不同光强下4种鸭跖草科植物的生物量分配、水分生理及光响应特征
2022-02-10薛晴陈斌杨小梅杨宇佳李子葳薄杉何淼
薛晴,陈斌,杨小梅,杨宇佳,李子葳,薄杉,何淼
(东北林业大学园林学院,黑龙江 哈尔滨 150040)
自然界中,诸多环境因素(光照、温度、水分以及CO2等)均会对植物的生长发育产生重要影响[1],其中光照因素作为重要的环境因素之一,为植物的生长提供能量来源,能够从多方面影响植物的结构特征和生理过程[2]。园林植物的生长同其他植物一样,需要依赖于周边环境、光照自然环境,其最重要的属性之一就是观赏特性。一般来说,不适宜的光照条件会造成植物叶片失绿、花叶弱化、徒长变弱等现象,使得观赏价值降低的园林植物很难完成美化环境的任务,故园林植物往往比其他植物更需要适宜的光照条件。适宜的光照条件可以促进其生长良好,进而产生更高的观赏价值和生态效益,这无疑对城市绿化具有重要意义[3]。然而自然界中的光环境是复杂多变的,这使得不同生活型的植物在经过漫长的生物进化过程后,形成了一定的适应性调节策略。当植物的生长环境光因素匮乏时,其能够从生物量分配、水分利用以及光响应特征等多方面做出调整,以规避或减弱光环境变化产生的伤害。研究表明,植物会采用增加干物质重量、减少根冠比等生物量分配方式来适应弱光环境[4]。马天光等[5]发现骆驼刺(Alhagi sparsifolia)在应对光强不足的环境时,主要依靠的是牺牲对水分胁迫有较好抗逆性的水分生理特征,并通过消耗更多的水分去维持植物生长所需的基本光合能力。此外,耐阴植物能够通过降低光补偿点(light compensation point,LCP)、光饱和点(light saturation point,LSP)以及暗呼吸速率(dark respiration rate,Rd),和提高表观量子效率(apparent quantum yield,AQY)等方式来维持弱光环境下的稳定生长[6]。近些年来,国内外关于植物对不同光强的生理生态响应主要集中在林用树种和蔬菜作物上[7],对园林植物响应光强变化的研究较少,因此,研究不同光环境下的园林植物响应策略,有助于阐明不同光环境下园林植物的生理生态适应性,为园林植物更好的应用于城市绿化奠定基础。
近年来,生态环境保护受到高度重视,生态文明建设被纳入国家发展总体布局,表明了实现人与自然和谐共生的现代化美丽中国既是目标也是使命。随着“美丽中国”建设的推进,对园林绿化投资与建设的力度也在不断加大。园林植物作为城市绿化中主要构成因素之一,承担着绿化美化以及发挥生态效益的重要作用[8],然而当前城市建设的迅速发展造成城市荫蔽空间越来越大,因此选育观赏价值高且兼具耐阴的植物资源对荫蔽空间进行科学有效的绿化,发挥绿地最佳的生态环境效益具有重要意义[9]。鸭跖草科(Commelinaceae)植物繁殖容易,管理粗放,大部分种类叶色绚丽,花朵繁茂,是理想的观花观叶且适用作地被的植物材料,具有广阔的应用前景[10]。