盐含量对餐厨垃圾好氧堆肥腐殖化过程及微生物演变的影响
2022-02-06沈玉君罗文海丁京涛孟海波周海宾程红胜徐鹏翔
刘 娟,沈玉君,罗文海,丁京涛,孟海波,周海宾,3,,张 曦,程红胜,王 健,徐鹏翔
盐含量对餐厨垃圾好氧堆肥腐殖化过程及微生物演变的影响
刘 娟1,2,4,沈玉君1,2,4,罗文海2,丁京涛1,2,4,孟海波1,4,周海宾1,2,3,4※,张 曦1,2,4,程红胜1,4,王 健1,4,徐鹏翔1,4
(1. 农业农村部规划设计研究院农村能源与环保研究所,北京 100125;2. 中国农业大学资源与环境学院,北京 100193;3. 新疆农垦科学院农田水利与土壤肥料研究所,石河子 832000;4. 农业农村部资源循环利用技术与模式重点实验室,北京 100125)
含盐量过高会对堆肥有机质腐殖化过程产生抑制作用,但作用规律仍不清楚。该研究以餐厨垃圾为研究对象,以不添加盐分的处理为对照,设置添加食用盐的处理作为试验组(添加质量分数分别为0.5%、1%和1.5%),进行好氧堆肥,研究不同盐分含量对基本腐熟度指标、有机质组成、腐殖质(Humus,HS)的影响,并结合微生物群落结构和相关性统计分析阐明其作用规律。结果表明,4个处理温度、碳氮比(C/N)、有机质组成等达到腐熟要求,但盐分添加提高了堆体电导率(Electrical Conductivity,EC),添加比例达到1.5%时,高温期缩短至13 d,种子发芽指数(seed Germination Index,GI)降低至65.5%,总有机物降解率降低6.5%,有机质腐殖化过程受到限制。高通量测序和相关性分析的结果表明,添加1.5%盐分主要通过抑制高温双岐菌()、糖单孢菌()和曲霉()、毛孢子菌()的活性,降低总糖、木质纤维素等有机物质的降解和后续HS形成,从而限制餐厨垃圾堆肥过程中腐殖化效果的提升。该研究将为餐厨垃圾等农村有机废弃物处理技术应用提供理论指导。
微生物;好氧堆肥;腐殖化;餐厨垃圾;盐分
0 引 言
经济的快速发展导致生活垃圾产量不断增加。据报道,2021年中国农村生活垃圾产量约为1.76×108t,其中餐厨垃圾的占比达到20%以上,且呈现逐年增加的趋势[1-2]。餐厨垃圾含水率高,有机物丰富,极易腐败发臭变质。不当处理会产生渗滤液和有害气体,导致病原微生物等有毒有害物质滋生,污染水体环境,制约农村人居环境改善提升[3]。近年来,中国高度重视农村人居环境整治,出台了《农村人居环境整治提升五年行动方案(2021-2025年)》,对农村有机废弃物就近处理和资源化利用提出了明确要求。同时,农村生活垃圾分类回收和资源化也会促进餐厨垃圾等的处理利用需求。由于好氧堆肥技术具有无害化、资源化效果好等优点,已成为餐厨垃圾处理利用的研究热点[4]。在堆肥过程中,有机质腐殖化是评价其技术应用和产品质量的重要指标。因此,有必要对餐厨垃圾堆肥过程中有机质降解和腐殖质形成进行研究。
