慢性低氧、成年期或早期发育过程中不发生变化[58,59,72],表明肌纤维线粒体氧气利用能力增强是遗传性的。然而,生活在高海拔的人类(与其它高原哺乳动物和鸟类不同,他们在高海拔不能维持特别高的代谢率)运动肌纤维中的线粒体没有进化出更大的氧利用能力[38,48,49]。相反,在O2运输途径的其它步骤的变化似乎有助于增加低氧下的VO2max[29]。
氧气扩散到骨骼肌的能力在许多高原类群中也有增强。对哺乳动物和鸟类的研究表明:与本地环境相比,高海拔类群骨骼肌中的毛细血管往往比低海拔类群多[21,49,54,61,90]。在一些观测中,在相同环境中饲养的高海拔类群和低海拔类群之间,肌肉毛细血管的这些差异持续存在[58,89],表明这种差异有进化的遗传基础。一些高原人种也进化出较高的肌肉肌红蛋白量[53],其肌细胞中的线粒体位于靠近毛细血管的位置,从而减少了细胞内的氧气扩散距离[59,89]。鹿鼠腓肠肌毛细血管有增加(见图3),并且O2运输对慢性低氧不敏感[58,59,72]。这可能是另一个氧运输途径演化的方面,有助于增强有氧能力。骨骼肌中O2扩散能力和线粒体O2利用增强的机制尚不完全清楚,但在高海拔鹿鼠中,与其能量代谢、肌肉发育和血管发育相关基因表达差异有关[14,16,90]。
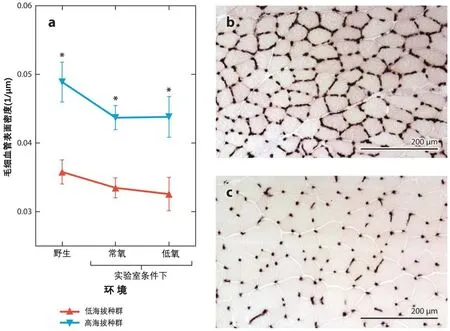
图3 高海拔鹿鼠的腓肠肌毛细血管有显著增加。(a)代表一个环境类群样本数据的偏差。在野外捕获的高原(b)和低地(c)鹿鼠的腓肠肌毛细血管。
高海拔类群的血液循环氧运输能力有所增强,循环中的氧运输强度取决于心脏输出量(心脏泵血流速)和动脉血的含氧量。后者是由血红蛋白数量(在血液中的浓度)和血红蛋白质量(氧合特性)决定。研究表明:高海拔类群在低氧环境下的心脏输出比低海拔类群更高。例如,西藏人在低氧运动过程中获得的最大心脏输出量比高原汉族人更高,即使后者在高原长期生活后也如此[13,27]。在低氧VO2max条件下,高原鹿鼠也可能具有较高的心脏输出量[114]。与此相反,长期生活在低氧环境后,高海拔类群血液中Hb含量往往低于低海拔类群,因此经常出现血液Hb含量随海拔升高而减少。
在高海拔地区,Hb-O2亲和力的调节增加了动脉血氧饱和度,从而补偿了动脉血氧饱和度的降低,提高了循环氧输送能力。这种变化可以通过内在Hb-O2亲和力的变化、Hb对红细胞内的变构辅助因子(如H+、CO2、Cl-离子和有机磷酸盐)的影响或这些辅助因子的细胞浓度变化来介导[105,108]。对比研究已经提供了强有力的证据,在哺乳动物和鸟类,增加的HB-O2亲和力有助于低氧适应[25,68,70,71,83,100,109,115,122]。例如,通过增加内源性氧亲和力和抑制对变构辅因子的敏感性,高海拔鹿鼠进化出更高的HB-O2亲和力[47,67,69],缺氧时动脉血氧饱和度也高于低氧环境[114]。总体而言,高海拔人增加的循环O2运输能力是由于最大心输出量的适应弹性和进化遗传性的增加,以及向增加动脉血氧饱和度和Hb-O2亲和力的进化。