然而目前对于鸭跖草科植物的研究多集中在中药成分、污染防治以及黄酮类物质含量上[11—12],对其在不同光环境下的响应机制研究较少。因此,本试验欲以4种鸭跖草科植物[紫鸭跖草(Commelina purpurea)、‘花叶’水竹草(Tradescantia fluminensis‘Variegata’)、吊竹梅(Tradescantia zebrina)、‘绿叶’水竹草(Tradescantia fluminensis‘Vairidia’)]为供试材料,通过人工模拟5种光强梯度,测定不同光环境下鸭跖草科植物的生物量分配、水分生理以及光响应特征,以期揭示4种鸭跖草科植物在不同光强下的响应规律,为鸭跖草科植物科学有效地应用到城市绿化建设中提供科学依据。
1 材料与方法
1.1 试验材料
2019年7月,选取紫鸭跖草(C.purpurea,代号‘CP’)、‘花叶’水竹草(T.fluminensis‘Variegata’,代号‘TF’)、吊竹梅(T.zebrina,代号‘TZ’)、‘绿叶’水竹草(T.fluminensis‘Vairidia’,代号‘TV’)为供试材料,扦插苗生长在温度24~26℃,相对湿度60%~70%,每天12 h光照,白天光强480μmol·m—2·s—1的育苗室内,每种试验材料60盆,一个月后,选取长势良好、生长健壮的植株进行试验。
1.2 试验设计
试验在黑龙江省哈尔滨市东北林业大学花卉研究所苗圃进行,将不同密度的黑色尼龙遮光网覆盖在事先搭好的木制遮光架上,遮光网距离植株体顶端30 cm,共设置5种(L0、L1、L2、L3、L4)光强梯度(遮光度分别为0、25%、50%、75%、95%),每个处理间隔50 cm,在每个光强梯度下,每种植物材料分别处理12盆,设置3个重复。每隔5 d浇一次水,处理45 d后,每个处理每种植物随机选取3盆进行各项指标的测定。
1.3 测定指标
1.3.1生物量测定 在测定完各项试验指标后,采用慢速流水冲刷的方式,从每个处理组中小心的挖出6个完整植株,尽可能地减少根系的损伤。将每一株的根、茎、叶分离,放入60℃烘箱中烘干至恒重,并使用千分之一电子天平(UX420H,天津)进行称重。相关指标计算公式如下:
1.3.2叶片相对含水量和失水速率的测定 2019年7月,摘取植株顶部第3~5片完整无病虫害的叶片,迅速保存于放有冰袋的带盖保温盒内,带回实验室立即进行相关参数测定。随机选取不同处理的植株叶片用千分之一电子天平进行称重,得到鲜重W1。然后将叶片完全浸泡在装有蒸馏水的烧杯中,密封好后在黑暗处进行饱和吸水,24 h后取出,擦干叶子表面多余水分后再次称重,得到饱和鲜重W2。然后将叶片放入105℃烘箱杀青30min,再60℃烘干至恒重,称重得到叶片干重W3,每个处理3次重复。叶片相对含水量(relative water content,RWC)计算公式如下:
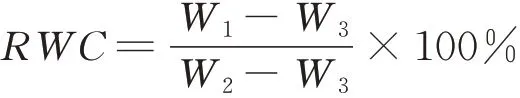

