基于6个玉米高代回交重组自交系群体的产量及其构成因素的杂种优势研究
2022-02-08张全艳丛一宁张培高徐春霞
张全艳,丛一宁,张培高,徐春霞,王 晶,刘 丽,3
(1. 云南大学农学院,昆明 650504;2. 云南省农业科学院粮食作物研究所,昆明 650205;3. 云南省农业科学院生物技术与种质资源研究所,昆明 650205)
【研究意义】杂种优势是生物界普遍存在的现象,其中玉米是最早利用杂种优势大幅度增加产量的作物之一[1],目前全球玉米杂交种应用面积占总种植面积的75%以上。合理划分杂种优势群,探索最优的杂种优势模式,是选育优良杂交玉米品种的前提。Jones等[2]根据亲缘关系从Reid黄马牙中划分出Lancaster和Reid两个群,构建了Lancaster × Reid杂种优势模式。Bouchez等[3]提出Non-Reid群,构建了Reid × Non-Reid杂种优势模式,现已成为美国玉米育种的主要杂种优势模式。欧洲则主要以Reid × EF及其衍生群为主要杂种优势模式,墨西哥和南美主要以Tuxpeno × ETO优势群为主[4]。20世纪90年代,王懿波等[5]提出将中国的玉米种质划分为5大类群及9个亚群,然而杂种优势划分类群太多,工作复杂且较难应用于玉米育种。因此,Fan等[6]提出将玉米自交系划分为3个杂种优势群,即Suwan1群、Reid群和非Reid群,构建了三角形杂种优势模式。目前中国温带玉米主产区主要以Reid、Lancaster、塘四平头、旅大红骨和P群5个杂种优势群相互杂交为主[5],而在南方地区以Suwan1、Reid和非Reid等热带×温带为主[7]。国内外对玉米杂种优势进行了大量研究,但主要集中于杂种优势群的划分,即基于不完全双列杂交(NCII)设计组配杂交组合F1,通过分析其产量及产量性状的一般配合力(General combining ability, GCA)和特殊配合力(Specific combining ability, SCA)划分杂种优势群,进而育种利用[8-10]。【前人研究进展】玉米杂种优势形成的遗传基础研究尚处于探索阶段。Garcia等[11]分析前人的数据,发现玉米杂种优势主要受显性基因影响。Tang等[12]利用永久F2群体(Immortalized F2, IF2)研究产量及其构成因素的杂种优势,发现显性和超显性间的互作对玉米产量及构成因素杂种优势的形成有重要作用。宋方威等[13]利用高代回交重组自交系BC1F5∶6群体解析温带玉米杂种优势形成的遗传基础,定位了一批株高和穗位高相关杂种优势QTL。Yi等[14]用重组自交系群体(Recombinant inbred lines, RIL)分析9个产量性状及其中亲优势(Mid-parent heterosis)的遗传差异,结果表明穗粒重、穗重和行粒数等性状及中亲优势均为显性效应和超显性效应。