基于涡度协方差法和植被指数的油菜光能利用率研究
2022-02-07文双雅石楠高倩文阳会兵高志强卢俊玮
文双雅,石楠,高倩文,阳会兵,高志强,卢俊玮
(湖南农业大学农学院,长沙 410128)
光能利用率(Light use efficiency,LUE)是光合作用关键参数,表示植被通过光合作用将所吸收能量转化为有机干物质的能力[1],是区域尺度以遥感参数模型监测植被生产力的关键参数[2]。因植被分布及气候环境综合影响,光能利用率表现显著空间异质性和时间动态性[3],光能利用率的不确定性导致基于LUE遥感模型估算结果存在一定偏差,因此了解LUE生理生态基础对LUE模型优化和对模型模拟结果可靠性评价具有重要意义[4]。早期光能利用率计算方法主要通过持续或间隔一定时间对植物生长季地上生物量进行全部收割,再与冠层截获并吸收的光合有效辐射相比得出光能利用率[5]。该方法所需仪器简单,但耗时费力,破坏植被,无法提供短时间尺度上LUE变化信息[6]。近年来,涡度协方差法成为目前以高时间分辨率直接测算生态系统碳通量的可行方法[7],为研究LUE时空变化特征提供重要数据支撑。
前人研究表明,目前结合农田通量观测数据开展作物光能利用率研究主要集中在水稻、冬小麦、玉米、大豆等作物。Chen等研究全球11个农田通量站点发现,夏玉米生长季生态水平光能利用率最高,其次为大豆、水稻和冬小麦[8];文双雅等利用通量观测数据计算的水稻生长季平均生态水平光能利用率为0.95%[9];费敦悦等研究安徽寿县稻田生态系统发现,其光能利用率波动范围为0.4%~4.6%[10]。油菜作为中国乃至世界最重要油料作物之一[11],以往研究多集中在不同光合器官及高光效育种方面[12],而基于涡度协方差法开展油菜光能利用率动态变化研究鲜有报道。
本研究基于涡度相关通量数据分析油菜光能利用率时序特征,探究日尺度上光能利用率对外界条件响应,以期为后续优化模型参数、推进模型发展,因地制宜应用光能利用率模型进行生态系统生产力估测提供参考。
1 材料与方法
1.1 研究站点
研究区位于湖南农业大学浏阳沿溪基地(113°84'E,28°30'N),属亚热带季风湿润气候,年平均气温17.3℃,1月平均气温5.4℃,7月平均气温28.7℃,年均降水量1 358.6~1 552.5 mm。研究区以涡度相关通量塔为中心,观测区面积约1.33 hm2,地势平坦,历年种植制度为水稻/油菜一年两熟制。土壤为潴育性水稻土,有机质、全氮、全磷、全钾含量分别为23.41、1.73、0.64、19.35 g·kg-1,pH为5.71。油菜品种为湖南农业大学油料作物改良中心育成的湘杂油787,2020年9月30日采用直播方式播种,2021年5月4日收获。土壤耕作采用铧式犁翻耕后,碎土整畦成厢。播后、苗前封闭除草。栽培措施和田间管理按当地常规实施(见图1)。