值得注意的是,中国餐厨垃圾盐分含量较高,一般为0.8%~1.5%(湿基),致使堆肥技术应用受到一定限制[5]。较高浓度的盐分会增加渗透压,降低微生物代谢酶的活性,影响微生物生长[6]。目前关于盐分对有机废弃物处理过程的影响研究,大部分学者关注厌氧发酵过程,如餐厨垃圾与废活性污泥共发酵过程,添加高水平的氯化钠(NaCl)(16 g/L)抑制了微生物生长,降低了关键酶活性,如CoA转移酶,辅酶F420等,从而减少短链脂肪酸的生成和甲烷(CH4)的形成[7]。Wang等[8]研究发现,在高盐浓度下,产甲烷菌比水解细菌更敏感,导致较低的CH4产量。现阶段只有少量研究分析了盐分对好氧堆肥的影响。任连海等[9]发现高盐影响餐厨垃圾堆肥效率,高温持续时间较短,不利于灭菌,微生物代谢酶活性降低,产水较少且水分损失多,导致含水率降低至36.18%,堆体pH值低于5.5,水溶性碳氮比下降缓慢。Wu等[10]在厨余垃圾蚯蚓堆肥中指出,盐分抑制有机物的降解和蚯蚓的生命活动,高盐导致蚯蚓细胞渗透压失衡而脱水,不利于铵态氮(NH4+-N)向硝态氮(NO3--N)转化,影响堆肥过程。此外,餐厨垃圾盐分中的钠离子(Na+)和氯离子(Cl-)对种子胚胎生存力产生毒性,影响种子萌发。于兆友等[11]研究发现高盐胁迫对皂荚种子萌发有明显地抑制作用,导致植物体内代谢紊乱。然而,盐分对餐厨垃圾堆肥有机质腐殖化过程的影响研究较少,高盐胁迫下微生物的变化规律至今鲜有报道。
本研究以餐厨垃圾为堆肥主料,研究不同含盐量对堆肥基本腐熟度指标、有机质组成及腐殖质(Humus,HS)的影响,结合微生物群落结构演替和相关性分析,揭示盐分对堆肥有机质腐殖化过程的作用规律,以期为餐厨垃圾盐分调控和资源循环利用提供理论支撑。
1 材料与方法
1.1 试验原料
本研究采用的餐厨垃圾取自农业农村部规划设计研究院食堂,人工剔除无机组分,其中蔬菜45.8%、主食18.2%、水果13.6%、肉类9.4%,其他组分13.0%(湿质量)。经测定,餐厨垃圾的盐分含量为0.21%。玉米秸秆取自江苏省连云港市某公司,粉碎至长度1~2 cm。食用盐购于中国盐业股份有限公司,NaCl质量分数≥97%。基本理化性质如表1所示。
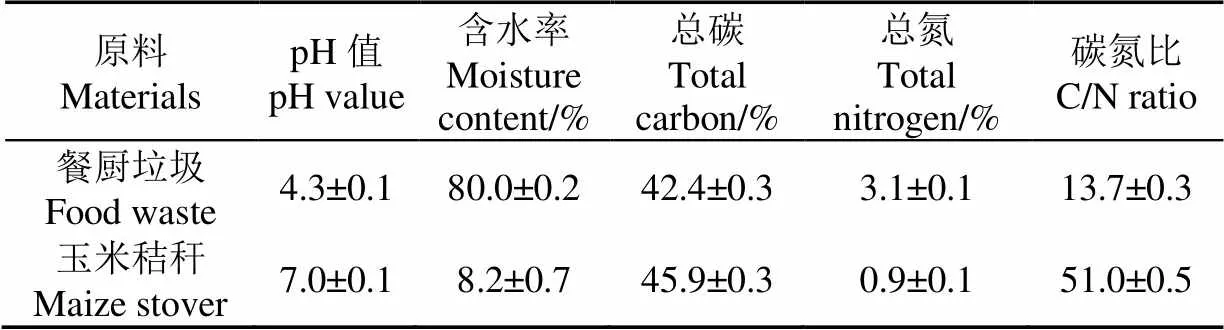
表1 堆肥原料的基本理化性质
1.2 试验设计
本研究共设置4个处理,餐厨垃圾与玉米秸秆按4∶1(湿重比)混合均匀,调节初始含水率至63%左右进行好氧堆肥。以不添加盐分的处理(T1)为对照,添加湿基质量分数分别为0.5%(T2)、1.0%(T3)、1.5%(T4)的食用盐作为试验组(见表2)。整个堆肥过程在60L的密闭式好氧发酵罐中进行(图1),发酵罐的结构如周海宾等[12]所示。堆肥周期为45 d,通风方式采用间歇通风,每隔10 min通1 min,通风量为1.0 L/(kg·min)。在第0、3、7、14、21、28、35和45 天进行翻堆和取样,固体样品每次取500 g混合均匀,一部分立即用于含水率的测定。其余样品分为3份,一份储存在4 ℃冰箱,用于测定基本腐熟度指标,一份自然风干,经粉碎后进行有机质及HS组分的测定,剩余部分冷冻于超低温冰箱(−80 ℃)用于微生物群落结构分析。其中,基本腐熟度指标、有机质和HS组分在测定时均设置了3个重复。