目前永久F2群体和三重测交(Triple test cross, TTC)群体(含高代回交群体)是玉米杂种优势形成机理研究的主要群体类型。【本研究切入点】现有的研究材料大多为温带玉米自交系,而对热带和亚热带玉米杂种优势形成的研究报道较少,并且高代回交重组自交系群体主要用于株高和穗位高等株型性状的杂种优势研究,目前未见利用该类群体分析产量及其构成因素杂种优势形成机理的报道。此外,国外杂种优势群主要分为2个类群,国内虽分为多个类群,但由于研究材料主要为F1杂交组合,未能进行不同杂种优势模式间产量杂种优势强弱的比较,因此不同杂种优势模式间产量及其构成因素的差异性比较尚未见报道。【拟解决的关键问题】本研究选用三角形杂种优势代表自交系Y46、Y107和MON2,组配6个BC1F7∶8群体,采用穗长、穗粗、穗行数、行粒数和单产5个产量及其构成因素的中亲优势和超亲优势(Over-parent heterosis),比较不同杂种优势模式间的产量杂种优势强弱,为中国温热玉米种质互导、种质创新和杂种优势利用等提供参考。
1 材料与方法
1.1 试验材料
Y46:热带玉米自交系,选自Suwan1种质。Suwan1源自泰国,其遗传基础较广泛、配合力高,抗倒伏、抗病性、抗旱能力强,商品品质好,是中国玉米种质扩增的核心材料。
Y107:中国温带地区重要骨干亲本,是20世纪90年代中国推广面积最大的杂交种掖单2号的亲本,其千粒重高、种胚大,产量潜力大。
MON2:选自美国自交系,为中国亚热带地区的重要骨干亲本。
根据Zhang等[15]和宋方威等[13]的高代回交群体构建方法,将上述优良自交系相互杂交组配3个F1杂交群体,继而连续自交形成3个RIL-F7∶8群体。分别从Y46与Y107组配的RIL-F7∶8群体中选择161个和91个RIL系与双亲回交组配得到2个BC1F7∶8群体Y46×Y107/Y46(正交)和Y46×Y107/Y107(反交)。分别从Y46与MON2组配的RIL-F7∶8群体中选择135个和140个RIL系与双亲回交组配得到2个BC1F7∶8群体Y46×MON2/Y46(正交)和Y46×MON2/MON2(反交)。分别从Y107与MON2组配的RIL-F7∶8群体中选择218个和104个RIL系与双亲回交组配得到2个BC1F7∶8群体Y107×MON2/Y107(正交)和Y107×MON2/MON2(反交)。
1.2 试验方法