图1 研究区数据观测条件Fig.1 Data observation conditions in the study area
1.2 数据观测
研究区通量观测采用开路式涡度相关系统,主要包括开路式CO2/H2O分析仪(LI-7500,Li-cor,USA)、三 维 超 声 风 速 仪(CSAT3,Campbell,USA)、数据采集器(CR1000,Campbell,USA、实时在线通量计算模块(Smart Flux,Li-cor,USA)及供电设备。数据采样频率为10 Hz,每30 min输出一组平均通量及10 Hz原始数据。采用归一化植被指数测量仪(SRS-NDVI,METER,USA)测量冠层尺度归一化植被指数(Normalized difference vegetation idex,NDVI)。仪器安装在油菜冠层上方,包含向上和向下两个传感器,可同时测量冠层对近红外波长(810 nm)反射率与红光波长(650 nm)反射率,SRS-NDVI观测仪时间分辨率为30 min,数据自动存储在数据采集器内(EM50,METER,USA)。研究区内配备小型气象站观测油菜生长季内环境因子变化,包括空气温度(Air temperature,Ta)、土壤温度(Soil temperature,Ts)、光合有效辐射(Photosynthetic active radiation,PAR)、饱和水汽压差(Vapor pressure deficit,VPD)等,观测频率为10 min,监测仪器每隔30 min计算1组均值,通过内置数据采集器自动存储。
1.3 数据处理
1.3.1 通量数据处理
使用Eddypro通量数据处理软件(Li-cor,USA)处理计算原始数据,使用Tovi通量数据分析软件(Li-cor,USA)对数据进行质量控制与插补[13]。总初级生产力(Gross primary productivity,GPP)是在单位时间和单位面积上,绿色植物通过光合作用所固定有机碳总量。生态系统总呼吸(Ecosystem respiration,Reco)是生态系统中植物和异养生物呼吸总和,由于涡度相关系统仅获得生态系统净碳交换量NEE(Net ecosystem exchange,NEE),冠层尺度GPP和Reco难以直接观测,为后续计算光能利用率,需要通过涡度观测系统观测并对经后续处理所得到NEE作拆分,该步骤在Tovi软件中进行:

