云南榧树群落种内与种间竞争研究
2022-02-10于达勇樊智丰马长乐李瞒瞒
于达勇,樊智丰,马长乐,2*,李瞒瞒
(1.西南林业大学 园林园艺学院,云南 昆明 650224;2.国家林业和草原局 西南风景园林工程技术研究中心,云南 昆明 650233)
植物体间的种间与种内竞争现象在自然群落中普遍存在。该现象表现为:在同一生长空间内相邻个体间为了最大限度地获取有限的资源(即水、光和养分),以牺牲其他树木的资源可用性为代价,对有限资源和空间进行争夺,最终阻碍到竞争双方的正常生长和发育[1-2]。种内和种间竞争现象是影响植物生存与发育、物种丰度和分布、群落结构与稳定性以及生态系统功能与健康的主要因素[3-4],能够推动群落的自然演替进程[5]。对植物种内种间竞争的研究一直是研究植物种群生态的核心问题。近年来,已有不少对濒危植物竞争状况的研究,如南方红豆杉(Taxuswallichiana)[6]、梓叶槭(Acercatalpifolium)[7]、太山柳(Salixtaishanensis)[8]等。研究发现,这些濒危植物种群的他疏现象大于自疏现象,受到周边树木的影响较大,不能很好地与其他植物共存。若要真正做到与周围物种稳定共存,群落中各物种必须对自己的限制超过竞争对手,这样双方才不会被对方完全接管[9]。
云南榧树(Torreyayunnanensis)是红豆杉科(Taxaceae)榧树属(Torreya)植物,国家二级保护野生植物,其分布区域较为狭窄,主要分布于滇西北横断山脉地区[10]。由于人为破坏严重和自然更新较慢,数量逐年锐减,已被《中国物种红色名录》确定为濒危物种[11],研究其群落竞争对该树种的保护和可持续利用具有重要意义。本研究采用Hegyi单木竞争模型对云南榧树群落的种内、种间竞争强度以及其与对象木径级间的关系进行定量分析,探索其竞争机制并预测该种群动态发展,为云南榧树种质资源保护和可持续经营提供科学依据。
1 研究区概况
研究区域位于云南省维西县和香格里拉市(25°04′50.11″-27°48′7.88″N,99°01′32.62″-99°29′39″E),海拔1 771~2 690 m,群落多位于林地的下位坡或山谷之中,坡度在30°~40°,属于亚热带与温带季风高原山地气候。该地区地势复杂且温、光、水分布不均匀,年平均气温15.3 ℃,最低温度-1.5 ℃;年日照总时数为2 104.5 h;年平均降水量938.1 mm,降雨日数100~160 d,年平均风速1.3 m·s-1,年平均降雪11 d,年平均霜期169 d。研究地内植被类群丰富,以高山针阔叶混交林为主,常见树种有川滇高山栎(Quercusaquifolioides)、木蜡树(Toxicodendronsylvestre)、云南樱桃(Cerasusyunnanensis)、飞蛾槭(Aceroblongum)、青冈(Cyclobalanopsisglauca)等。
2 研究方法
2.1 数据采集
在对云南榧树主要分布区进行充分踏勘后,选择具有代表性的地段设置7块30 m×30 m样地进行调查。记录各样地的经纬度、海拔、坡度、坡向等基本信息,以样地的水平边与垂直边为参照建立坐标体系,调查样地内胸径大于等于5 cm木本植物的物种、胸径、树高、冠幅以及在样方中的相对坐标值。最后建立该区域的植被信息数据库,根据对象木与竞争木的坐标值,计算出竞争木与对象木之间的距离Lij[12]。