将6个BC1F7∶8群体分别种植于云南不同海拔高度的试点昆明点(25°02′N,120°45′E;海拔1960 m),砚山点(23°19′N,104°45′E;海拔1520 m)。本试验采用随机区组设计以有效地减少土壤肥力差异对试验的影响、降低产量性状的误差,设2次重复。每个小区长2.5 m,株距25 cm,密集种植10株,施肥、除草、除虫按田间标准管理方法进行。玉米成熟后,从每行的第3株开始连续收获5穗,晾晒至含水量低于14%时进行考种。调查穗长(Ear length, EL)、穗粗(Ear diameter, ED)、穗行数(Ear row number, ERN)、行粒数(Kernel number per row, KNPR)和单产(Yield per unit area, YPUA)5个产量及其构成因素。
1.3 数据统计与分析
利用SPSS软件对产量及其构成因素进行基本统计量和方差分析,如果试点间和重复间无显著差异,则杂种优势分析用试点和重复的平均值进行分析;如果存在显著差异,则比较试点间和重复间的趋势是否一致。通过计算各个BC1F7∶8群体的产量及构成因素的中亲优势和超亲优势比较杂种优势强弱,计算方法[16]如下。
中亲优势(%)=(BC1F7∶8-MP)/MP×100
超亲优势(%)=(BC1F7∶8-HP)/HP×100
式中,BC1F7∶8、MP和HP分别代表高代回交重组自交系群体、2个亲本和高值亲本各产量及其构成因素的平均值。
2 结果与分析
2.1 高代回交重组自交系群体产量及其构成因素的差异性比较
在2个不同环境条件下,对Y46、Y107和MON2所组配的6个BC1F7∶8群体的穗长、穗粗、穗行数、行粒数和单产5个产量及其构成因素进行比较,结果如表1所示。昆明点的产量及其构成因素均高于砚山点,变异系数则表现为砚山点高于昆明点,表明昆明点的产量潜力和稳产性均优于砚山点,说明环境对玉米产量及其构成因素有显著影响。在昆明点,Y46×Y107/Y46群体的穗长、穗粗、穗行数、行粒数和单产平均值分别为15.20 cm、3.80 cm、9.90行、29.90粒和8884 kg/hm2,除穗粗和穗行数以外,其它产量构成因素均高于Y46×Y107/Y107群体的平均值,表明正交群体的产量及其构成因素高于反交群体,砚山点分析结果与该结果趋势相反。Y46×MON2/Y46群体的穗长、穗粗、穗行数、行粒数和单产平均值分别为14.50 cm、4.20 cm、12.90行、27.50粒和8321 kg/hm2,明显高于反交群体,砚山点分析结果与该结果基本相同。Y107×MON2/Y107群体的穗长、穗粗、穗行数、行粒数和单产平均值分别为13.10 cm、3.50 cm、10.10行、23.90粒和7160 kg/hm2,除穗长外其它4个产量构成因素均低于Y107×MON2/MON2群体的平均值,表明上述因素在该杂交组配中反交群体大于正交群体,而砚山点与该结果则趋势相反。综上所述,3个正反交群体间产量及其构成因素均存在显著差异,说明在玉米杂交组配中,应同时进行正反交组配,选择高产的正交或反交群体应用于生产实践。进一步研究发现2个试点6个BC1F7∶8群体除单产外,穗长、穗粗、穗行数和行粒数的峰度和偏度绝对值均小于1,符合正态分布,说明BC1F7∶8群体间各产量构成因素以加性效应为主,其表型可用于后续的QTL定位研究。