表2 试验设置
注:a为基于湿基质量。
Note:ais based on wet weight.
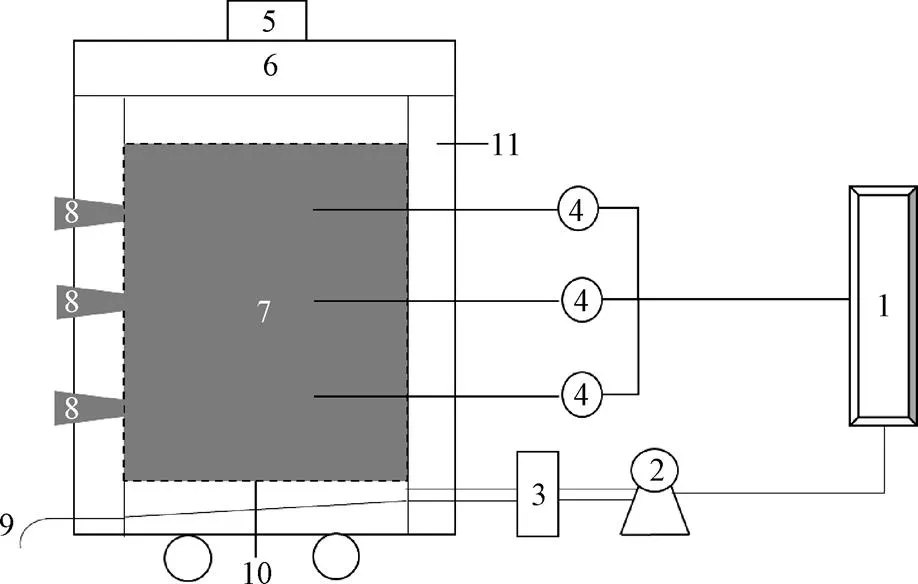
1.通风控制系统 2.气泵 3.流量计 4.温度传感器 5.出气口 6.密封盖 7.堆肥物料 8.固体采样口 9.渗滤液排出口 10.筛网 11.保温层
1.3 样品测定
温度采用温度传感器(SBWZ-2460,上海虹天,中国)进行实时采集。新鲜样品与去离子水按固液比1∶10充分混合后静置过滤,所得滤液由pH计(SX-610,上海三信)和电导率仪(DDS-307A,上海精科雷磁,中国)分别测定pH和电导率(Electrical Conductivity,EC)。盐分含量采用电导法测定[13]。种子发芽指数(seed Germination Index,GI)的测定方法是取上述滤液5 mL于盛有10粒小白菜种子的培养皿中,置于25±1 ℃培养箱中培养72 h,测定根长和种子发芽率。采用烘干法(105 ℃,8 h)测定含水率。总碳(Total Carbon,TC)、总氮(Total Nitrogen,TN)含量使用元素分析仪(PE2400,PerkinElmer,美国)测定。
采用灼烧法(550 ℃,4 h)测定挥发性固体(Volatile Solid,VS)。溶解性有机物(Dissolve Organic Matter,DOM)用总有机碳分析仪(TOC-L SSM-5000A,岛津,日本)测定。蛋白质和脂肪分别采用GB 5009.5-2016和GB 5009.6-2016测定。纤维素、半纤维素、木质素的测定采用Van Soest法[14]。总糖采用3,5-二硝基水杨酸比色法测定。HS组分的测定参考蒋志伟[15]的方法。DOM提取液在465和665 nm处的吸光度比值E4/E6和DOM提取液在254 nm处的吸光度乘以100与溶液TOC值之比SUVA254采用佟影影[16]的方法测定。三维荧光光谱采用荧光分光光度计(F-7000,Hitachi,日本)进行测定,具体设置如下:E:250~600 nm,扫描间隔2 nm,E:200~400 nm,扫描间隔10 nm,扫描速率:1 200 nm/min。