同时再次随机选取不同处理的植株叶片用千分之一电子天平进行称重,得到鲜重W1(此时记为T0)。然后将叶片置于室温(25℃)下进行自然失水12 h,期间每隔1 h进行称重,得到实时鲜重W i(时间点记为T i),最后将叶片放入105℃烘箱杀青30 min,再60℃烘干至恒重,称重得到叶片干重W2。叶片失水速率(rate of water loss,RWL)计算公式如下:
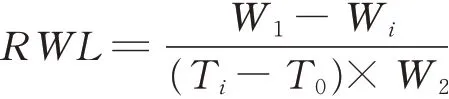
1.3.3水势的测定 2019年7月,选择晴朗无云的天气,每组处理中每种植物各选取3株进行试验,每个处理3次重复。选择每株植物中上部的1个二级分枝,用PMS压力室(PMS,美国)进行带叶小枝的水势测定。清晨水势于4:30(北京时间,下同)测定,正午水势于14:00测定。
1.3.4光响应曲线的测定和拟合 2019年7月,选择晴朗无云的天气,在上午9:00—11:00,采用Li-6400XT便携式光合仪(美国),对每组处理中选取的不同种类的3株植物叶片进行光响应曲线的测定。利用光合仪中红蓝光源的标准叶室设定光合有效辐射(photosynthetic active radiation,PAR),从低到高光强梯度设定为0、20、50、100、150、200、400、600、800、1000、1200、1400、1600、1800、2000μmol·m—2·s—1,叶室温度为25℃,相对湿度控制在60%~70%,CO2浓度控制在400μmol·mol—1,记录不同处理下每种供试材料的净光合速率(net photosynthetic rate,Pn),并读取胞间蒸腾速率(intercellular transpiration rate,Tr),水分利用率(water use efficiency,WUE)计算公式如下:
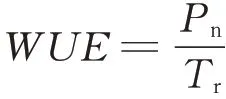
利用Pn数据,采用非直线双曲线模型进行拟合,计算如下:
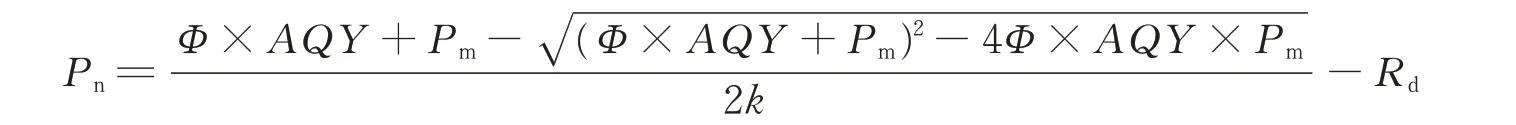
式中:Pn为净光合速率,AQY为表观量子效率(apparent quantum yield),Φ为光合有效辐射,Pm为最大净光合速率,k为光响应曲线曲角,Rd为暗呼吸速率,选用当AQY值为0、20、50、100、150、200μmol·m—2·s—1时得到的净光合速率(Pn),运用Excel软件进行线性回归分析,计算出最大净光合速率(Pm)、光补偿点(light compensation point,LCP)、光饱和点(light saturation point,LSP)以及表观量子效率等,多组数据测定与分析取平均值。
1.4 数据处理
利用Excel 2016整理与计算数据,利用SPSS 22.0进行数据分析,用Duncan法进行多重比较(P<0.05),并对处理结果进行双因素方差(ANOVA)分析。
2 结果与分析
2.1 不同光强对生物量的影响