2.2 高代回交重组自交系群体产量及其构成因素的方差分析
2.2.1 不同试点间产量及其构成因素的方差分析 从表2可以看出,6个BC1F7∶8群体产量及其构成因素在不同试点间差异达显著水平,其中穗粗、穗行数、行粒数和单产在两试点间差异均达极显著水平;穗长除了Y107×MON2/MON2群体在两试点间差异不显著外,其它群体在两试点间达显著水平。说明不同试点对BC1F7∶8群体的产量及其构成因素有显著影响。

表1 高代回交重组自交系群体产量及其构成因素的基本统计量
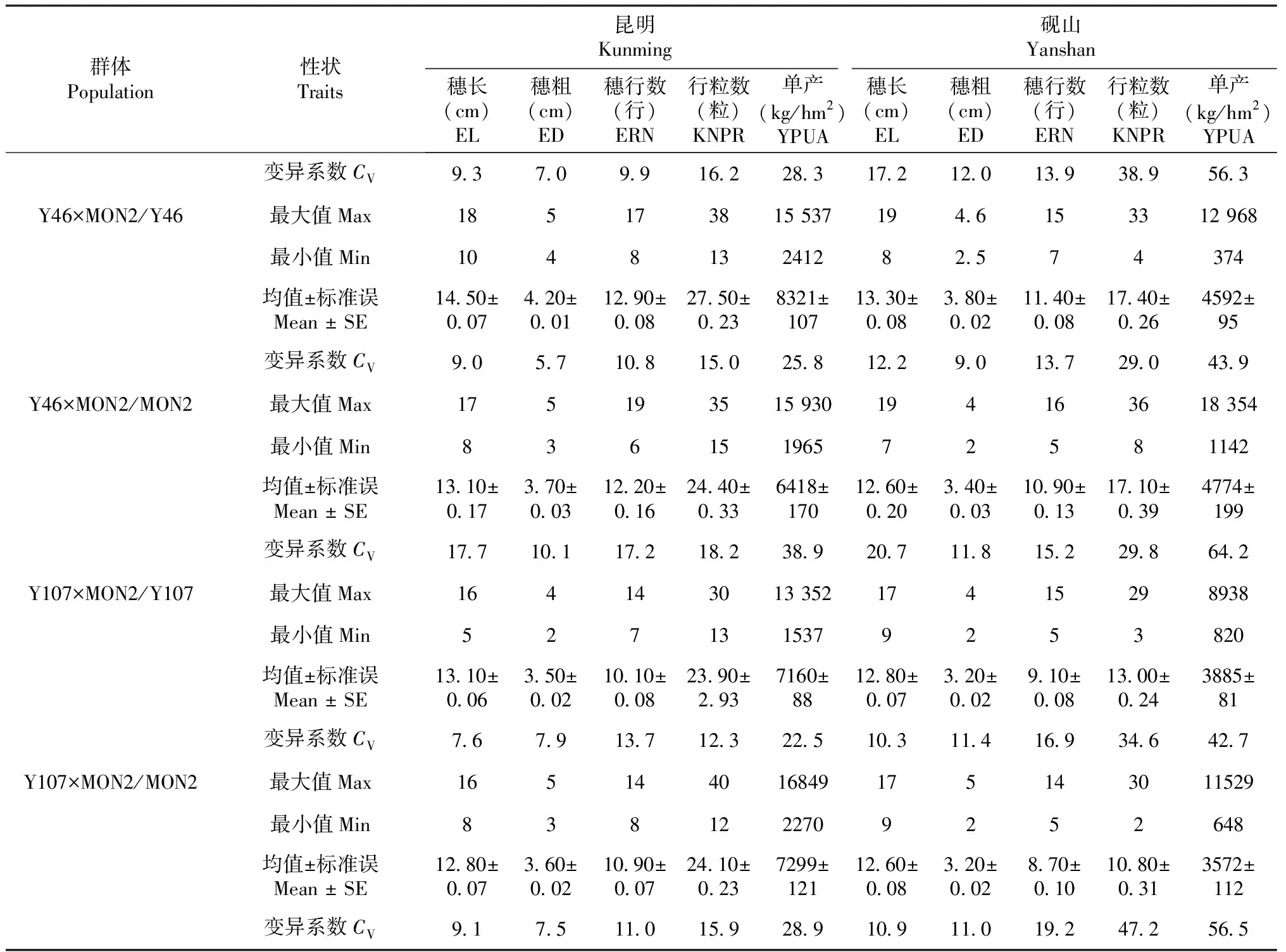
续表1 Continued table 1
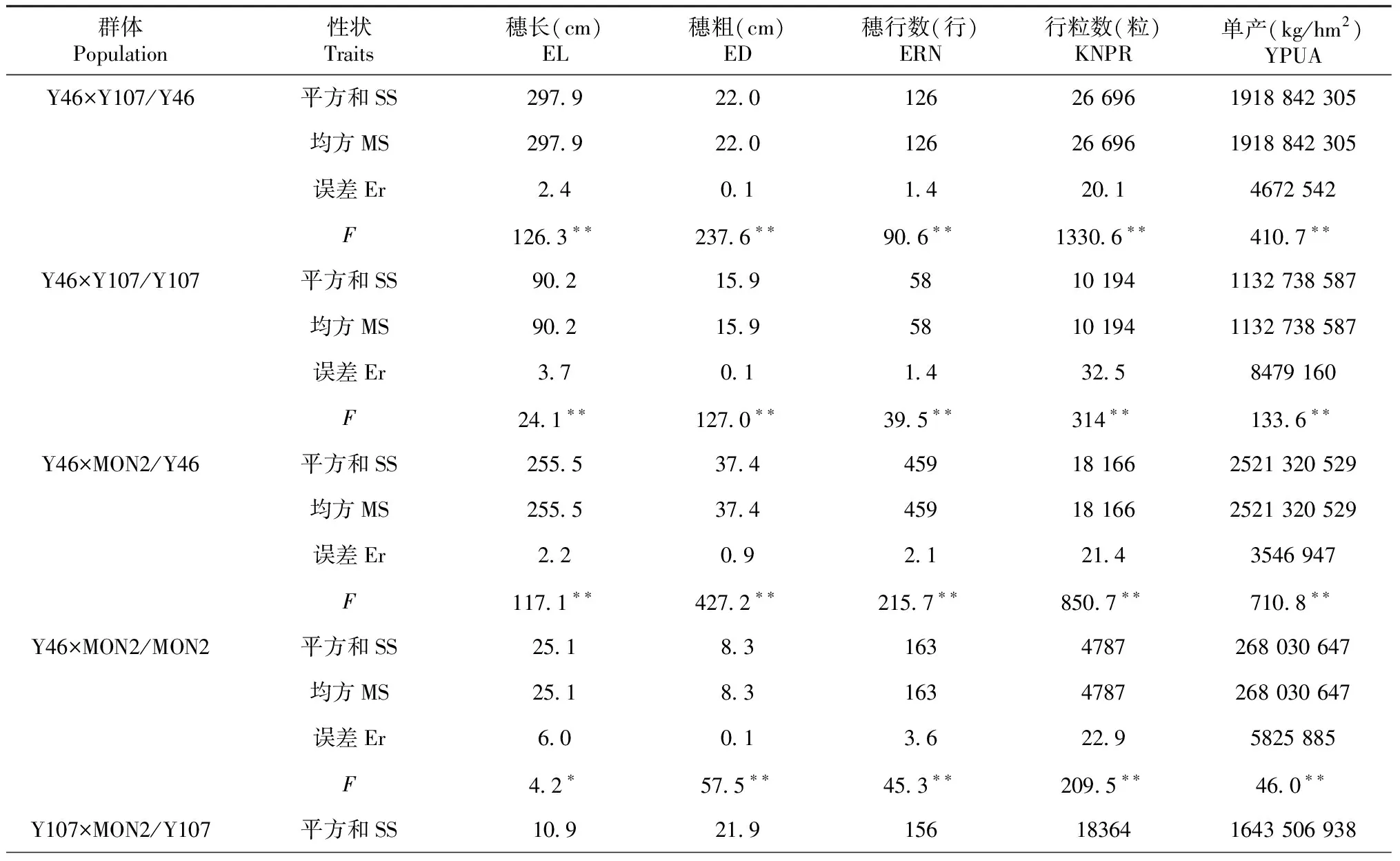
表2 不同试点间产量及其构成因素的方差分析
续表2 Continued table 2
2.2.2 重复间产量及其构成因素的差异显著性比较 差异显著性比较结果表明,在Y46×MON2/Y46群体不同重复间除行粒数(P=0.200>0.05)外,其它产量及产量构成因素(P=0.001~0.002<0.01)均达极显著水平;在Y46×Y107/Y107和Y46×MON2/MON2群体的不同重复间穗长(P=0.514~0.610>0.05)、穗粗(P=0.143~0.390>0.05)、穗行数(P=0.392~0.737>0.05)、行粒数(P=0.504~0.511>0.05)和单产(P=0.108~0.436>0.05)差异均不显著;在Y46×Y107/Y46群体不同重复间除穗长(P=0.001<0.01)差异达极显著水平外,其它产量构成因素差异均不显著。在Y107×MON2/MON2群体不同重复间除行粒数(0.05
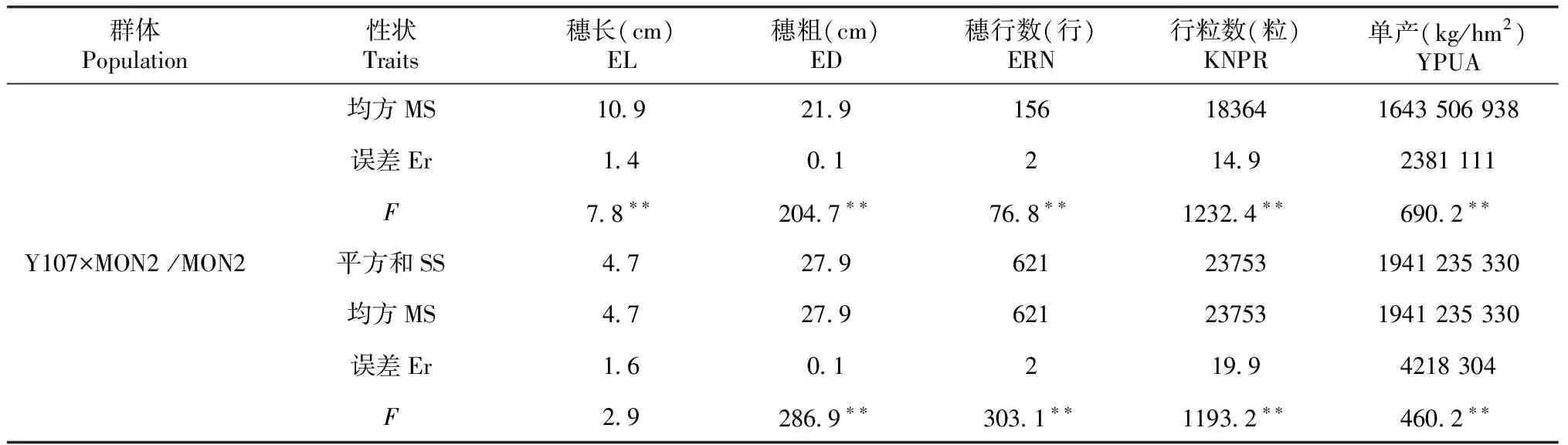
2.2.3 高代回交重组自交系群体内不同材料间产量及其构成因素的方差分析 如表3所示,6个BC1F7∶8群体内不同材料间产量及其构成因素差异基本达显著。在Y46×Y107/Y107和Y46×MON2/MON2群体重组自交系内不同材料间穗长、穗粗、穗行数、行粒数和单产的差异均达极显著水平。