利用高通量测序测定微生物群落组成。采用土壤基因组(DNA)提取试剂盒(FastDNA® Spin Kit for Soil, MP Biomedicals, Solon, USA)对获得的沉淀物进行DNA提取。提取的DNA浓度和质量通过NanoDrop2000和1%琼脂糖凝胶电泳确定。利用引物338F(5ʹ- ACTCCTACGGGAGGCAGCAG-3ʹ)和806R(5ʹ- GGACTACHVGGGTWTCTAAT-3ʹ)通过聚合酶链反应(PCR)扩增细菌16S rRNA的V3~V4高可变区。选择引物ITS1F(5ʹ-CTTGGTCATTTAGAGGAAGTAA-3ʹ)和ITS2R(5ʹ-GCTGCGTTCTTCATCGATGC-3ʹ)通过PCR扩增真菌ITS的ITS1区,并在Illumina MiSeq PE300平台(上海美吉生物医药科技有限公司)完成测序。
1.4 数据分析
使用Excel 2019对数据进行汇总整理,利用SPSS 22.0中的单因素方差(ANOVA)分析进行显著性差异检验。利用Canoco 5.0进行冗余分析,确定堆肥过程中优势菌群(细菌、真菌)与基本腐熟度指标、有机质、HS组分之间的相关性,其余图像均使用OriginPro 2021绘制。
2 结果与讨论
2.1 基本腐熟度指标
4个处理堆体温度均表现出相同的趋势,经历了升温、高温、降温和腐熟4个阶段(图2a)。堆肥初期有机物的生物降解导致各处理温度在前7 d内进入高温阶段(>55 ℃)。随着易降解有机物的消耗,温度逐渐下降。翻堆后堆体温度升高,可能是由于翻堆改善了堆体孔隙结构,并对堆体各层物料进行混合后,促进了有机质降解[3]。堆肥42 d后趋于环境温度,说明充分腐熟。与其他处理相比,T4处理堆肥高温期最短(13 d),这可能是由于添加1.5%的盐分抑制了微生物的生长繁殖,不利于有机物的快速降解[17]。
由图2b可以看出,堆肥初期pH值较低,主要是由于餐厨垃圾贮存过程快速水解,产生乙酸、丁酸等有机酸[18]。所有处理的pH值在21 d内显著上升,之后稳定在8左右直至堆肥结束。pH值上升的主要原因是含氮有机物矿化产生大量NH4+-N[19]。另外,观察到前3 d内T1处理pH值出现小幅下降,而其他3个处理pH值持续升高,说明盐分可能会影响堆肥初期有机物的降解,从而减少有机酸的产生。
堆肥过程中各处理的含水率基本呈现波动下降趋势(图2c),这是由于一方面高温、曝气使堆体水分损失,而另一方面有机质降解也不断产生水分[20]。堆肥结束,T4处理的含水率下降幅度较大,明显高于T1~T3处理(<0.01),说明盐分添加量达1.5%时可以加快含水率的下降,原因可能是高盐导致微生物代谢活动受阻严重,有机质降解缓慢,产水较少[9]。
各处理的EC值均呈现上升的趋势(图2d),主要是由于有机物分解产生NH4+-N等盐离子,且水分蒸发和气体释放致使堆体总重量减少,形成“浓缩效应”。Li等[21]在餐厨垃圾和花园废物堆肥中也发现了类似的变化。一般认为,EC>4 mS/cm的堆肥产品会对植物生长产生不利影响[22]。经过45 d的堆肥后,T2~T4处理最终的EC值分别为6.7、8.6和8.5 mS/cm,远高于EC的阈值,不利于堆肥腐熟,可能是因为盐分的添加引入了大量的Na+和Cl-,造成无机盐离子累积。