在许多高海拔类群中,氧气在肺的气-血界面扩散的能力增强,通过增加摄取到血液中的O2,肺的O2扩散能力的增加也有助于提高低氧时动脉血氧饱和度。由于总肺体积和/或肺泡表面积的增加,高原哺乳动物和鸟类的肺扩散能力似乎增强[6,60,77]。这些差异可以通过弹性来解释,因为在早期生活中低氧[11,40,41]和成年期[19]可以诱导肺的生长和/或重塑现有的肺结构。肺结构的进化变化是否也有助于提高高海拔类群的氧扩散能力尚不清楚,一些研究显示:肺结构的变化有遗传因素[88,121],也有环境因素的诱导[6,53]。因此,低氧诱导可以增加肺的O2扩散能力。
与O2运输途径中的其它步骤相反,在高海拔的人类或一些脊椎动物中,肺通气增加的证据很少。例如,在高海拔的藏族和安第斯土著人中,即使低海拔居民很好地适应了高海拔环境,运动期间的总通气量也低于低海拔类群。由于高海拔人的肺具有高的O2扩散能力,抵消了低通气率,因此他们的肺具有更高的气体交换效率[118,123]。高原生理研究还显示:高原人类的绝对肺通气量较低,空气对流需求较低(运动期间总通气量和VO2max的比值)[6],是高原人特有的发育弹性和遗传性[8,9]。还有其它变化(如肺氧扩散能力的增加)会充分增强从空气中向血液中摄取氧气的能力,从而使高原人呼吸减少,将原本支持呼吸肌肉的代谢所需的部分氧气转移给其它组织。较低的肺通气速率也可以减少呼吸失水,并将血液二氧化碳/酸碱平衡的扰动降至最低[80,123]。然而,除以上这些,人们对高原人的其它生理方面知道很少。
总之,垂直梯度上VO2max的显著变化是由O2运输途径的各个步骤的弹性变化和进化遗传引起。对高海拔鹿鼠的研究表明,在运动肌肉(在产热和运动过程中氧气需求的主要部位)中,氧扩散能力和氧气利用能力的增强有助于增加VO2max,目前尚不清楚这些适应弹性是否会导致这些肌肉表型的任何显著变化。适应弹性和进化遗传增强的循环氧输送能力和肺氧扩散能力有助于增加VO2max值[92,94,117]。但肺通气量的增加等高原适应并没有保持高原人口繁育,因此这些高原适应特征存在补偿特征。
3.2 逆海拔梯度模式与不适应弹性的衰减
前面讨论的都是适应低氧的生理弹性,有利于适应高海拔环境[112],然而,还有一些高原低氧反应是遗传性的不适应。例如,有些常见的慢性低氧反应明显是病理性的,并加剧高原病,如人类的慢性高原病和牛的牛皮癣病[22,43,64,85,111]。高原原住民已经进化出许多机制,减弱了许多高原低氧的不适应反应,使一些主要表型逆海拔变化有两个研究。
低氧性肺动脉高压(HPH)是特别严重的高原病[22,98,113],当肺动脉在缺氧时收缩,然后重塑、变厚,随着时间的推移变得不可扩张[98]。肺血管的收缩在低海拔情况下是有益的,目的是匹配通气和血流的区域变化。整个肺的通气和氧气水平的适度变化是正常的,血管收缩或扩张与氧气供应(通气-灌注匹配)有关,增强了空气-血液界面气体交换的效率。然而,这种收缩在高海拔是适得其反的,因为整个肺部缺氧,并且肺动脉血管的整体收缩引发血管变化,最终在HPH中达到顶峰[22,98,113]。HPH可通过损害氧摄取并导致肺水肿、右心室肥大和心力衰竭而危及生命[113]。然而,一些高海拔物种表现出很少甚至没有HPH[26,32]。高原鹿鼠在慢性缺氧环境下不出现右心室增厚,与处于相同条件下的低海拔小鼠相比,它们表达了已知不同的参与心肌肥大的基因[116]。高海拔地区应防止低氧诱发的肺动脉血压升高,从而使HPH的发生率及其病理效应降至最低。