在Y46×Y107/Y46和Y46×MON2/Y46群体重组自交系内不同材料间穗长、穗粗和穗行数差异均达极显著水平;其中,在Y46×Y107/Y46群体重组自交系内不同材料间行粒数差异未达显著水平,单产差异达极显著水平;而在Y46×MON2/Y46群体重组自交系内不同材料间行粒数差异达显著水平,单产差异不显著。在Y107×MON2/Y107群体重组自交系内不同材料间仅有穗粗和穗行数差异达极显著水平,其它产量构成因素差异不显著。而在Y107×MON2/MON2群体重组自交系内不同材料间穗长和穗粗差异达极显著水平,单产差异达显著水平,穗行数和行粒数差异不显著。表明这些家系遗传背景差异较大或亲缘关系较远。
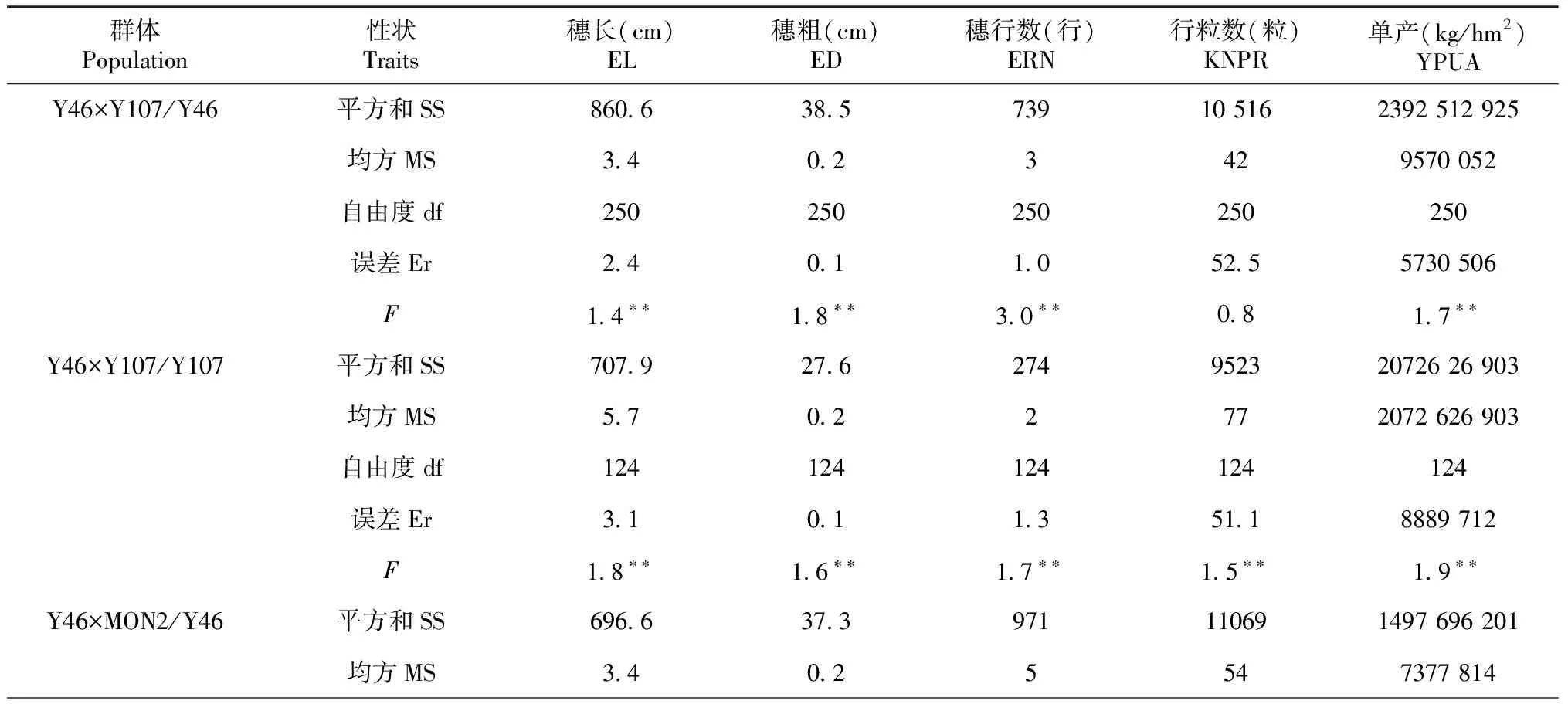
表3 高代回交重组自交系群体内不同材料间产量及其构成因素的方差分析

续表3 Continued table 3
综上所述,在不同试点中,BC1F7∶8群体的产量及其构成因素差异达显著水平,而重复间差异不显著,群体内重组自交系内不同材料间产量及其构成因素差异显著,说明玉米产量及其构成因素主要由基因型决定,但同时受环境的影响。
2.3 高代回交重组自交系群体产量及其构成因素的主成分分析
利用线性回归模型分析6个BC1F7∶8群体各产量构成因素与产量的相关性。结果表明,穗长、穗粗、穗行数和行粒数与产量的相关系数分别为0.63、0.83、0.76和0.83,均达显著水平。从表4可知,2个主成分可以反映BC1F7∶8群体各产量构成因素的大部分信息,Y46×Y107/Y46群体和Y46×Y107/Y107在砚山点中主成分累计方差贡献率均低于80%,分别为76.68%和79.97%。其它群体的主成分累计方差贡献率均高于80%,Y46×Y107/Y46群体在昆明点主成分累计方差贡献率最高,为84.97%,Y107×MON2 /MON2群体在砚山点主成分累计方差贡献率最高,为84.06%。就产量构成因素而言,穗粗的第一主成分和第二主成分特征向量值最高,分别为0.65~0.92和-0.37~0.65,穗长最低,分别为0.18~0.80和-0.55~0.94。因此,在杂交玉米高产育种中,建议将穗粗作为新品种选育的主要改良目标。同时,兼顾行粒数对产量的影响。

表4 高代回交重组自交系群体产量及其构成因素的主成分分析
2.4 高代回交重组自交系群体产量及其构成因素的杂种优势分析
由表5可以看出,昆明点Y46与Y107组配的群体行粒数的中亲优势和超亲优势、穗长和单产的中亲优势以及反交群体穗长的超亲优势都表现为正向杂种优势效应。两试点的结果基本一致。Y46与MON2组配的群体在两试点间产量及其构成因素的杂种优势都为负向杂种优势效应。昆明点Y107与MON2组配的群体穗长、行粒数和单产的中亲优势以及其正交群体穗长的超亲优势都呈正向杂种优势效应,砚山点均表现为负向杂种优势效应。