从图1可以看出,随光强的降低,4种鸭跖草科植物的地上生物量均呈现出先升高后降低的趋势。CP、TZ与TF的地上生物量均在L2时达到峰值,L4时达到最小值,与最大值相比,分别降低了21.96%、67.69%、73.45%;TV在L1时达到峰值,L4时达到最小值,显著降低了50.00%(P<0.05)。而对于总生物量,除TZ外,CP、TF与TV的总生物量也呈现出先升高后降低的趋势。CP、TV在L1时达到最大值,L4达到最小值,与最大值相比,分别降低了46.26%、59.18%;TZ、TF在L2时达到峰值,L4达到最小值,与最大值相比,分别降低了69.30%、76.72%。可见强光与极弱光都会对4种植物的生长产生抑制,4种植物也均能通过增加地上生物量的方式来应对光强的减弱。CP在所有光强下,地上生物量与总生物量均无显著变化,表明其对光强的适应性较强。
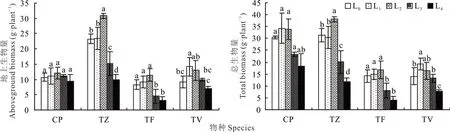
图1 不同光强对生物量的影响Fig.1 Effect of different light intensities on biomass
2.2 不同光强对物质分配规律及冠根比的影响
不同光强对4种鸭跖草科植物的物质分配规律及冠根比的影响不同(图2)。CP的根贡献率最小值出现在L4光强条件下,与L0相比,显著降低了26.15%(P<0.05);茎贡献率在L4达到最大值,与L0相比,显著增加了86.67%(P<0.05);叶贡献率在各处理间均无显著变化。而TZ的根与叶贡献率在各处理间均无显著变化;茎贡献率在L4达到最大值,与L0相比,显著增加了32.26%(P<0.05)。TF的根贡献率最小值出现在L4光强条件下,与L0相比,显著降低了47.62%(P<0.05);茎与叶贡献率均在L4达到最大值,与L0相比,分别增加了39.29%、33.33%。TV的根贡献率最小值出现在L4光强条件下,与L0相比,显著降低了70.59%(P<0.05);茎贡献率在L4达到最大值,与L0相比,显著增加了52.63%(P<0.05)。

图2 不同光强对物质分配规律及冠根比的影响Fig.2 Effect of different light intensities on mater ial distribution and crown r oot r atio
除CP外,TZ、TF与TV的冠根比均呈现出“升高—降低—再升高”的变化趋势,但四者均在L4达到最大值,并与L0相比,分别显著增加了94.55%、94.21%、160.56%、345.05%(P<0.05),TZ、TF与TV均在L3时开始呈现下降趋势,这可能是由于在L3条件下,三者的根贡献率较高,而茎与叶贡献率相对较低造成的。可以看出4种鸭跖草科植物能够在生物量分配方面做出调整以适应不同的光强环境,具体表现为增大茎、叶贡献率以及冠根比。
2.3 不同光强对叶片水势与相对含水量的影响