1.3.2 植被指数数据处理
NDVI反映作物长势,利用冠层对近红外波长(810 nm)反射率与红光波长(650 nm)反射率之差与两者之和比值可计算得到NDVI:

1.3.3 生态水平上光能利用率计算
生态光能利用率不仅反映整个生态系统植被对光能利用效率,也反映植被结构特征对生态系统光能利用率影响,本研究主要分析LUEeco变异性及其影响因素。使用通量观测计算获得总初级生产力(GPP)与光合有效辐射(PAR)比值可得到生态水平上光能利用率(Ecosystem light use efficiency,LUEeco):

1.3.4 表观初始量子效率计算
由于生态系统呼吸与光合作用之间无明确定量关系,使NEE与PAR之间关系弱化,因此,理论上GPP可更好表征生态系统冠层光合效率与光合能力[14],GPP与PAR关系可用下式描述[15]:

其中,PAR为光合有效辐射,Pmax为最大光合速率,Km为拟合常数,由以上参数可计算生态系统表观量子效率α:

1.4 数据分析
使用Microsoft Excel 2013整理数据,SPSS 26.0软件作简单相关分析、多元线性逐步回归分析和通径分析,采用Origin 2021软件绘制图表。
2 结果与分析
2.1 LUEeco变化特征
9月30日为油菜播种日,本研究仅考虑油菜生育期10月到次年5月光能利用率变化,油菜生长季内不同月份LUEeco日变化如图2所示。日尺度上LUEeco表现为3段式变化:9:00前,由于光合有效辐射逐渐升高且增长迅速,这一时段内PAR增长显著大于GPP增加,导致LUEeco不断下降;9:00~15:00,光合有效辐射达到较高水平,油菜植株由于受到较高光合有效辐射影响,出现“午休效应”,此时PAR和GPP波动均较小,LUEeco维持在较低水平;15:00后,PAR迅速降低,LUEeco升高。不同月份LUEeco日变化差异主要体现在曲线高低上。
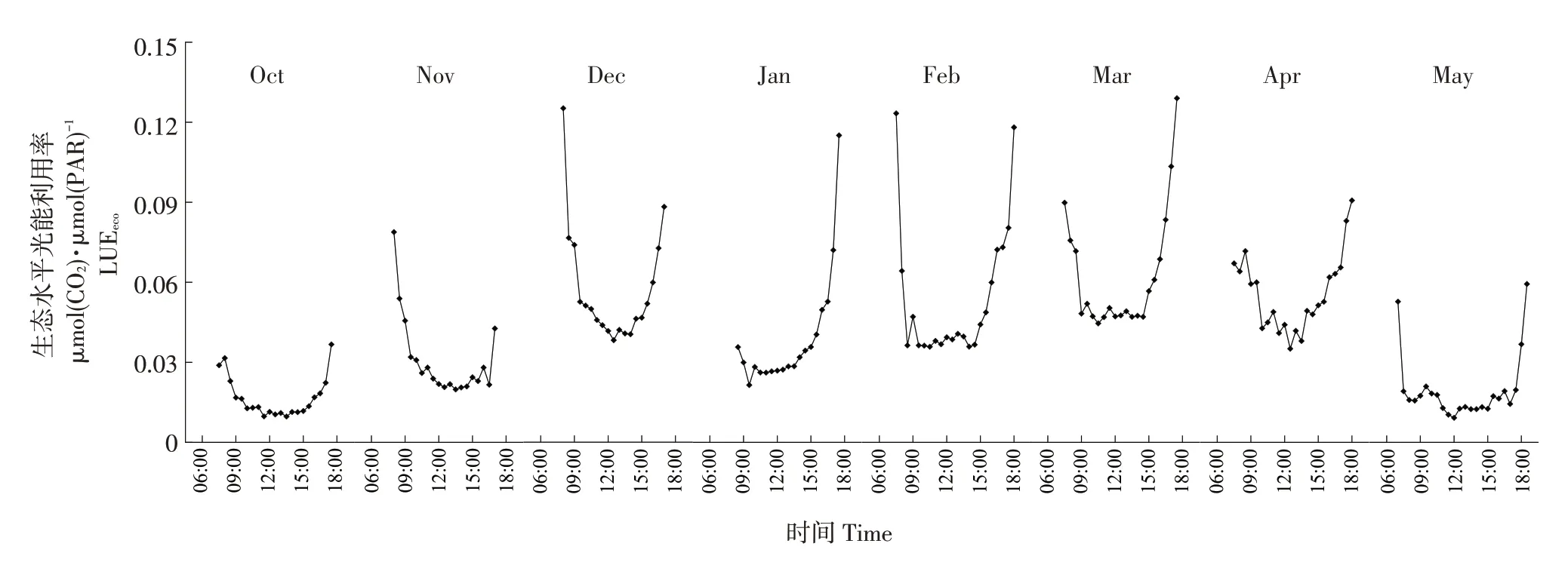
图2 不同月份LUEeco小时尺度变化Fig.2 Variation of LUEeco hourly scales at different months