在人类和其它低地哺乳动物中,对严重低氧的适应反应涉及血液Hb浓度的增加。这种增加最初是由血浆容量减少引起的,血液中红细胞生成素增加刺激红细胞数量增加[99]。血液Hb浓度的增加似乎是对缺氧的有益反应,因为它增加了血液O2承载能力,然而,与此相关的血液黏稠度增加会产生更高的外周血管阻力,从而降低心脏输出量。因此,在高原藏族中,运动引起的VO2max与血液Hb浓度呈负相关[102,119]。同样,人类在高海拔地区怀孕期间,血液中Hb浓度的增加与死胎、早产和出生体重轻的发生率增加有关。以上这些似乎都源于子宫胎盘血液循环中的血流量减少[66]。低氧诱发的红细胞增多症在慢性高原病的发病机制中也起着重要作用,许多高海拔地区的本地人没有表现出持续升高的血液Hb浓度[2,5,58,111]。由于慢性低氧的红细胞生成反应迟钝或血浆体积的补偿性增加,从而产生逆海拔梯度模式。例如,在慢性低氧条件下,与白足鼠(Peromyscus leucopus)相比,高原鹿鼠的血红蛋白浓度显著降低(见图4)。同样,在海拔约4000m的藏族人群中,血液中的血红蛋白浓度出人意料的低,一般都在海平面人群的范围内。藏族人的Hb浓度的这种钝化增加与低氧诱导(HIF)因子低有关[101]。改变HIF通路中基因表达的非编码改变,如EPAS1和EGLN5可能重新校准低氧对红细胞生成反应的稳态设定点。或者,Hb浓度降低可能是HIF偶联调节性变化引起的与通气、心血管功能或肌肉扩张能力有关的其它性状的遗传变化的次要结果[104,111,119]。

图4 在相同慢性低氧条件下,高原鹿鼠(Peromyscus maniculatus)的血液血红蛋白(Hb)浓度增加很少,而低海拔白足鼠(Peromyscus leucopus)增加很多。
3.3 低氧化学反射弹性的演化:高原生理调控系统的变化
调节体内稳态的生理系统十分复杂,其中,化学反射是由化学感受器接受化学刺激的一类本能反射,而低氧化学反射是由于血液O2水平降低导致调节肺O2摄取和循环O2运输(见图5)[43,51,56]。这种反射是由颈动脉体感受到动脉血PO2降低引起,颈动脉体是位于颈动脉上的主要氧化学感受器[57,82]。颈动脉体是由神经支配,当被刺激时,它们将信息传送到脑部区域调节呼吸肌和增加肺活量称为低氧通气反应(HVR)[43,81]。颈动脉体刺激还激活交感神经系统(SNS),导致儿茶酚胺(肾上腺素、去甲肾上腺素等)从肾上腺髓质释放到血液循环中。这种缺氧的交感-肾上腺反应通过增加心脏输出量调节循环,包括引起许多组织中血管收缩和增加全身血管阻力,其净效果是血流优先向核心缺氧敏感器官(如脑和心脏)重新分配[34,43]。交感-肾上腺反应是急性重度低氧存活的关键[97,103],但在高原慢性低氧时,它会变得有害,因为会导致血压升高,心脏负荷增加。阻碍血液流向一些组织器官(包括消化器官和生殖器官)[4,12,34]。事实上,慢性低氧可导致长期保持或增强低氧化学反射。低氧适应(VAH)发生在低氧条件下几天到几周之后,导致颈动脉体低氧敏感性增加引起呼吸进一步增加[75,81],以及大脑对来自颈动脉体传入信号的敏感性增加[73,74]。慢性低氧时,交感神经肾上腺激活也持续存在[4,12,34]。因此,慢性低氧对低氧化学反射的影响引起了一些有益于肺氧摄取和组织O2供应(例如,通气增加)的生理反应,但其它反应(例如,持续交感神经肾上腺激活)会影响O2对某些组织的供应。
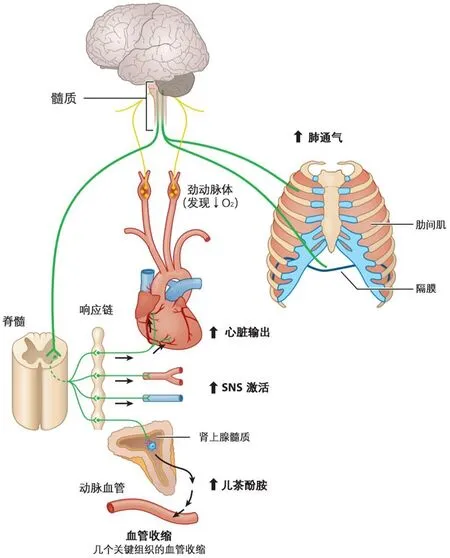