从图3可以看出,4种植物的叶片含水量变化虽然呈现出不同的趋势,但是与L0相比,其他4种处理下的植株叶片含水量均有不同程度的升高,TZ与TV均在L2条件下达到最大值,与L0相比,分别增加了5.74%、2.04%;而TF在L3条件下达到最大值,与L0相比,显著增加了2.83%。可以看出遮阴环境能够提高4种植物的叶片相对含水量,从而提高其保水能力。
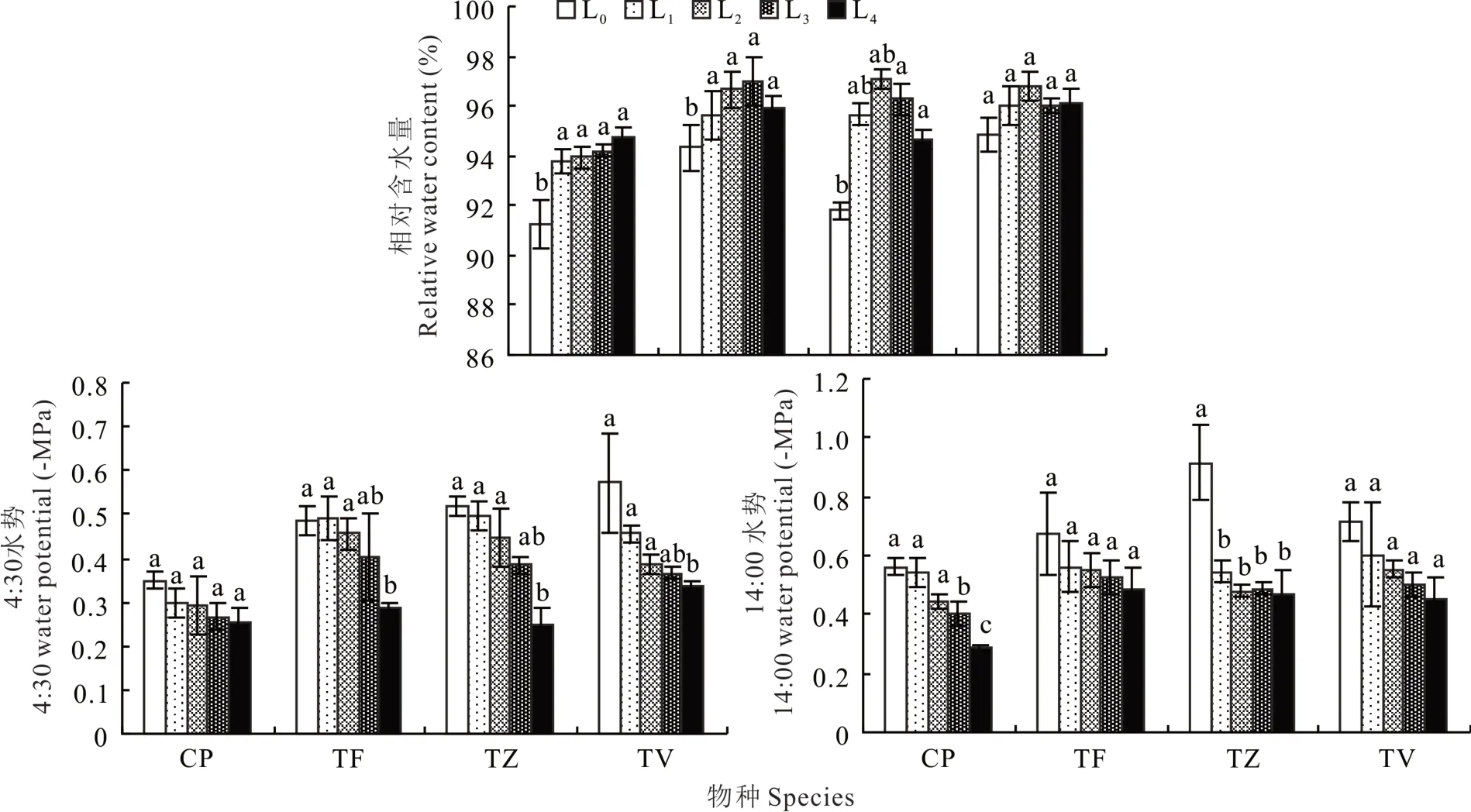
图3 不同光强对叶片水势及相对含水量的影响Fig.3 Effect of different light intensities on water potential and relative water content
4种植物的正午水势变化较清晨水势变化更加显著,变化幅度更大。其中CP、TZ与TV的清晨水势与正午水势变化趋势相同,均呈现上升趋势。4种植物均在L0时水势最低,随着光强的减弱,水势呈现不断上升的趋势,在L4达到峰值。与L0相比,4种植物的清晨水势分别增加了28.57%、40.82%、51.92%、42.11%,正午水势分别增加了50.00%、26.87%、48.91%、37.50%。可见L0光照条件下,4种植物的水分亏缺程度较大。
2.4 不同光强对失水速率的影响
由图4可见,4种植物在L0条件下的失水速率均较低,且变化幅度较小,变化趋势较为平稳。随着光强的降低,失水速率呈现不同程度的增加,且变化幅度也有不同程度的增大,这种趋势在CP中表现得最为明显。较其他两种植物,CP、TZ的失水速率在所有处理下均具有较低水平。此外,可以发现正午水势与失水速率的变化趋势具有较高的一致性(图3和图4)。
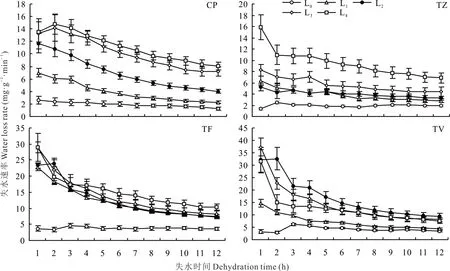
图4 不同光强对叶片失水速率的影响Fig.4 Effect of different light intensities on water loss rate
2.5 不同光强对光响应曲线的影响
由图5可知,在不同的光强处理下,4种植物的光响应曲线均呈现出随着光合有效辐射(PAR)的增加而先增加后趋于平缓的趋势,均表现为当PAR≤400μmol·m—2·s—1时,4种植物的净光合速率(Pn)有较大的增长速率,当PAR>400μmol·m—2·s—1时,4种植物的Pn逐渐达到一个平台期,变化幅度非常微小,趋近于各自的光饱和点和最大净光合速率。然而在不同处理下,4种植物的光响应情况又表现出明显差异。在L0、L1、L3与L4处理下,CP的最大净光合速率(Pm)均高于其他3种植物。此外,还可以看到当PAR>1400μmol·m—2·s—1时,4种植物在不同处理下的净光合速率会产生不同程度轻微的下降,这可能是由于光强过大而产生的光抑制现象。
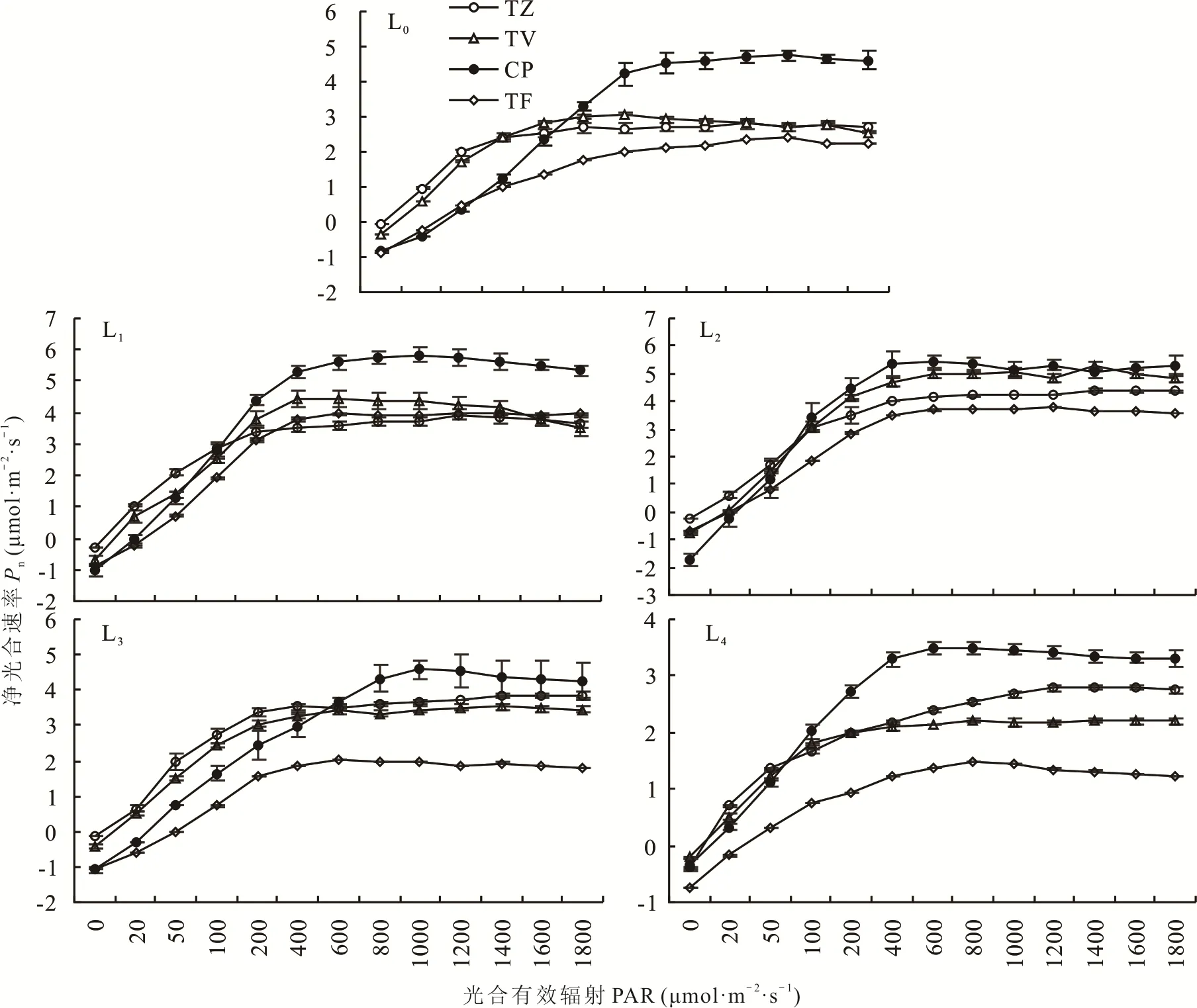
图5 不同光强对光响应曲线的影响Fig.5 Effect of different light intensities on light response curves