在Y46×Y107/Y46群体中,其穗长、穗粗、穗行数、行粒数和单产的中亲优势分别为29.8、-7.3、-17.1、57.5和12.0,超亲优势分别为-5.1、-13.6、-28.9、15.1和-25.8,低于反交群体穗长(30.0,0.6)、穗粗(-4.8,-9.1)、穗行数(-14.9,-24.0)和单产(16.7,-15.3)的中亲优势和超亲优势,其中反交群体行粒数的中亲优势(52.0)低于正交群体,超亲优势(22.2)高于正交群体,由此可见反交群体杂种优势强于正交群体,且两试点的群体杂种优势分析结果基本一致。在Y46×MON2/Y46群体中,其穗长、穗粗、穗行数、行粒数和单产的中亲优势分别为-10.4、-6.2、-7.7、-1.8和-32.3,超亲优势分别为-10.9、-8.3、-13.9、-11.3和-33.7,显著高于反交群体穗长(-18.5,-20.7)、穗粗(-18.2,-23.3)、穗行数(-9.3,-18.4)、行粒数(-14.3,-15.7)和单产(-50.2,-55.6)的中亲优势和超亲优势,说明在Y46与MON2组配的群体中正交群体杂种优势比反交群体强,并且砚山点与昆明点趋势一致。在Y107×MON2/Y107群体中,其穗长、穗粗、穗行数、行粒数和单产的中亲优势分别为12.4、-9.2、-6.1、2.8和2.0,超亲优势分别为5.6、-11.5、-12.2、-0.5和-14.2,反交群体穗粗(-6.0,-9.5)、穗行数(-1.4,-9.6)的中亲优势和超亲优势大于正交群体,而穗长(10.4,-7.2)和单产(0.2,-26.6)的中亲优势和超亲优势均小于正交群体,行粒数(18.9,-12.3)的中亲优势大于正交群体,超亲优势小于正交群体,说明Y107与MON2组配的反交群体杂种优势强于正交群体,而砚山点杂种优势则表现为正交群体大于反交群体。
将Y46与Y107、Y46与MON2和Y107与MON2组配的6个BC1F7∶8群体的产量及其构成因素进行综合比较,结果显示Y46×Y107/Y107群体穗长、穗粗、穗行数、行粒数和单产的中亲优势和超亲优势均大于其它群体。在Y46×Y107/Y46与Y107×MON2/MON2群体中,Y46×Y107/Y46群体穗长(29.8,-5.1)、行粒数(57.5,15.1)和单产(12.0,-25.8)的中亲优势和超亲优势均大于Y107×MON2/MON2群体。Y107×MON2/Y107群体穗长(12.4,5.6)、穗行数(-6.1,-12.2)、行粒数(2.8,-0.5)和单产(2.0,-14.2)的中亲优势和超亲优势均大于Y46×MON2/Y46群体,砚山点与昆明点基本相同。