碳氮比(C/N)被看作是评价堆肥腐熟的重要指标。整个堆肥过程所有处理的C/N呈下降趋势(图2e),可归因于堆肥过程有机物的矿化速率高于氨气(NH3)的排放速率[23]。一般认为,C/N < 20时堆肥达到腐熟,但这并不适用于初始C/N较低的餐厨垃圾堆肥。因此,进一步测定了GI值来衡量堆肥腐熟。各处理的GI值整体呈现增加趋势(图2f),可能是由于有机酸、NH4+-N、酚类等有毒物质的释放和HS的形成[24]。堆肥结束,T4处理的GI值最低,仅为65.5%,总有机物降解率降低6.5%,未达《有机肥料》(NY/T 525-2021)的腐熟标准(GI≥70%),说明添加1.5%盐分会抑制种子发芽,不利于堆肥腐熟。
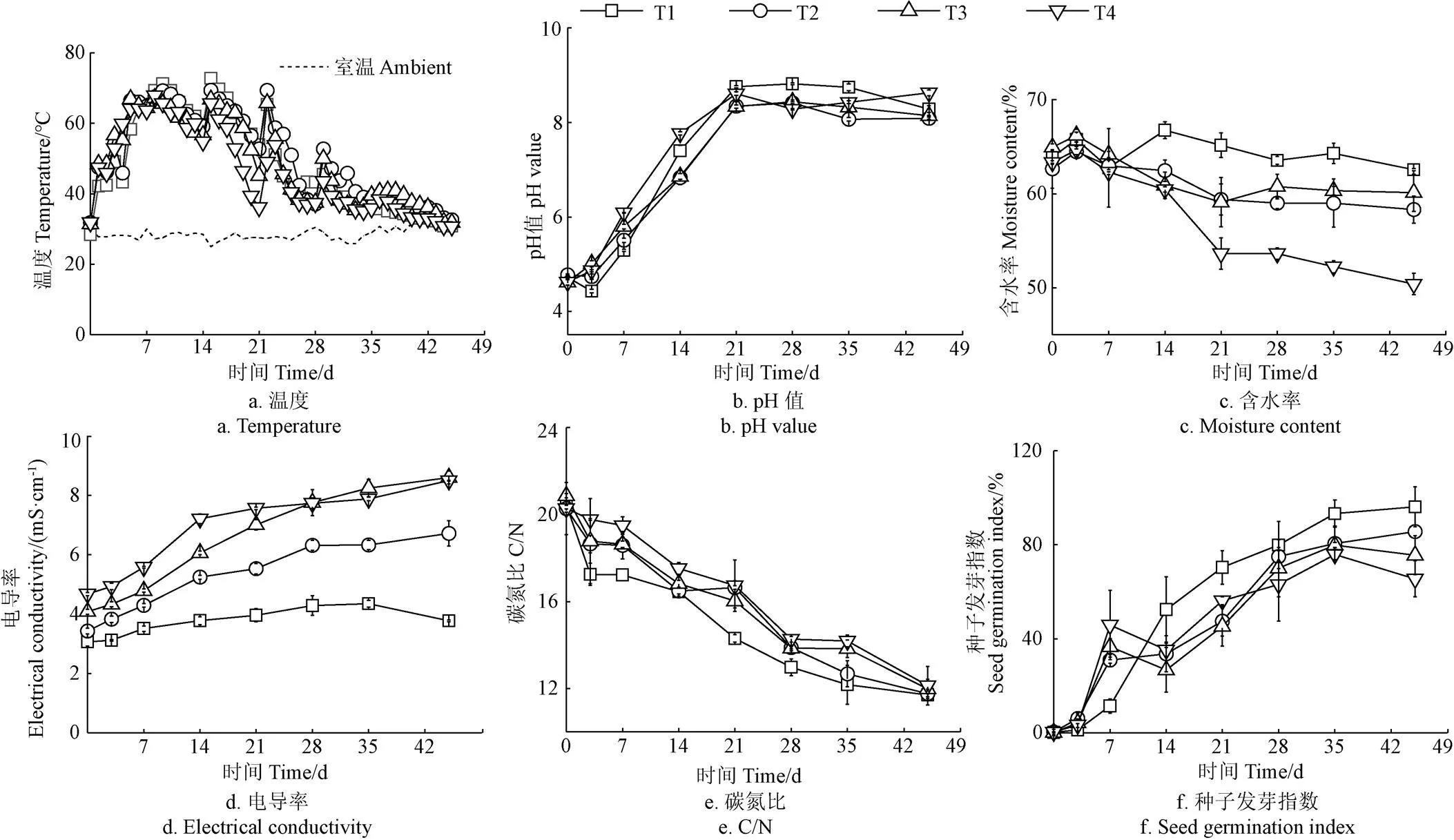
图2 餐厨垃圾与玉米秸秆联合堆肥过程中基本腐熟度指标的变化
2.2 有机质组成变化
随着堆肥过程中微生物的自身利用与矿化损失,4个处理VS含量逐渐下降(图3a)。堆肥结束,T1处理VS降解率达到19.9%,而T2、T3和T4处理VS降解率为16.5%、16.1%和13.3%,说明盐分的加入会抑制有机物降解,特别是盐分添加量达1.5%时抑制作用最强。这一现象与温度变化一致(图2a),可能是由于高盐导致微生物活性受阻,减缓新陈代谢过程,从而限制有机质降解[25]。
整个堆肥过程,4个处理DOM含量逐渐下降(图3b)。其原因是DOM一部分被微生物生长代谢所消耗,一部分被合成稳定的HS。高温阶段(第14 天),T1、T2、T3和T4处理DOM下降明显,分别从48.2 mg/g左右下降至28.1、36.9、40.5和31.7 mg/g,说明DOM被微生物快速降解利用。
图3c为堆肥过程中木质纤维素组分的变化,可以观察到纤维素、半纤维素和木质素表现出不同的变化趋势。总体来说,半纤维素降解率>纤维素降解率>木质素降解率,这是由于微生物会首先利用结构简单的半纤维素和纤维素,随后利用具有高度复杂分子结构的难降解木质素[26]。堆肥结束,T3和T4处理的木质纤维素分别从原来的41.6%、39.9%增加到44.1%、44.2%,可能是因为易降解有机物先被微生物利用转化,从而导致木质纤维素的相对含量有所上升。对比分析发现,T1与T2处理的木质纤维素含量从42.3%、40.0%降低到27.6%、38.4%,且T2处理降解率仅为4.1%,低于T1处理(34.6%)。这些结果说明添加盐分会抑制木质纤维降解菌的代谢和增殖,降低木质纤维素的分解。
如图3d所示,所有处理的蛋白质含量均升高,主要是因为蛋白质分解的同时其他有机物也在分解,产生浓缩效应。堆肥前期,T2~T4处理的蛋白质含量有所下降,可能是由于初期有机氮转化为NH3进一步挥发[27]。堆肥结束,T1~T4处理的蛋白质含量分别为24.7%、19.3%、23.5%和17.0%,T4处理的蛋白质含量显著低于其他处理(<0.01),说明高盐条件(1.5%)不利于蛋白质降解。