在植物的光合作用中,最大净光合速率能够在一定程度上衡量植物对光能的利用能力。由图5和表1可知,CP、TZ与TV均在L2条件下达到最大净光合速率,分别较L0显著增长了27.62%、67.02%、83.07%;而TF在L1条件下达到最大净光合速率,较L0增长了51.69%。可见遮阴条件可以在一定程度上提高4种植物的最大净光合速率。
2.6 不同光强对光合参数的影响
通过非直线双曲线模型拟合得到4种植物的光合参数,由表1可知,模型的决定系数(R2)均大于0.9,表明该模型的拟合效果较好,能够较好地反映出4种植物的光响应过程,可以看到不同光强对4种植物叶片的光合参数影响有所不同。随着光强的减少,CP、TF、TZ、TV的Pm均呈现出先升高后降低的趋势,其中TV、TZ的增幅较大,在L2条件下,分别较L0显著增长了83.07%、67.02%。4种植物的AQY值为0.023~0.087μmol·m—2·s—1,各物种叶片间的差异较小,但与L0相比,涨幅有很大不同,除CP在L2达到最大值外,TF、TZ、TV均在L1达到最大值,CP、TF、TZ、TV在达到最大值时分别较L0增加了165.22%、52.63%、7.50%、15.09%。TZ的LCP值偏低,为5.69~10.28μmol·m—2·s—1,在L2达到最小值,较L0显著降低了44.65%(P<0.05),而TF的LCP在不同处理间无显著差异。4种植物的LSP与Rd均存在显著差异,LSP呈先升高后降低的趋势,与Pm呈现出类似的趋势。CP的Rd偏高,在L2时达到了最大值,为1.74μmol·m—2·s—1,较L0增长了125.97%;而TZ在所有处理下的Rd均偏低,导致Pm/Rd值较大,为16.24,分别是CP、TF、TV的4.00、2.93、2.28倍。TF、TV的WUE呈现出先升后降的趋势,其中CP、TZ的WUE在不同处理间无显著差异,均表现出较高的水平。
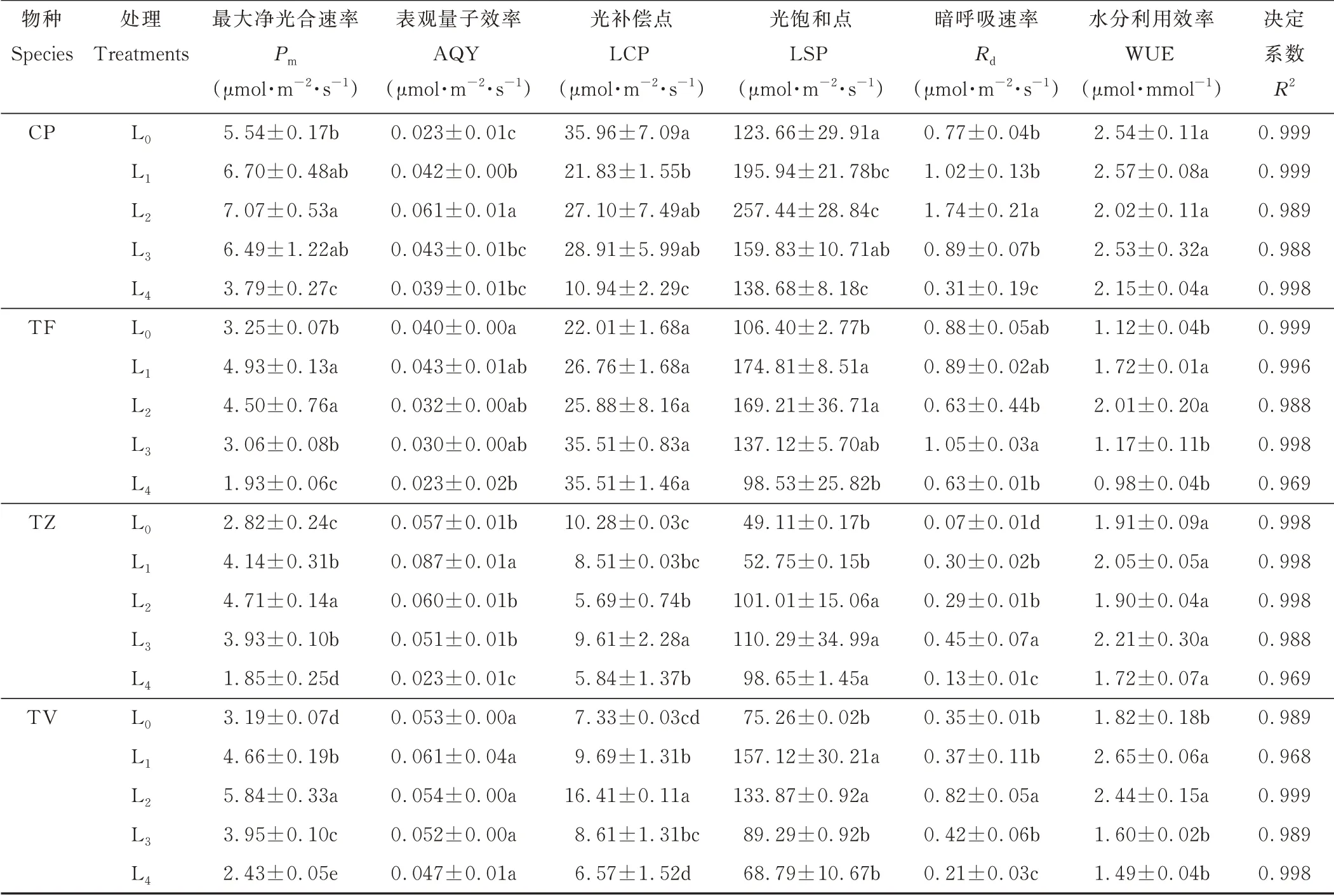
表1 不同光强对光合参数的影响Table 1 Effect of different light intensities on photosynthetic parameter
3 讨论
3.1 光强对生物量及物质分配规律的影响
光是植物的唯一能量来源,植物对光的捕获能力与利用效率直接决定着其能否很好地适应环境并维持自身的良好生长[13]。当外界光环境发生变化时,对植物的生物量及物质分配、水分生理以及光响应特征都会产生一定程度的影响。一般情况下,植物在弱光环境中,生物量、相对生长速率会变低[14—15]。而为了适应荫蔽环境,植物通常会进行生长与形态调节,即所说的形态可塑性,去适应复杂的光环境。有研究表明,当光强减弱时,幼苗的株高生长量、地上生物量会增加,以捕获更多的光能[16],本研究也得到相似的结果。随着光强的降低,4种植物的地上生物量均呈先升高后降低的趋势,一方面说明植物可以增加地上生物量去适应环境,另一方面表明4种鸭跖草科植物具有一定的耐阴性[17—18],尤其以TZ、TF的变化最为显著。