油菜LUEeco月变化如图3所示,10月到次年5月LUEeco表现为先升后降趋势,与油菜生长发育状况具有一致性。10月油菜播种后处于苗期,植株矮小,冠层结构疏松,叶片和冠层利用光能能力均较弱,因此LUEeco表现为较低水平,随着植株变高、冠层变密,油菜光合能力不断增加,3月,油菜处于花期时LUEeco达到最高值,为0.0633 μmol(CO2)·μmol(PAR)-1,之后随着叶片衰老,到5月(角果期),LUEeco下降到较低水平。
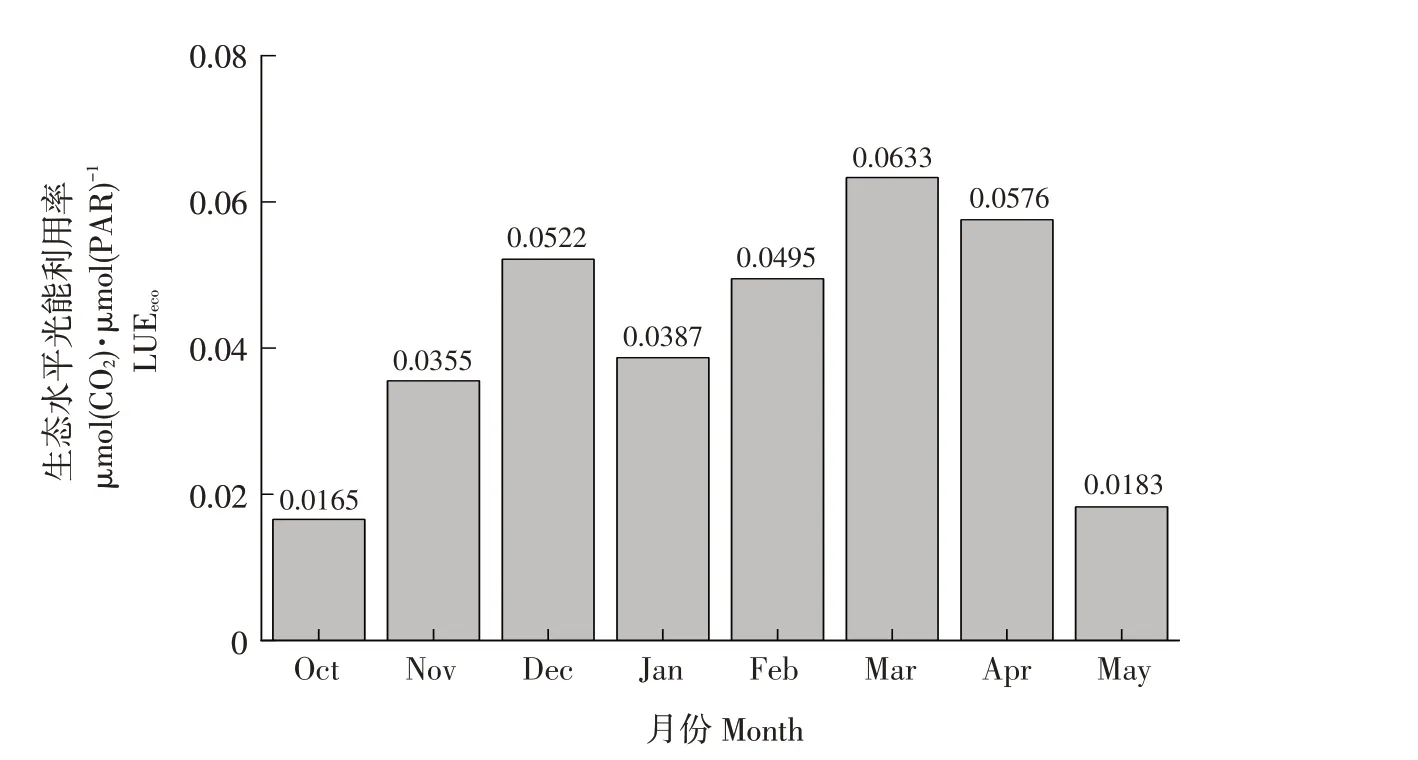
图3 LUEeco月变化Fig.3 LUEeco monthly change
图4为油菜生长季逐日LUEeco变化动态,从整体上看,LUEeco在油菜苗期时表现为较低水平,随着生育期推进,LUEeco表现为波动上升,且后期波动幅度相较于前期更大,可能因外界环境条件而引起变化。LUEeco范围在0.0011~0.1216 μmol(CO2)·μmol(PAR)-1,油菜全生长季平均LUEeco为0.0347 μmol(CO2)·μmol(PAR)-1。
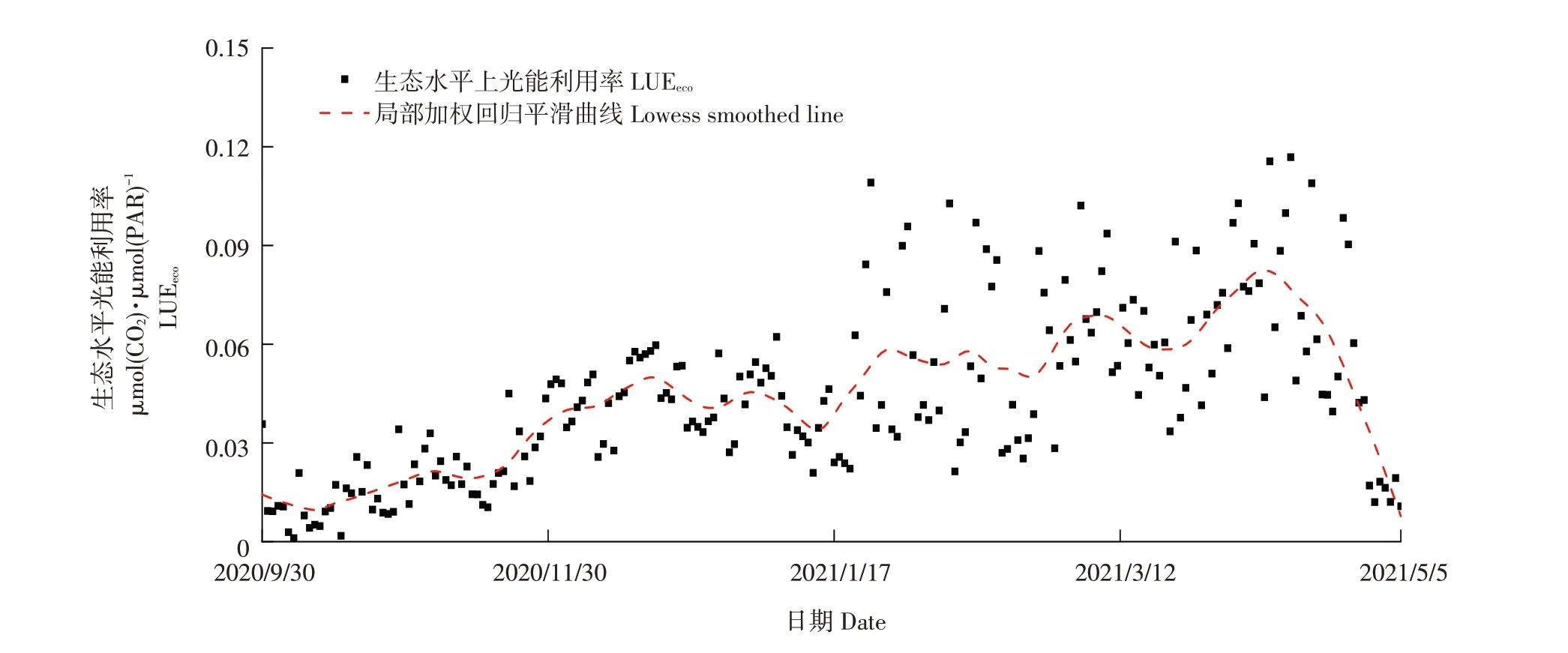
图4 油菜全生育期LUEeco日变化Fig.4 Daily variation of LUEeco during the whole reproductive period of rapeseed
2.2 α变化特征
生态系统表观量子效率(α)反映生态系统光合作用过程中潜在光能利用能力[6],为植物对弱光利用效率量度。本研究利用式(4)和式(5)计算得到α在不同月份变化特征,见表1。油菜生长季α值月变化表现为先升后降,呈抛物线变化趋势,与LUEeco月变化具有相似性。α值随生育进程逐渐上升,3月达到最大值,此时油菜处于花期,植株长至最高且植被覆盖度高,良好冠层结构有利于植株吸收来自各方向散射光[16],α为0.1375 μmoL(CO2)·μmoL(PAR)-1,之后随冠层对弱光利用率逐渐下降,α也随之下降。这一变化反映油菜生长发育从缓慢到旺盛,最后衰老趋势。α值变化范围为0.0387~0.1375 μmol(CO2)·μmol(PAR)-1,平均值为0.0788 μmol(CO2)·μmol(PAR)-1,较低表观量子效率出现在10月及次年5月。