堆肥过程中4个处理的脂肪和总糖含量均呈现下降趋势(图3e,图3f)。总糖和脂肪能够被微生物降解作为能量来源或为HS形成提供前体物质[28]。堆肥前期,易降解的有机物较多,微生物代谢旺盛,脂肪和总糖含量迅速下降。堆肥后期,主要降解木质纤维素等难降解的物质,可溶性有机物和干物质减少,微生物活性降低,造成脂肪和总糖降解缓慢。整个堆肥过程,T4处理的脂肪和总糖降解率分别为31.3%和36.5%,显著低于T1处理(<0.05),说明盐分添加比例达1.5%时,微生物的耐受能力较弱,生长代谢受抑制,从而减少脂肪和总糖降解。
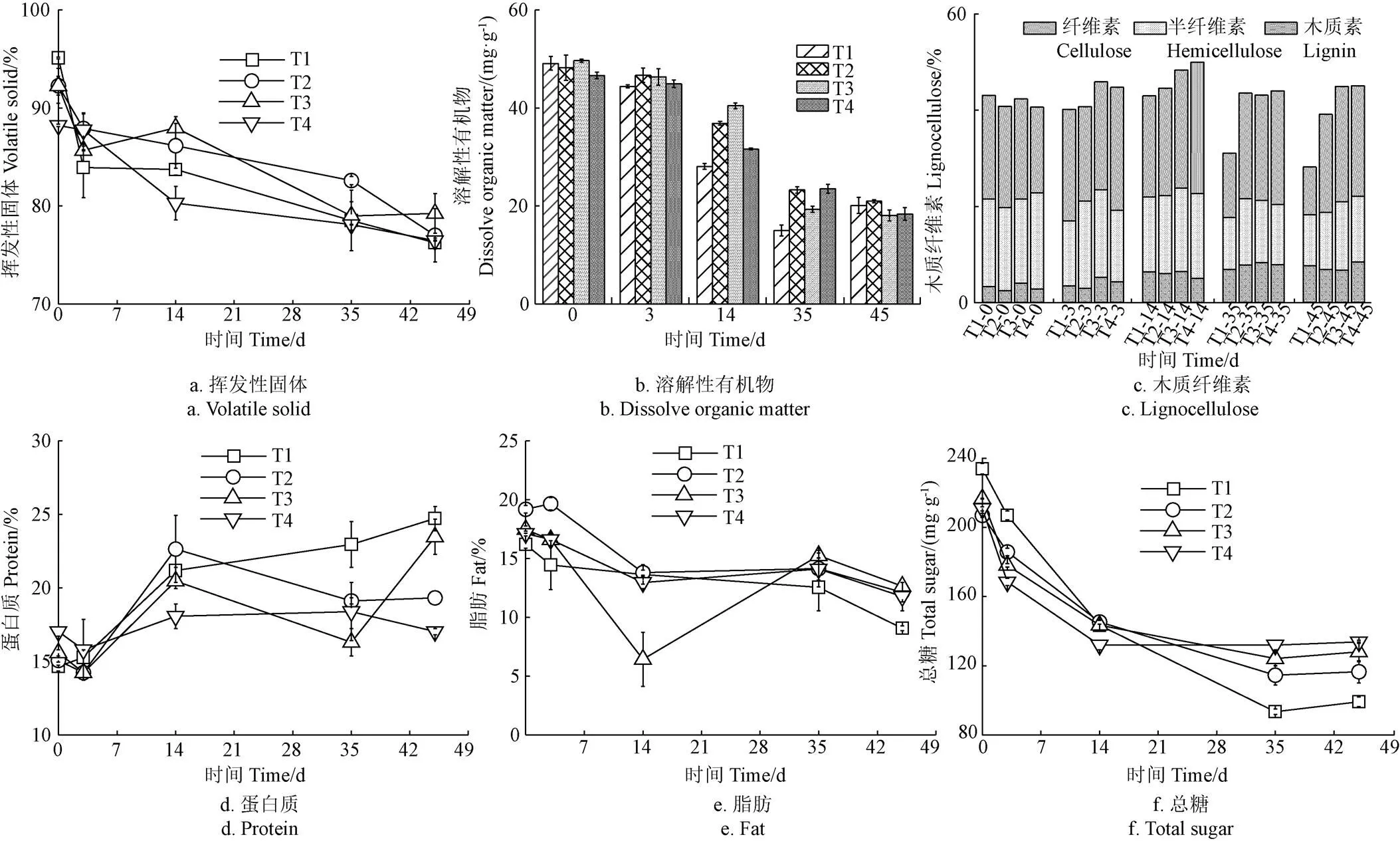
注:挥发性固体、木质纤维素、蛋白质及脂肪以质量分数表示。
2.3 腐殖化过程
堆肥过程中各处理的HS、胡敏酸(Humic Acid,HA)和富里酸(Fulvic acid,FA)含量变化如图4所示。堆肥初期(0~3 d),所有处理的HS含量呈现下降的趋势(图4a),这一结果与玉米秸秆堆肥中HS的变化趋势一致,反映了HS的不稳定化合物减少[29]。堆肥的14~45 d,HS含量逐渐增加,可能是由于有机物降解产生的酚类、醌类、羧酸、氨基酸等前体物质不断合成HS[30]。堆肥结束,T2~T4处理的HS含量显著低于T1(<0.05),说明了盐分的加入不利于HS的合成。这可能是由于盐分的加入抑制了有机物的降解(图3a),减少了HS前体物质的产生。HA作为HS的主要成分,其变化趋势与HS一致,呈现先减少后增加的趋势(图4b)。因此,有研究指出HS的增加也有可能是HA的产生引起的[31]。T1~T4处理最终HA含量分别为83.7、67.3、59.5和54.6 mg/g,相比于第0天,HA含量分别增加了142.2%、117.4%、76.4%和97.3%,表明添加盐分抑制HA的产生。这可能是由于盐分的加入抑制了FA向HA的转化以及微生物对HA的合成。与HA和HS的变化不同,FA含量在整个堆肥过程中呈现下降的趋势(图4c)。堆肥过程微生物可以利用FA为碳源,形成结构更稳定的HA[32]。与堆肥初期相比,T1处理最终FA质量分数降低了28.2 mg/g,略高于T2(25.3 mg/g)、T3(17.6 mg/g)和T4处理(20.3 mg/g),表明盐分的加入抑制FA的转化,从而降低了HA的含量。
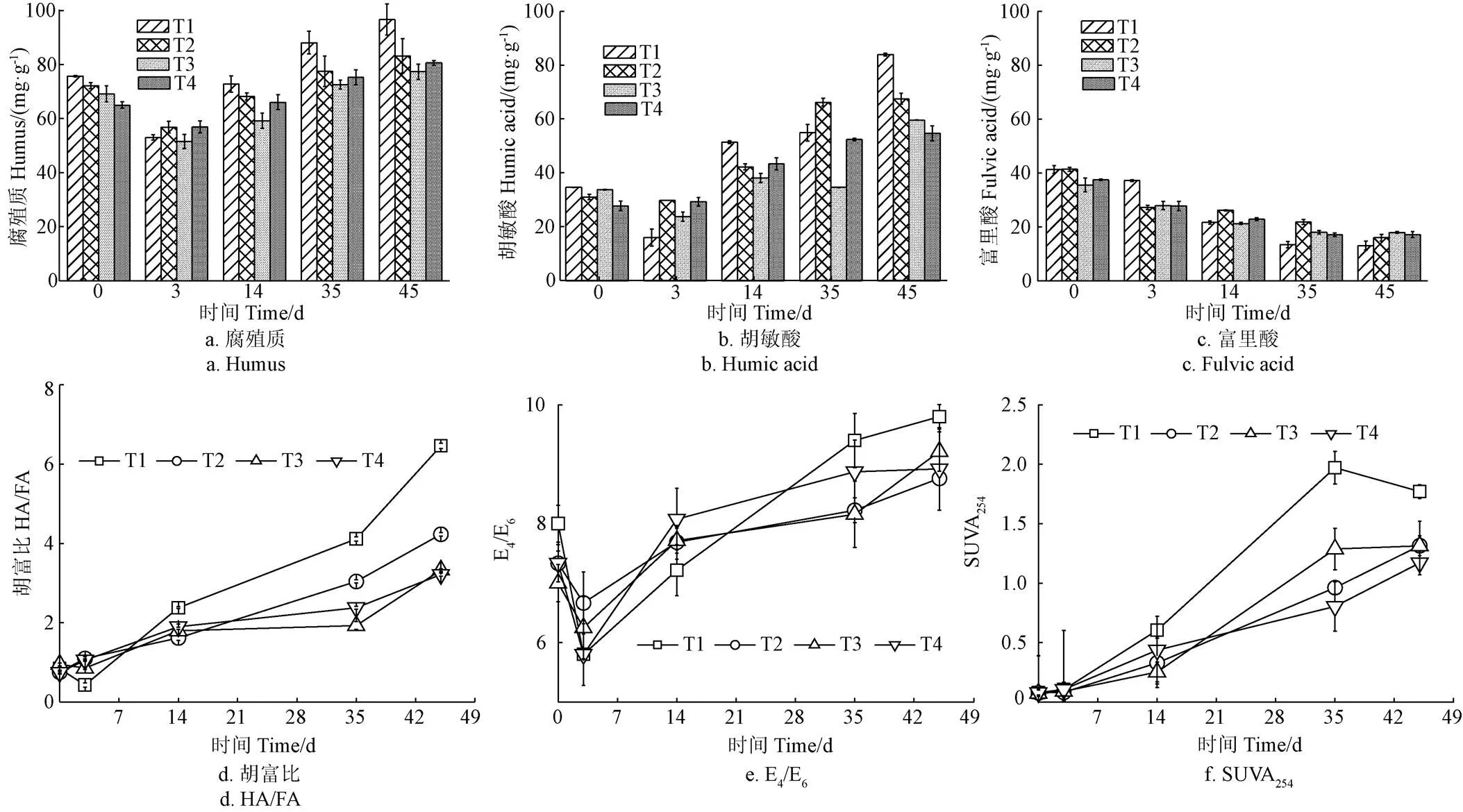
注:E4/E6为 DOM提取液在465 nm 和 665 nm 的吸光度比值。