一般来说,大多数植物的生物量积累都存在一个最佳光强范围[19],本研究中4种植物的总生物量均在L2达到较高水平后开始呈现下降趋势,且均在L4时达到最低,说明当遮光度超过50%时,植株物质积累速率开始放缓,当光照强度超过最佳光强范围时,植物的生物量积累开始减少,尤其在极端的弱光环境(遮光度95%)中生长会受到显著抑制,甚至导致植物死亡[20]。这表明较强或者较弱的光照环境都会对植物的生物量积累产生抑制,但适度遮光能促进生物量积累[21]。
植物干物质的分配格局及变化是其响应环境变化的重要策略表征,许多研究表明,植物可以通过改变自身干物质在根与冠之间的分配比例来适应复杂的光环境[22]。在谭淑娟等[23]的研究中,根的贡献率在强光下呈现出较高的趋势,本研究中也发现了相似的规律,其中以CP表现得最为明显。在遮光度为0%~50%条件下,CP的根贡献率没有发生显著变化,且在不同处理下均高于其他3种植物,表明CP的地下生物量较大,对光的适应性较强。相反,随着光强的减弱,TZ、TF与TV表现出较小的根贡献率、较大的叶与茎贡献率以及较大的冠根比,表明其在弱光条件下能够通过增加地上部分(叶与茎)的物质分配比例,提高对光能的捕获能力,从而弥补光能的减少,这与前人的研究结果一致[24]。4种植物的茎贡献率随着光强的降低呈现出逐渐升高的趋势,这说明4种植物在弱光下都采取了增加节间长的方式去获取有限的光资源[25],但是微弱的节间变化并不能够弥补总生物量的下降,进一步说明当遮光度达到75%或以上时,植物的响应策略并不能完全缓解其生长发育受到的抑制情况。
3.2 光强对水分生理的影响
植物的光合作用离不开水分的参与,光环境的改变会对植物体内的水分状况产生一定的影响,进而影响植物的生长发育[26]。通常情况下,水势被当作一个直接水分指标去反映植物的水分状况和水分亏缺程度,清晨水势可以在一定程度上反映植物的水分恢复状况,而正午水势可以反映植物在当日所受最大环境胁迫的程度[27]。在唐钢梁等[28]的研究中,骆驼刺叶在重度遮阴下的水势最高,而在正常光照下水势最低。本研究也进一步证实了这个结论,4种植物均在L0时水势最低,随着光强的减弱,水势呈现不断上升的趋势,并在L4达到峰值。这表明强光会加剧植物体内的水分亏缺,而降低的水势可以提高植物的吸水能力,以弥补水分的亏缺[29]。此外,叶片相对含水量和失水速率是衡量植物保水力的重要指标,并有研究表明,植物拥有较低的失水速率是一种抗性生理机制,也就是具有较高的保水能力[30—31]。本研究中,4种植物的叶片相对含水量呈现增长趋势,这种响应很可能与弱光条件下蒸腾作用的减弱相关。一方面,光强减弱会使环境温度降低,相对湿度增加,进而降低了叶片温度,即减少了水分的蒸腾;另一方面,植物通过减弱蒸腾速率、增加体内水分含量可以缓解环境胁迫下产生的压力[32]。虽然随着光强的降低,不同种类植物的失水速率均有一定的升高,但是相比其他两种植物,CP与TZ的失水速率处于较低水平,表明CP与TZ的光适应性较强,这与前人的研究结果一致[33]。此外,正午水势与叶片的失水速率表现出较高的一致性,表明虽然遮阴在一定程度上能够提高相对含水量,但是同时也明显地降低了叶片的保水能力[34]。然而,从另一个角度说明,失水速率不同程度的增加,表明植物气孔的开放程度增加,进而促进气体交换及光合作用,最终促进物质积累[35]。