2.2 竞争指数的确定
目前,对群落内植物邻体间的竞争模型有很多,Hegyi单木竞争指数模型运用最为广泛,它利用对象木与竞争木的胸径比和距离,能较准确地反映出林木之间的竞争状况[13]。故本研究采用Hegyi单木竞争模型对云南榧树群落进行分析,其公式如下[14]:
(1)
式中:CI为竞争指数;Dj、Di为竞争木胸径和对象木胸径;Lij为对象木与竞争木之间的距离;N为竞争木的数量。竞争指数CI越小,对象木与竞争木之间的竞争越弱。通过该公式先求出单个竞争木对对象木的竞争指数,再将不同单木竞争指数进行分类与累加即为云南榧树的种间和种内竞争指数。
2.3 最佳竞争范围的确定
在样地内所有胸径大于等于5 cm的对象木中选择20株并以此为中心,采用逐步扩大竞争半径的方法,分别计测不同竞争半径为2、3、4、…、30 m时样圆内竞争木的平均竞争强度,做出不同竞争半径与竞争强度间的拟合关系图,以竞争强度变化不明显时的半径作为最佳竞争半径。
2.4 数据分析
根据野外调查所测定的植物数据,使用AutoCAD 2014绘制样地植被分布图,计算对象木与竞争木之间的距离;使用SPSS 20.0进行数据的分析和回归预测,Origin 85进行图形的绘制。
3 结果与分析
3.1 云南榧树群落邻体间最佳竞争范围的确定
为了保证研究结果的准确性和可靠性,需要确定真正的具有明显生态效应竞争木的作用范围[15]。根据回归结果可见(图1):当竞争半径处于0~10 m时,平均竞争强度呈明显下降趋势。而大于10 m后,随着范围增加和竞争木数量增加,一些竞争效应较小的竞争木便会将有效竞争木的作用掩盖掉,从而使得平均竞争强度较低。因此以半径10 m范围内的植物为竞争木能较好地反映云南榧树的竞争情况。
3.2 云南榧树种群径级特征
共调查云南榧树85株,胸径范围5~185 cm,平均胸径为25.3 cm。根据曲仲湘等[16]立木分级标准对云南榧树胸径进行分级,结果显示云南榧树径级分布不均匀,以中大型树木为主。其中中树最多,占据总数的47%以上,其次为大树和小树,说明云南榧树的年龄结构为稳定性。伴生树种共调查177株,胸径范围5~135 cm,平均胸径22.3 cm,也是以中树为主,占总数的41%以上(表1)。可见云南榧树的胸径略大于周边伴生树种但差距不大,说明该群落中云南榧树在径级方面略优势于其他树种。在对比云南榧树与伴生树种的平均树高后发现,同种径级阶段下云南榧树的树高均小于伴生树种,还发现云南榧树处于大树阶段的平均树高才达到伴生树种的平均树高(9.29 m)水平,即在树高方面,云南榧树只有成年后才能占据树木的中上层。
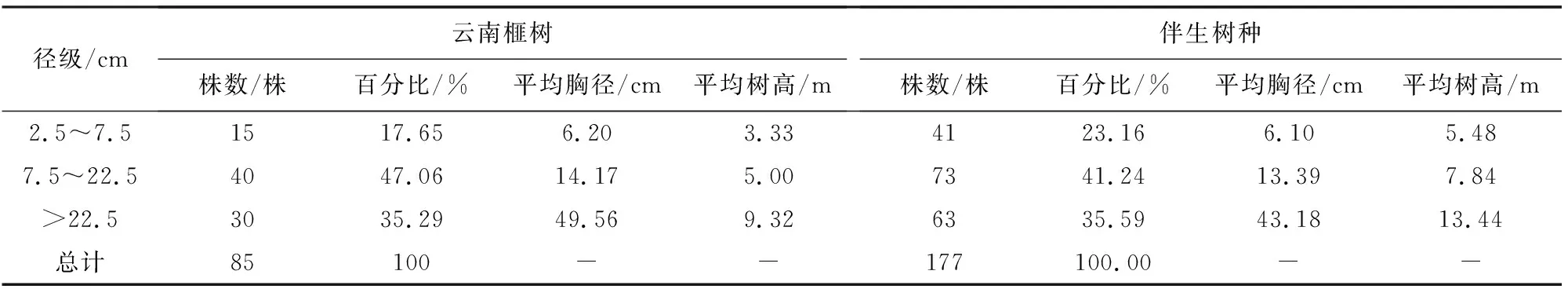
表1 对象木与竞争木基本信息Table 1 Basic information of object tree and competitive tree
3.3 云南榧树种内竞争强度