总体而言,6个BC1F7∶8群体及其构成因素的杂种优势强弱排序为Y46×Y107/Y107>Y46×Y107/Y46>Y107×MON2/MON2>Y107×MON2/Y107>Y46×MON2/Y46>Y46×MON2/MON2,说明在三角形杂种优势模式中,Suwan1×Reid杂种优势模式组配的群体,其后代产量及其构成因素的中亲优势和超亲优势较强,以Reid×非Reid杂种优势模式组配的群体,其后代产量及其构成因素的中亲优势和超亲优势居中,以Suwan1×非Reid杂种优势模式组配的群体,其后代产量及其构成因素的中亲优势和超亲优势较弱。
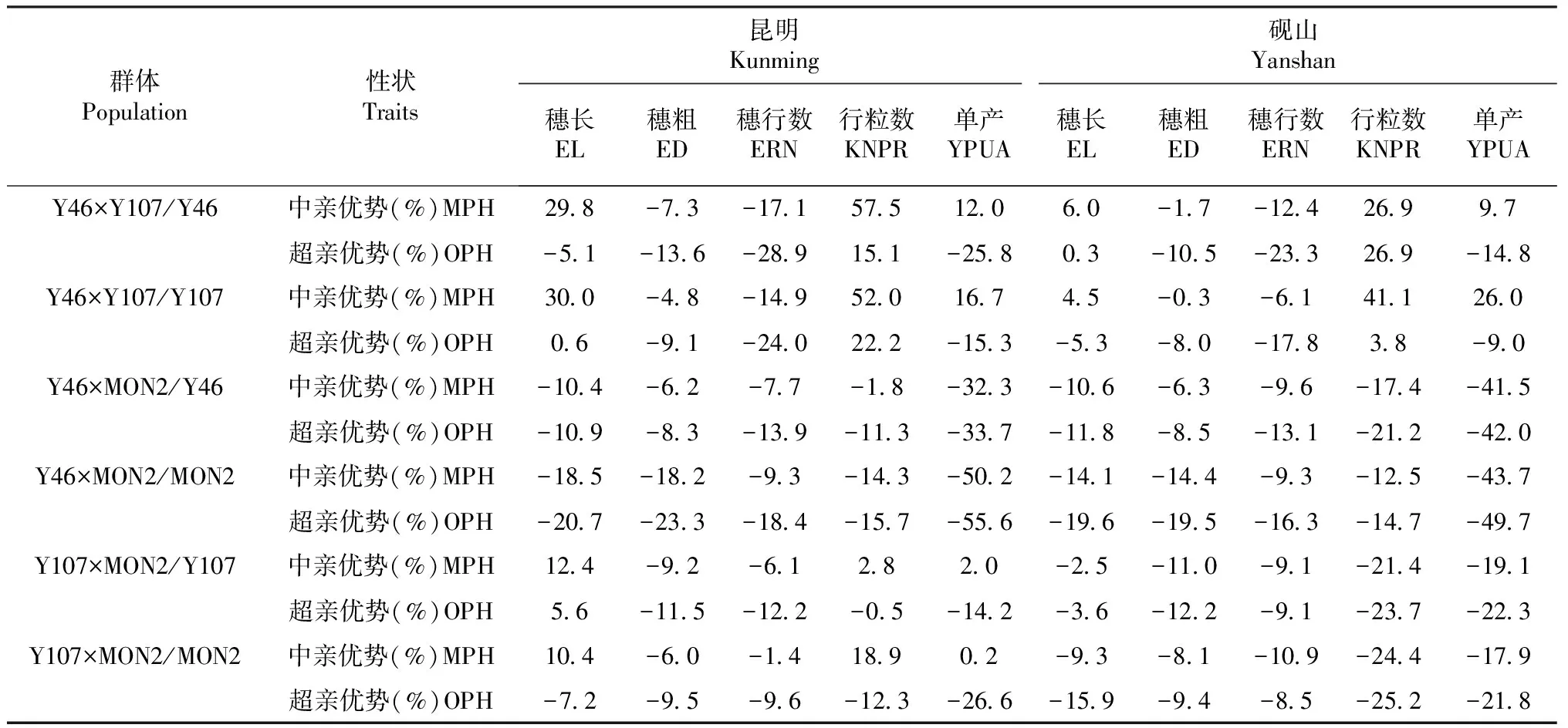
表5 高代回交重组自交系群体产量及其构成因素的杂种优势分析
3 讨 论
玉米产量性状是由微效多基因所控制的数量性状,遗传力较低,其穗长、穗粗、穗行数和行粒数等性状均是构成产量的重要因素,剖析产量构成因素的遗传规律对玉米高产育种具有重要的指导意义。宗宪春等[17]研究表明,不同生产方式导致行粒数和穗粒重显著下降,严重影响玉米的产量。Tan等[18]研究结果表明,日温差、日照时长和有效积温与穗粗、行粒数、百粒重呈正相关,导致产量大幅度上升。多个研究均表明千粒重、行粒数和穗长等对玉米产量具有显著的影响[9,19]。本研究中穗长、穗粗、穗行数和行粒数与产量间显著相关,相关系数均大于0.63,通过主成分分析发现,穗粗和行粒数对玉米产量有重要决定作用,进一步验证了前人的研究结果[9,17-19]。
正反交是2个基因型不同的亲本互作母本或父本的杂交群体,而亲本互换与后代表达情况密切相关。冯汉宇等[20]通过正反交试验研究花青素基因在玉米杂种性状中的表达情况,发现转基因植株做父本对转基因杂交育种有重要影响。Liu等[21]发现,梨的选育过程中酸含量明显取决于母本的选择。Dermail等[22]发现,正反交影响着杂种优势效应,可以提高玉米产量。这些研究表明,亲本材料的选择对杂交后代性状的影响程度不同,各性状间也将存在一定差异,要想得到优良的目标性状,亲本选配至关重要。此外,本研究利用6个BC1F7∶8群体分析正反交对穗长、穗粗、穗行数等产量及其构成因素的影响,也发现正反交群体间产量及其构成因素差异显著,特别是在Y46与MON2组配的群体中,正反交差异较大。说明在Suwan1×非Reid杂种优势模式中,要特别注意母本的选择,选择适宜的正交或反交,以最大限度发挥产量潜力优势。本研究比较不同重复间的差异显著性时,发现Y46×MON2/Y46群体不同重复间除行粒数外,其它产量及产量构成因素均达显著或极显著水平,可能原因是该群体的产量稳定性较差,受环境的影响较大,但尚需进一步验证。