3.3 光强对光响应特征的影响
植物的光合作用是一种对环境因素较为敏感的生理过程,光强是影响光合作用的主要因素之一。虽然不同植物对光环境变化的响应特征不同,但是在逆境与胁迫条件下,植物都会向着利于光合作用的方向发展[36]。光响应曲线是评估植物光合特征的强有力工具,在本研究中,随着PAR的增加,4种植物的Pn均是先快速增长,而后进入平台期,这与前人的研究结果一致[37]。在L3与L4弱光条件下,CP、TZ具有较高的Pn,表明其弱光的利用能力较强。可以发现4种植物的Pm均呈现出先升后降的趋势,且CP、TZ与TV均在L2条件下达到最大值,尤其以TZ、TV的增长更为显著,表明强光会抑制4种植物的光合作用,适当的遮阴有利于提高植物的光合效率。通常情况下,AQY值越高,光能利用率就越高[38]。CP、TZ较其他两种植物具有较高的AQY,且涨幅也较大,表明这两种植物利用光能的可塑性较强,即耐阴性较强。除此之外,植物的光补偿点与光饱和点也是衡量植物耐阴性的重要指标,刘海燕等[39]发现具有较高LSP和较低LCP的植物,拥有更强的生态适应性。较低的Rd值能够反映出对光合产物的较低消耗,同时形成了较高的光合效率(Pm/Rd)[40]。本研究中,TZ的LCP值偏低,同时其与TV拥有较低的Rd,意味着TZ与TV能够减弱呼吸作用所造成的碳损耗,以此维持了碳代谢平衡。WUE是用来表征叶片尺度上的水分利用情况的关键性指标,能有效地反映植物体内的耗水机制[41]。研究结果显示,CP、TZ的WUE在不同处理间无显著差异,均表现出较高的水平,表明这两种植物拥有较高的气孔开闭调节能力以利于光合作用过程中气体和水分的交换,从而提高了光合及有效利用水分能力[42]。TF、TV分别在L2、L1条件下达到最大水分利用效率,表明适当的遮阴有利于二者水分利用效率的提高,这可能与高光强下的温度较高、空气湿度较小有关[43—44]。4种植物均在L4达到水分利用效率的最低值,说明极端的荫蔽环境不利于植物的光合作用。
4 结论
不同光强对4种鸭跖草科植物的生物量分配、水分生理及光响应特征有显著影响,结果显示4种鸭跖草科植物对光强均具有较好的适应性,能通过增加茎生长的方式提高地上生物量分配以获得更多的光资源,降低水势与保持较高的相对含水量去维持体内的水分平衡和正常的光合作用,提高表观量子效率与水分利用效率以提高光利用效率,其中紫鸭跖草和吊竹梅表现出了更佳的光适应性与较强的光利用效率。而全光照和极端的弱光环境会抑制植物的生长发育。因此在园林应用中,遮光度25%~75%的光环境更有利于4种植物达到最佳的生长状态,发挥最大的生态效益。