表1 不同月份油菜GPP和PAR拟合参数Table 1 Fitting parameters of GPP and PAR in oilseed rape at different growth stages
2.3 LUEeco和α相互关系
基于涡度相关通量数据估算得到的光能利用率主要以LUEeco和α两类为主,但在前人研究中,大多针对LUEeco和α进行单独动态变化分析,二者之间关系探讨较少,本研究发现,LUEeco和α月变化均表现为单峰曲线,与油菜生长发育联系密切,因此本研究从月份尺度上初步探究二者内在联系。由图5可知,α数值明显高于LUEeco,LUEeco和α表现显著线性关系,即随α增加,LUEeco也逐渐增加。如果将两者拟合线强制通过零点(图中蓝线),发现两者线性关系变为LUEeco=0.5219α,即LUEeco接近α二分之一。
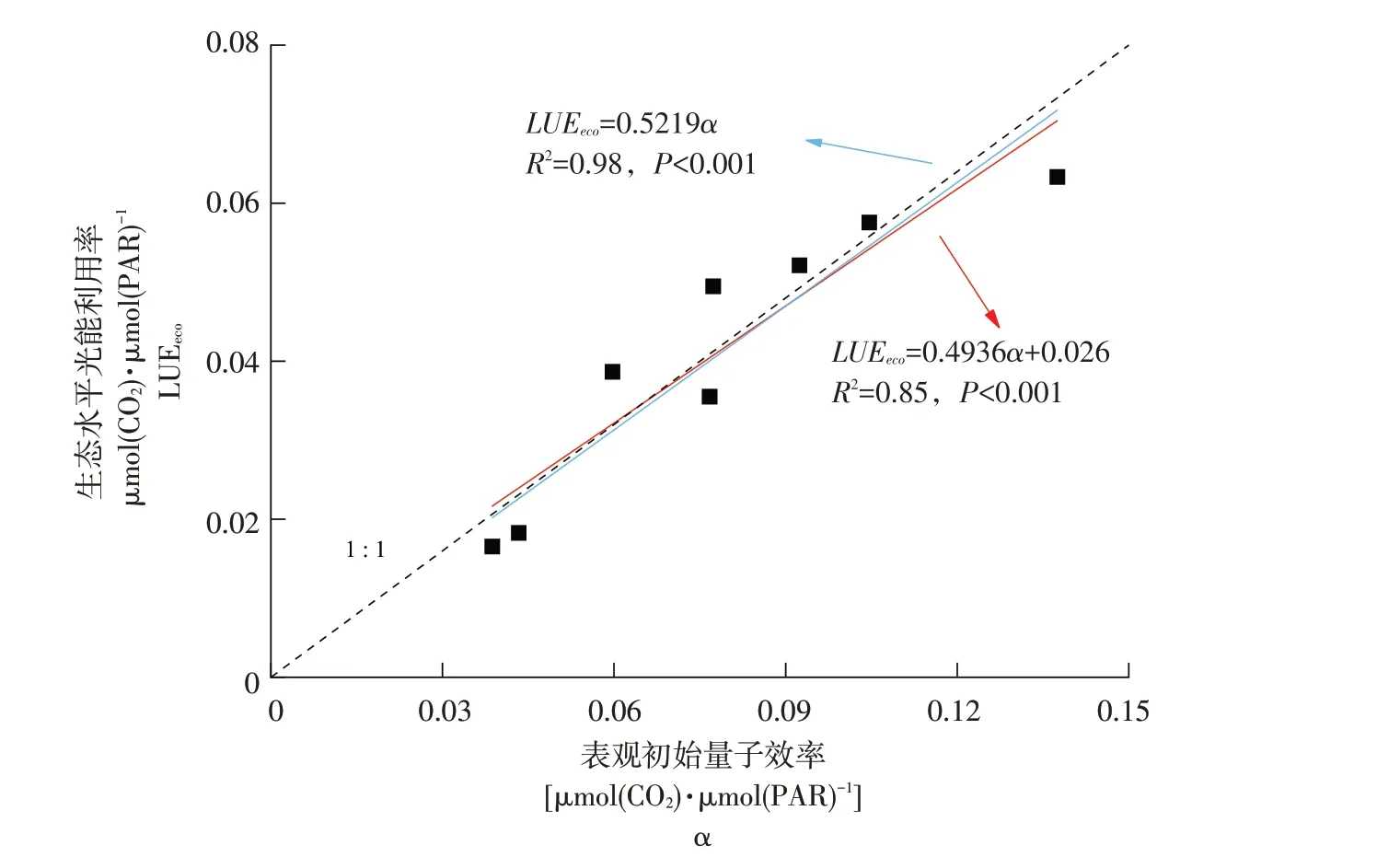
图5 不同月份LUEeco和α关系Fig.5 Relationship between LUEeco and α for different months
2.4 试验地环境因子和NDVI季节变化
利用常规气象观测数据与近地面遥感观测数据,得到研究区环境因子与归一化植被指数(NDVI)季节变化特征(见图6)。
由图6可知,空气温度(Ta)和土壤温度(Ts)具有极显著相关性,油菜生长季平均气温为12.86℃,平均土温为15.37℃;冠层NDVI表现为先升后降抛物线变化,在油菜薹期及花期表现较平稳;油菜生育期内光合有效辐射(PAR)和饱和水汽压差(VPD)波动均较大,光合有效辐射总量为2 481.25 moL·m-2,日均值为11.36 moL·m-2·d-1,日平均饱和水汽压差为0.28 KPa。

图6 研究区环境因子和植被生长季节变化Fig.6 Seasonal variation of environmental factors and vegetation growth in the study area
2.5 LUEeco季节变化影响因子
由于观测期内表观量子效率样本量较少,本研究仅探讨季节尺度上LUEeco影响因子。由表2可知,日尺度上LUEeco与PAR、Ta、Ts、VPD和NDVI均存在极显著相关关系,LUEeco与PAR、Ta、Ts、VPD均呈现极显著负相关,与NDVI呈现极显著正相关,各因子与LUEeco简单相关系数排序依次为PAR>NDVI>VPD>Ts>Ta。