研究表明,GCA效应较高的自交系更容易获得高产杂交种,组合间配合力总效应越高,杂种优势越强[23]。此外,一般配合力由基因的加性效应决定,可逐代遗传,特殊配合力由基因的非加性效应所控制,易受外部环境条件的影响,不能在上下代间稳定遗传。杂种优势群的配合力主要以提高群内的GCA为主,同时还应保持或提高群间的SCA,只有GCA和SCA效应都高时,才能组配出强杂种优势组合。前人[23]的研究只能对杂种优势进行划分,不能将不同杂种优势模式间形成的杂种优势进行比较。本研究利用6个BC1F7∶8的中亲优势和超亲优势来分析Suwan1、Reid和非Reid 3个类群间产量及其构成因素的杂种优势强弱,明确了Suwan1×Reid杂种优势模式中产量及其构成因素的杂种优势最强,Reid×非Reid次之,Suwan1×非Reid较弱。该研究与前人[24]利用产量平均值初步推测的结果基本一致。
杂种优势的利用是提高玉米产量的重要途径[25]。基于前期使用热带、亚热带与温带玉米相互杂交,构建三角形杂种优势模式,即Suwan1×Reid、Suwan1×非Reid、Reid×非Reid[12]。利用基于NCII设计组配的F1杂交组合,Fan等[15]进一步研究发现三群(Suwan1、Reid和非Reid)比两群(Reid和非Reid)育种效率更高,可提高77.8%的一般育种效率(General Breeding Efficiency, GBE)。本研究利用BC1F7∶8群体比较其中亲优势和超亲优势,发现Suwan1×Reid杂种优势明显强于Reid×非Reid、Suwan1×非Reid杂种模式,可见Suwan1×Reid后代中产量及其构成因素不仅保留了Suwan1种质配合力高、商品品质好等特点的同时,还导入Reid种质遗传效应稳定、果穗较好的优良基因,因此Suwan1×Reid后代的产量及其构成因素杂种优势表现较强。而Reid×非Reid杂种模式也有部分产量及其构成因素表现出正向杂种优势,尤其是穗长和行粒数,今后在创新种质资源或改良目标性状时多加关注。Suwan1×非Reid杂种模式的产量及其构成因素均表现出负向杂种优势,可能是随着育种进程长期化,热带和亚热带种质间存在广泛的基因交流,造成了Suwan1与非Reid种质遗传差异较小,进而导致杂种优势表现程度较弱[24,26]。
在玉米中,一般利用F1和RIL等群体对产量性状进行杂种优势分析,而利用BC1F7∶8群体进行产量及其构成因素杂种优势分析的研究报道较少。与常规群体相比,由于BC1F7∶8群体经过多次回交,重组自交系间遗传差异相对较小,因此,更容易精确地定位QTL的位置,并且对主效、微效和加性效应的QTL检测更高效[27]。
4 结 论
Suwan1×Reid杂种优势模式的产量及其构成因素杂种优势较强,Reid×非Reid次之,Suwan1×非Reid较弱。