对云南榧树受到的种内与种间竞争强度进行对比分析(图2、图3),发现云南榧树的总竞争强度为579.34,其中种内竞争强度为330.44,占总竞争强度的57.04%;而种间竞争强度为248.90,占总竞争强度的42.96%。这说明云南榧树在发育过程中受到的种内和种间竞争大致相等,并且种内竞争略大于种间竞争,云南榧树不会因竞争劣势而被其他树种所替代。对不同径级下云南榧树受到的竞争强度研究发现,随着云南榧树径级的增加,受到的种内竞争强度逐渐减小。径级在5~20 cm的榧树受到的种内竞争较为激烈,竞争强度总计270.08,占总种内竞争的81.7%。其中径级在5~10 cm的竞争强度高达128.34,在全部径级中受到的种内竞争最为强烈。径级在5~30 cm的过程中种内竞争整体呈现为稳定的下降趋势,但由于30~35 cm中的一些榧树受到局部高大榧树的激烈竞争,导致局部数据出现变大现象。当径级大于40 cm时,云南榧树的竞争强度仅为3.43,并在此之后便趋于稳定。结果表明,正在生长的小树更容易受到较多的种内竞争,随着云南榧树径级的不断增大受到的种内竞争会逐渐减小最终趋于稳定,但若生长到过于高大,离其他小树过近,则会对它们造成强烈的竞争,影响其他榧树的生长。
3.4 云南榧树种间竞争强度
对不同径级下云南榧树所受到的种间竞争强度进行分析发现,云南榧树受到的竞争压力主要集中在幼苗阶段,其中径级在5~10 cm的榧树受到的种间竞争最为强烈,竞争强度高达174.42,在种间竞争中处于劣势地位。但径级达到10 cm后,受到的种间竞争会急剧下降,直到径级大于25 cm后,竞争强度降至极低点2.33,且保持稳定,此时云南榧树受到竞争木的影响较小,说明小树在生长过程中容易受到其他树木的剧烈排斥,若顺利度过幼年时期便能占据群落中的优势地位。
云南榧树受到种间竞争不仅与自身径级相关,还会因竞争植物种类的不同而存在差异。云南榧树所在群落物种丰富(共45种),与榧树发生竞争关系的有37种,种间竞争的总竞争强度为248.90。不同种对榧树的影响差异较大,云南榧树种间竞争强度由大到小顺序依次为:川滇高山栎、胡桃、飞蛾槭、栗(表2)。竞争强度最高的是川滇高山栎,竞争指数为68.42,其次为胡桃与飞蛾槭,竞争指数为25.42和22.13,在群落中对云南榧树的竞争压力最大,其中川滇高山栎和飞蛾槭为当地常见树种,数目较多,较容易与云南榧树产生竞争关系,而胡桃树在样地中虽然个体数量少,但由于胸径较大且离对象木较近,因此也对云南榧树造成了较大的竞争作用。

表2 不同种类竞争木的竞争强度Table 2 Competition intensity of different kinds of competitive trees
3.5 云南榧树胸径与竞争强度关系及其预测
植物间竞争受到个体大小与生长情况、植株间的距离、自然环境等因素的影响,其中胸径大小对竞争的影响较大,因此以胸径为自变量、竞争指数为因变量进行非线性回归模拟,发现竞争指数与云南榧树的胸径之间呈负相关并服从幂函数关系,即CI=AD-B。式中,CI为竞争指数,D为对象木的胸径,A、B为模型参数。对云南榧树胸径与种内种间竞争的关系进行幂函数拟合,结果分别为CI种内=43.417D-1.056、CI种间=59.605D-1.408(图4、图5)。显著性水平均小于0.01,所得模型可用于预测云南榧树胸径与竞争的关系。