所有处理的HA/FA在0~14 d缓慢上升,然后在35~45 d显著增加(图4d),反映了堆肥过程中腐殖化程度增强且主要发生在堆肥后期。堆肥结束,T1~T4处理的HA/FA分别为6.5、4.2、3.3和3.2,均超过1.9,表明最终产品的腐熟[33]。堆肥结束,4个处理的腐殖化程度顺序为T1>T2>T3>T4,说明盐分的加入抑制了腐殖化过程,盐分添加量达到1.5%(湿质量)时,抑制作用最强。
为了进一步了解腐殖质结构的变化,采用紫外光谱和三维荧光光谱对其进行表征。由图4e可以看出,所有处理的E4/E6在堆肥前3 d有所降低,可能是由于芳香族化合物初步缩聚或有机物矿化[34],随后呈现上升趋势,与崔东宇等[35]在牛粪堆肥中的研究结果一致。堆肥前后T4处理的E4/E6增加了20.7%,低于T1处理(22.5%),说明盐分添加量达1.5%会减弱生物化学过程,抑制大分子腐殖酸的形成以及芳香族化合物的聚合。随着堆肥的进行,4个处理的SUVA254都有所增加(图4f)。这可能是由于氨基酸与木质素类降解物不断缩合,非腐殖质向HS转化,实现堆肥腐殖化[34]。堆肥结束,T4处理的SUVA254为1.2,低于T1~T3处理(1.3~1.8),说明添加1.5%盐分导致芳香碳含量较低,极性官能团的芳香环较少,腐殖化程度受抑制。
图5a显示了堆肥过程中DOM 的三维荧光光谱图。根据Chen等[36]的研究结果,三维荧光图可分为Ⅰ(类酪氨酸)、Ⅱ(类色氨酸)、Ⅲ(FA类)、Ⅳ(可溶性微生物代谢产物)、Ⅴ(HA类)5个区域。随着堆肥的进行,荧光峰强度逐渐向V区移动,说明产生了稳定的HA类物质。为避免视觉干扰,采用荧光区域积分对各区域荧光强度占比进行定量分析发现(图5b)。随着堆肥的进行,所有处理的P、P及P值均呈下降趋势,反映了堆肥过程可以促进蛋白质类物质和可溶性微生物代谢产物分解,从而形成稳定的HS[37]。值得注意的是,T1~T4处理HS类物质的荧光强度占比(P+P)分别从原来的82.1%、83.8%、82.6%、83.5%增加到96.3%、95.3%、95.7%、94.6%,增加幅度T1>T3>T2>T4,进一步说明高盐条件(1.5%)会抑制蛋白质类物质的分解,影响前体物质向HS转化,从而减少HS的产生,降低腐殖化程度。
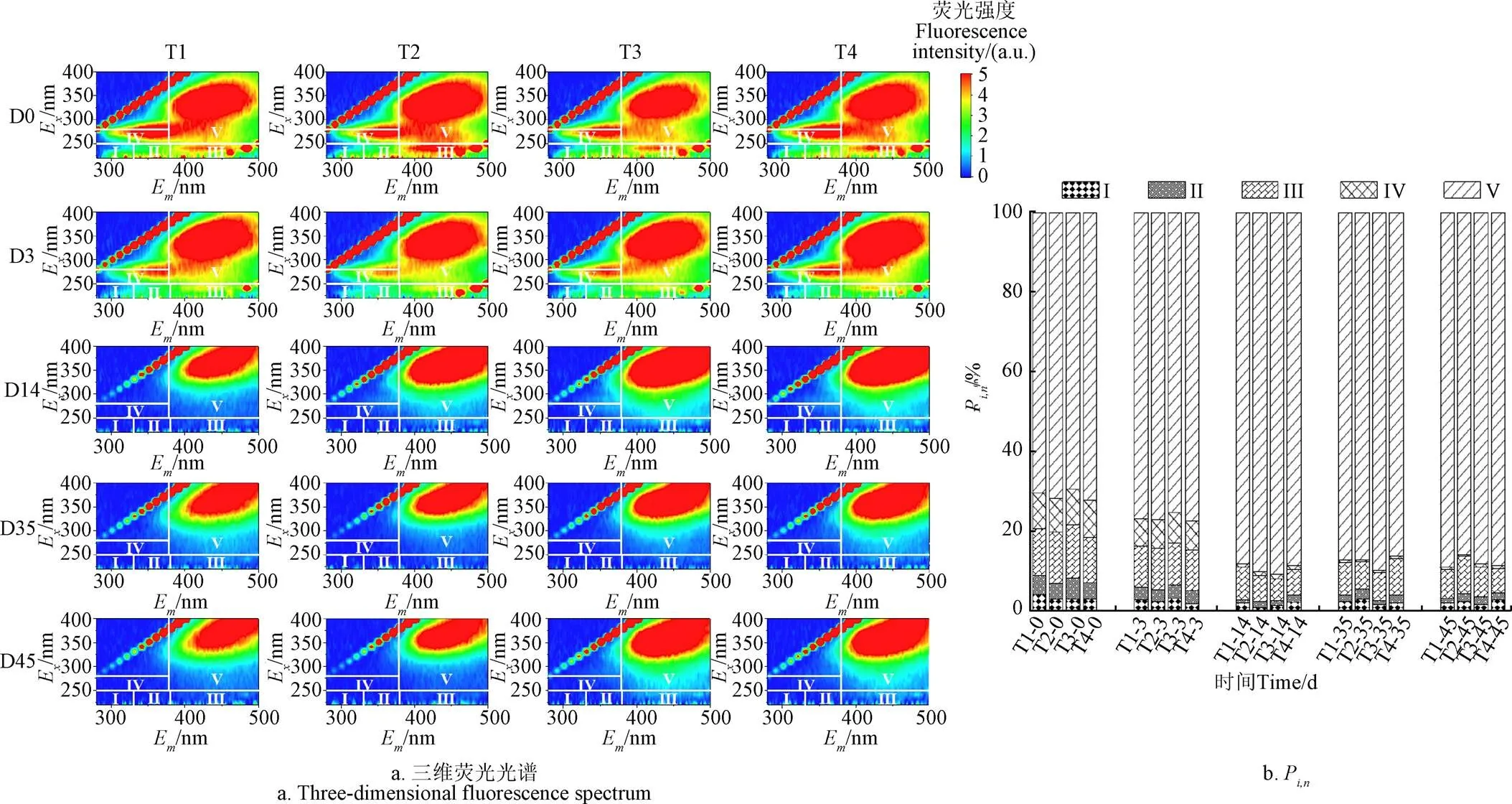
注:0、3、14、35、45为堆肥时间,d。Em为发射波长Emission wavelength 的缩写。Ex为激发波长Excitation wavelength的缩写。Ⅰ ~ Ⅴ分别为类酪氨酸、类色氨酸、类富里酸、可溶性微生物代谢产物和类胡敏酸物质。Pi,n为i区体积积分占整个区域积分(n)的百分比。
2.4 微生物群落演替
2.4.1 微生物群落多样性