因各因子对LUEeco作用并非是简单线性关系,不同因子间相互组合、相互作用,对LUEeco造成不同程度影响,仅依靠简单相关系数无法探究其内在规律联系,因此进一步对LUEeco与5个因子进行多元线性逐步回归分析,去除影响不显著因子,建立LUEeco最优回归方程,结果如表3所示。模型3是最优模型,根据多元线性逐步回归结果建立回归模型为LUEeco=-0.098-0.002PAR+0.192NDVI+0.001Ta,模型复相关系数为0.792,决定系数为0.627,调整系数为0.622。
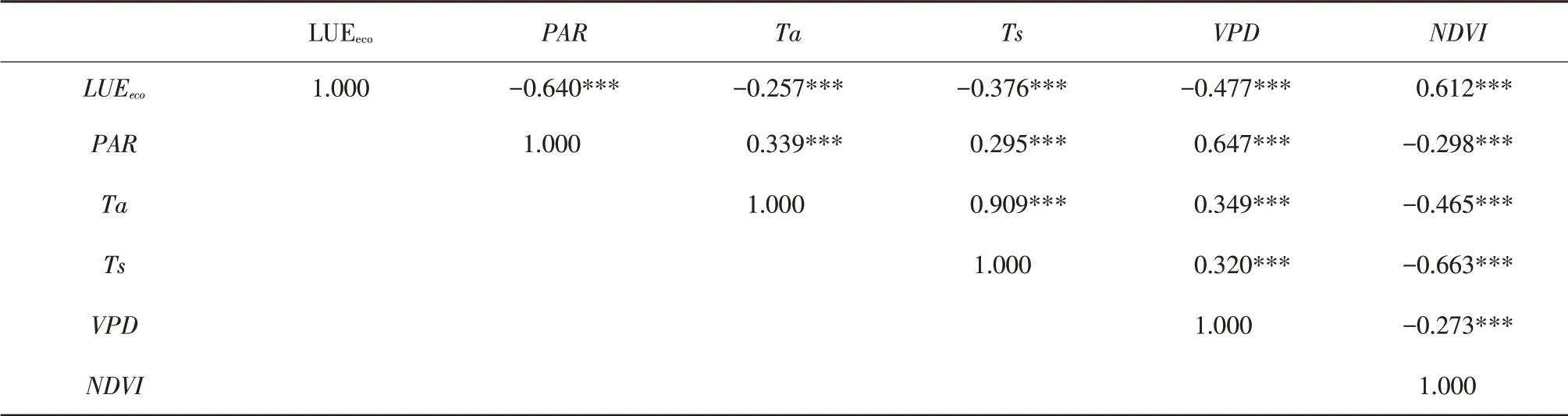
表2 LUEeco与环境因子和NDVI简单相关系数Table 2 Simple correlation coefficients of LUEeco with environmental factors and NDVI
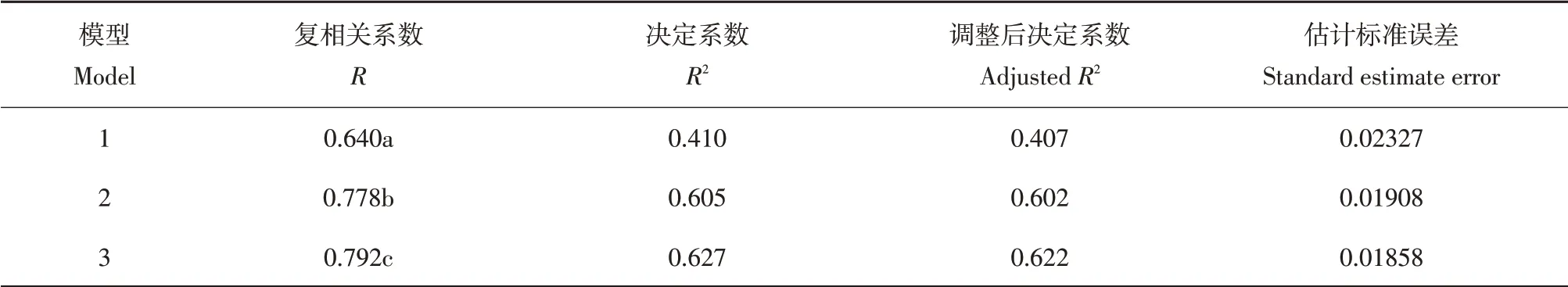
表3 多元线性逐步回归方程模型结果Table 3 Multiple linear stepwise regression equation model overview results
由表4可知,PAR、NDVI和Ta,3个变量回归系数均达到极显著水平(P<0.01)。从3个因子对LUEeco回归系数看,对LUEeco变化影响作用最大的是PAR(-0.541),Ta决定作用最小(0.173)。

表4 回归方程系数显著性检验Table 4 Significance test of coefficients in regression equation
为明确PAR、NDVI和Ta对LUEeco影响的途径和大小,进一步对其作通径分析,LUEeco与主要影响因子PAR、NDVI和Ta的通径分析见表5,由此可合成LUEeco变化通径分析如图7所示。各因子对LUEeco作用由直接作用与间接作用构成,其对LUEeco的综合作用用决策系数大小衡量。由图表可知,LUEeco决定因子直接通径系数绝对值表现为:PAR>NDVI>Ta,说明PAR对LUEeco变化的直接作用最大,其中PAR对LUEeco直接通径系数为负数,表明其对LUEeco的影响为负效应,NDVI和Ta对LUEeco的直接通径系数为正数,表明其对LUEeco的影响为正效应。PAR、NDVI和Ta直接通径系数均大于各自间接通径系数之和,说明3个因子对LUEeco的影响主要体现在直接作用上。决策系数排序依次为PAR>NDVI>Ta,表明PAR对LUEeco变化的综合决定能力更大。