预测结果表明,种内和种间竞争强度均随着对象木胸径的增加而减弱。对象木在幼年时期受到较大的种间和种内竞争,10 cm处为转折点,当对象木胸径增加到10 cm后受到的种内种间竞争压力均明显减弱且逐渐趋于稳定。表明虽然云南榧树幼年时期在群落中处于劣势地位,但随着个体的不断生长,其自身的竞争能力逐渐增强,周围的竞争木对其的影响也会随之减弱,因此对云南榧树的保护应着重于小树的保护。
4 结论与讨论
采用Hegyi单木竞争模型对榧树群落进行分析,对结果影响较大的参数有:竞争木数量、对象木和竞争木的胸径以及两者之间距离,若想运用该模型表现植物对资源的竞争能力,则必须合理确定对象木的竞争范围。通过逐步扩大范围的办法[17]所确定的云南榧树有效竞争半径为10 m,比绝大多数树种的竞争半径都大。原因推测为在所调查的云南榧树群落中,对象木的胸径较大,冠幅较广,与其产生树冠相重叠的部位现象的竞争木较多,在半径10 m内的竞争木均与对象木存在不同程度的竞争关系。
对云南榧树群落种内和种间竞争进行研究发现,云南榧树的种内竞争强度略大于种间竞争强度。该结果符合经典竞争理论,即在稳定条件下,如果某物种的种内竞争比种间竞争激烈,那么具有相似要求的物种可以共存[18]。因为在群落中该物种限制自身种群增长的程度便会大于限制竞争对手种群增长的程度[19],这样群落不会被单一物种所占据,更容易保持群落的稳定。由此推测:云南榧树能与周边物种建立起共存的竞争关系,形成稳定的群落结构。在使用Hegyi单木竞争模型计算时,胡桃仅因为胸径大、离榧树近便产生了较大的竞争效应,说明采用的指标较为单一,若要真实地反映竞争情况还需要在冠幅大小、林荫重叠等方面进行改良。
除此之外,植物的竞争能力还与植物的生长阶段有关[20]。本研究中云南榧树不同生长阶段数量由大到小顺序为:中树、大树、小树,年龄结构为稳定型。对不同径级的云南榧树所受到的种内和种间竞争进行分析,发现径级较小的云南榧树受到的种内与种间竞争较大,处于竞争中的劣势地位;反之较大的榧树受到竞争却较小。此结果推测:云南榧树幼年时期,冠幅和树高较小加之为裸子植物生长速度较慢,长时间受到上层树木的遮挡和排挤,温、光、水等生存资源不足,导致该阶段植物生长困难。但随着云南榧树径级的增加其受到的种间和种内竞争逐渐下降,直至生长成中、大型林木,此时云南榧树的胸径与冠幅较大且树高也位于群落中上层,对环境资源的竞争能力大于其他竞争木,处于竞争的优势地位。再对竞争强度与对象木胸径的拟合模型进行分析,预测同样显示为:种内和种间竞争强度随胸径的增加而降低,以10 cm为分界点,胸径小于10 cm的幼苗受到种间竞争压力大,但超过10 cm后受到的竞争压力已位于较低水平且下降幅度不明显。因此对云南榧树的保护需侧重于幼年时期,采取一系列的人工抚育措施,如择伐、抚育幼苗等方式来降低云南榧树的种间竞争压力,为幼苗的生长给予充足的空间,使其顺利进入成年时期,以保障幼苗的正常成长,促进云南榧树的自然更新。
致谢:感谢西南林业大学邓莉兰教授在标本鉴定方面给予的帮助,感谢维西县林业局高级工程师张海涛在外业调查上提供的帮助,感谢王珞娴、牟一丹和闻楷英同学在数据处理方面所做的帮助。