图5 低氧化学反射是调节低氧通气和循环反应的关键调控系统。颈动脉体检测到动脉血中氧分压(PO2)降低,从而增加传入神经元(黄色)向大脑发送的信息,进而改变支配呼吸肌(膈肌和肋间肌)的传出神经元(绿色)的输出,从而导致肺通气增加,并激活交感神经系统(SNS)的节前神经元。后者增加神经节后SNS神经元的活性,导致儿茶酚胺(肾上腺素、去甲肾上腺素等)从肾上腺髓质(肾上腺的内部组织)释放到循环中。这种对低氧的交感-肾上腺反应会增加心脏输出量,引起许多组织血管收缩,而一些重要的低敏感器官(如脑和心脏)会血管扩张。
保持或增加反射应该有益于肺氧摄取,但是钝化反射的影响会减弱慢性低氧对心血管系统和一些组织血流的供给。这种关系可以解释为什么低氧化学反射控制呼吸会在不同海拔的类群中出现不同的高原反应状态。一些高海拔哺乳动物和鸟类的肺通气和HVR的敏感性与低海拔同类相似或增强[3,7,42,65,78,93],但其它方面却显得迟钝[9,52]。关于低氧对交感神经肾上腺反应的影响尚不清楚,但慢性低氧时交感神经肾上腺系统的激活在高原生命较弱[4,79,87]。高海拔鹿鼠的研究显示:随着呼吸调节的发展,高原低氧化学反射弹性显著衰减,有助于稳定肺氧摄取。
在慢性低氧条件下,低地鹿鼠明显改变呼吸,而高原鹿鼠不会。低地鹿鼠表现出VAH,包括在慢性低氧条件下增加肺通气,并使呼吸模式更有效(在总通气量恒定下,更高的潮气量和更低的呼吸频率),与低氧条件下动脉O2饱和度的改善有关[44,45,46]。低海拔鹿鼠的VAH与颈动脉体的显著生长有关[44],其它哺乳动物也有同样情况[75,81]。相反,高原鹿鼠不表现出VAH,但表现出与低氧环境下低海拔类群相似的固定呼吸量增加,并且在缺氧条件下维持较高的O2饱和度波动[44,46]。高海拔鹿鼠肺泡通气量和肺氧摄取量有固定增加,这与VAH反应的遗传同化(环境诱导的表型变成遗传型的过程)一致,同时颈动脉体没有扩大,这是低地生命VAH的典型表现[44]。后一个性状很重要,因为颈动脉体的扩大会增加大脑的传入信号,从而激活交感神经系统。
高海拔鹿鼠肾上腺髓质中儿茶酚胺分泌显著减少,高原鹿鼠的儿茶酚胺分泌比低地同种类要低得多,主要是儿茶酚胺合成减少所致。另外,慢性低氧降低了高原鹿鼠肾上腺髓质对下游刺激的敏感性,人没有这种现象,因此,高原鹿鼠颈动脉体对大脑的传入信号越弱,分泌到循环中的儿茶酚胺就越少。海拔越高,分泌儿茶酚胺越少,高原反应就越重[87]。
高原鹿鼠的低氧化学反射已经具有在慢性低氧条件下保持O2稳态的能力。在复杂的稳态调节系统中,一个调节表型是:O2运输(呼吸和肺氧摄入量增加),表现出适应高原条件;另一个调节表型是:交感肾上腺激活的衰减,不适应慢性低氧。慢性交感神经激活是VAH弹性的一个重要标志,通过其它机制诱导增加固定呼吸和肺氧摄取量,可以避免或减弱这种不适应。增加固定呼吸和肺氧摄取量会通过低氧化学反射钝化交感神经肾上腺激活。因此,生理调控系统的弹性能导致适应和不适应的表型分化,也是类群分化的生理基础。
4. 全球气候变化、表型弹性和海拔变化
随着全球变暖,低地物种活动的海拔范围会上移,导致高山和亚高山物种竞争性迁徙。低地物种上移的情况取决于对环境和低氧的适应模式。人类和哺乳动物的低氧研究表明:表型弹性限制了高海拔适应。在高处,慢性低氧引起的直接适应与不适应机制都在演化。