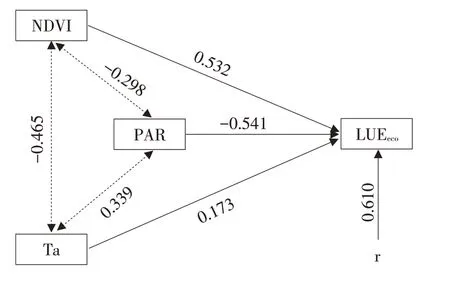
图7 LUEeco的通径分析Fig.7 Path analysis diagram of LUEeco

表5 环境因子和NDVI与LUEeco的通径分析Table 5 Path analysis of environmental factors and NDVI with LUEeco
3 讨论与结论
3.1 光能利用率变化特征
油菜不同时间尺度光能利用率变化特征均具有一定规律性。LUEeco全天日变化呈现U型曲线变化,与前人针对生态水平上光能利用率日动态研究结果一致[16-18]。LUEeco总体表现为早晚较高且存在跳跃式变化,午间LUEeco较低且平稳波动小,由于午间光照强度过高时,植物所接收到光强超出其利用能力,因此发生光抑制现象,影响光能利用率,植物出现“光合午休”现象与午间光照过强密切相关[5]。LUEeco月变化和季节变化与油菜生长发育过程有较大联系,但受环境要素影响,出现波动。本研究中油菜生长季平均LUEeco为0.0347 μmol(CO2)·μmol(PAR)-1,若将其换算为百分制则为0.946%,当前针对油菜实际光能利用率研究发现其仅为0.615%~1.056%,低于水稻、小麦、大豆等作物[12],本研究结果在这一范围内。表观初始量子效率(α)又称为表观初始光能利用率,表征植被潜在最大光能利用能力。本研究中,LUEeco值均低于α值,α和LUEeco之间表现为极显著线性正相关,总体上LUEeco约为α的二分之一,与张雷明等对长白山阔叶红松林的研究结果类似[6],说明实际光合效率与潜在光合效率之间存在差异,本研究仅分析针对月份尺度上两者关系,后续研究中还需考虑使用高频光能利用率数据进行关系探讨。可观察到,LUEeco和α值在1月均存在明显下跌,因为一般对于越冬作物而言,在其营养生长期需经过一段时间较低温度诱导后才转为生殖生长,1月时油菜正处于越冬期,也是油菜由营养生长阶段转变为生殖生长必经阶段,此时光能利用率较低,符合其生长状况。
3.2 光能利用率影响因素
影响光能利用率因素较多,除受植物生理功能影响,还因外界条件改变而出现较大程度变异。油菜作为一种C3作物,其光能利用效率较低,研究表明,C4植物光合能力显著高于C3植物[19],这是因两者CO2同化循环途径不同,在光生态适应性上也表现不同,由不同植物生理结构导致。
本研究中,油菜季节尺度上LUEeco动态变化受到光合有效辐射、温度和作物长势协同调控,3个因子可综合解释62.2%LUEeco变化原因。光合有效辐射作为植物进行光合作用能量来源,无论日尺度上还是季节尺度上光能利用率均会因光照条件改变而改变,此外,由于入射辐射由直射光和散射光两部分构成,研究证实植被对散射辐射利用效率相较于直射辐射更高[20]。归一化植被指数反映植被冠层结构特征,当植物生长旺盛时,植被冠层拦截到光合有效辐射更充足,光合作用面积更大,光能利用能力越强。本研究中,归一化植被指数作为重要遥感植被指数之一,与LUEeco存在极显著相关关系,能较好解释LUEeco动态变化原因,说明使用遥感观测数据可较好描述光能利用率变异特征。温度会通过影响植物光合作用过程中各种酶系统活性进而对光合作用造成显著影响,费敦悦等探讨安徽寿县水稻光能利用率与温度关系,结果表明,日动态中光能利用率与温度有较好的二次函数关系,在季节变化中LUE与温度大致表现为线性正相关[10]。也有研究显示,较大空间尺度上光能利用率与温度相关性并不显著[21]。植物生长环境水分亏缺和水分过多均会直接或间接影响植物光合作用[22],饱和水汽压差是表征环境水分状况参数之一,本研究中油菜生长季饱和水汽压差并非影响LUEeco变化主要因子,但VPD和LUEeco之间呈现显著负相关关系,即LUEeco随VPD增加而降低,因为当VPD过高时植物为减少水分流失而关闭气孔,导致气孔导度降低,影响LUEeco。除本研究中已探讨外界条件,空气中二氧化碳和氧气浓度、植物本身养分状况、土壤肥力均对植物光能利用效率造成一定程度影响[23-24]。
当前研究中,并未完全区分植物本身生理性质还是对外界条件响应引起光能利用率具体改变,因为光能利用率动态变化是内外部因素协同作用结果,同种作物不同品种间及同种作物在不同环境条件下均存在差异,因此探讨光能利用率变异特征和控制机制对于优化基于光能利用率生产力估算模型、提高模型估算精度具有重要意义。本研究仅针对一季油菜开展观测试验,在后续研究中还需开展对比及长时序观测试验。受时空分辨率和地面覆盖程度限制,仅依靠地面涡度观测数据难以进行不同时空格局光能利用率研究,而地面光谱测量作为联系遥感数据重要手段,可较好关联涡度数据和遥感数据,后续可将气象数据观测、植被冠层涡度观测及地面光谱观测相结合,进行涡度数据-气象因子-遥感信息联合光能利用率数据综合研究,以期为大尺度遥感定量研究提供丰富数据资源和参考依据。