如图6所示,所有处理细菌群落的Chao1和Shannon指数在堆肥前后有不同程度的下降,而Simpson指数增加,表明细菌群落的丰富度和多样性降低。该结果与Zhang等[38]在猪粪堆肥中的研究结果一致,可能是因为堆肥初期易降解物质足以维持大量微生物的生长,但是随着大量有机物和水分的消耗,微生物之间的竞争加剧,导致丰富度和多样性降低。其中,各处理的Chao1指数在堆肥后期(即腐熟阶段)有所升高,可能是因为微生物适应了环境条件的改变。堆肥结束,相比于其他3个处理,T4处理的物种丰富度和多样性较低,说明高盐添加量(1.5%)降低了细菌丰富度和多样性。基于各处理的OTU水平进行主成分分析(Principal Component Analysis,PCA),可以进一步解释细菌群落的变化。由图6d可知,PC1和PC2分别解释了25.7%和14.3%的细菌群落差异。沿PC1轴有3个聚集体,从右向左腐熟程度越来越高。PCA分析结果显示,T1与T2~T4处理在第35天和45天距离较远,说明细菌群落差异较大,盐分的添加显著改变了细菌群落的结构,尤其是在降温和腐熟期更为明显。
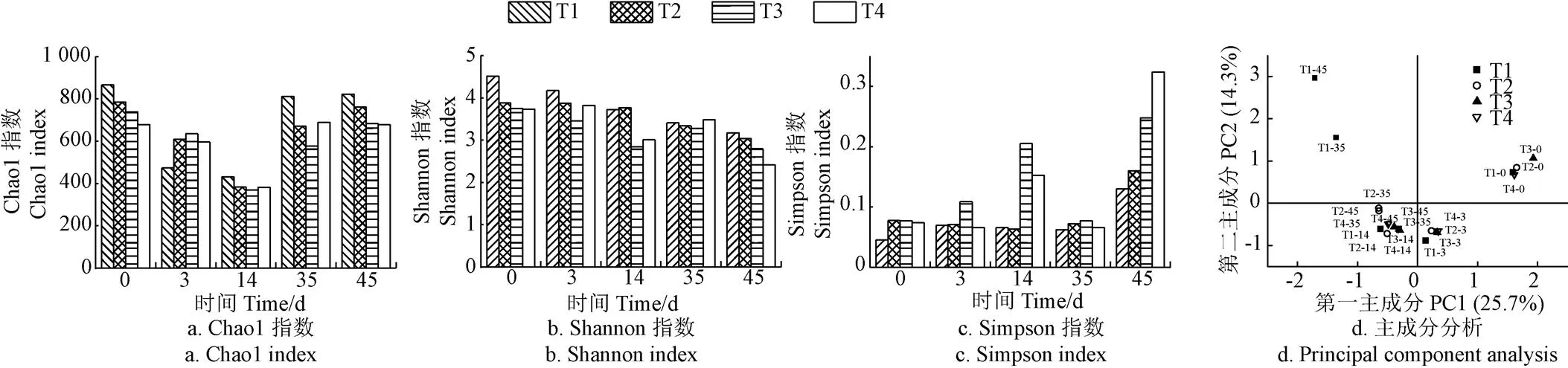
图6 堆肥过程中细菌多样性的变化
与细菌群落相比,真菌多样性相对较低(图7)。所有处理真菌群落的Chao1和Shannon指数在堆肥第3天和第14天出现了下降的趋势,而Simpson指数则相反,表明真菌群落多样性在堆肥升温和高温阶段受到限制。经过45 d的堆肥后,T4处理的Chao1、Shannon指数较低,而Simpson指数较高,表明其真菌多样性较差。基于真菌群落OTU水平进行PCA分析,两个轴共解释了52.2%的真菌群落差异(图7d)。其中,T4处理在第35天和第45天与其他处理距离较远,差异较大,证实了盐添加量达到1.5%时,影响降温和腐熟时期的真菌群落。
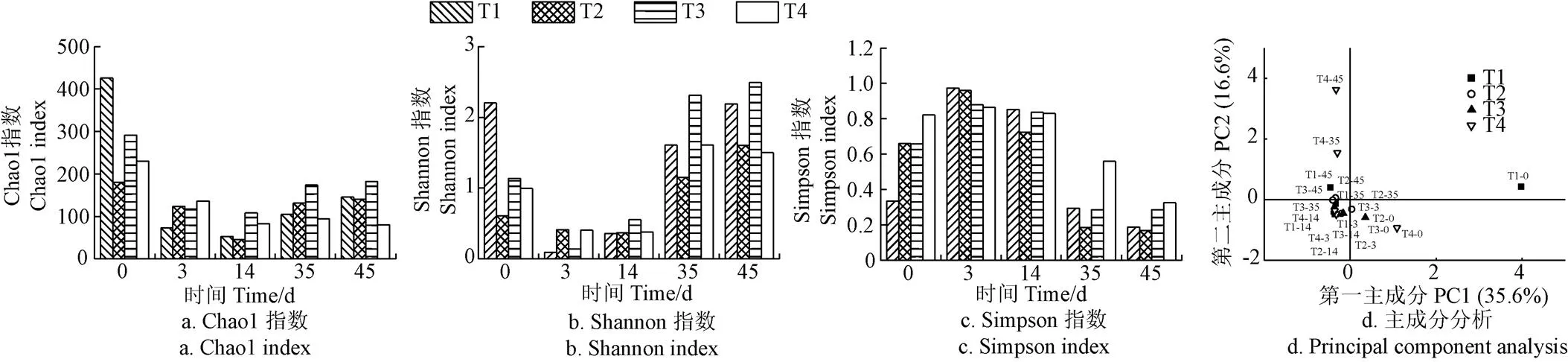
图7 堆肥过程中真菌多样性的变化
2.4.2 微生物群落结构
堆肥过程中不同处理在属水平上的细菌群落变化如图8所示。整个堆肥过程,主要优势属包括芽孢杆菌属()、乳酸杆菌属()、糖单孢菌属()、海洋杆菌属()及其他属水平上的微生物。
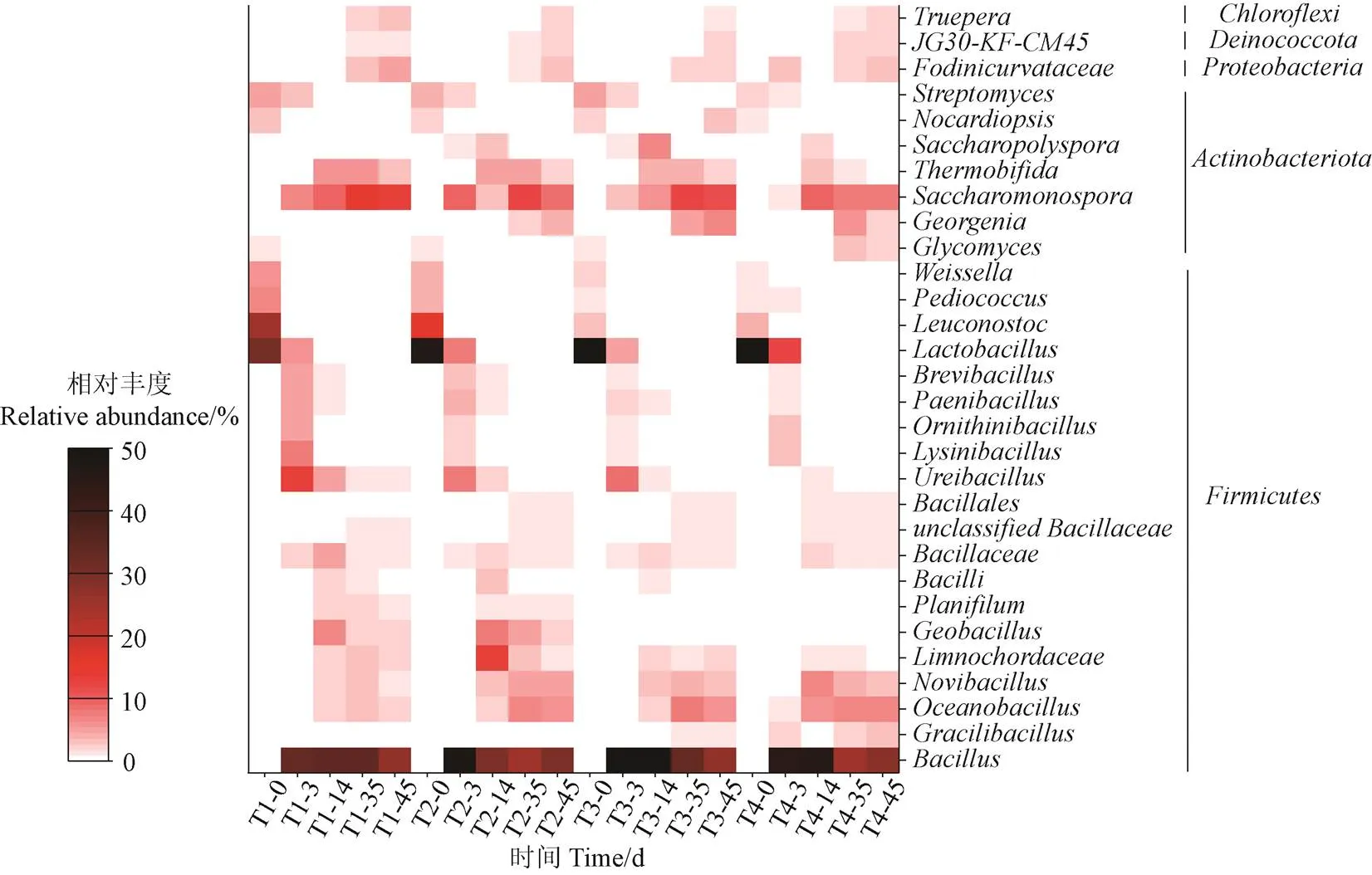
图8 堆肥过程中细菌群落在属水平(前30)上的相对丰度
堆肥初期,各处理的优势属为厚壁菌门()中的,可以降解淀粉、蛋白质、油脂和纤维素等有机物,产生有机酸[39],导致堆肥初期pH值较低(图2b)。相比于T1~T3处理,T4处理的相对丰度较高,说明了其具有耐盐性。随着堆肥的进行,的丰度逐渐降低,可能是因为温度的降低和氧气的增加抑制了其活性。
堆肥升温期,隶属于的和解脲芽孢杆菌属()占据优势。值得注意的是,T4处理的相对丰度仅为0.6%,明显低于T1~T3处理(7.8%~13.0%),说明高盐抑制了升温阶段的活性。堆肥高温阶段,的相对丰度较高,能抵御高温环境,参与有机生物降解[40]。此外,放线菌门()中的高温双岐菌属()在高温期也有所增加,证明了其嗜热性。能够分泌木质纤维素降解酶,加快难降解有机物分解[41]。堆肥结束,4个处理的相对丰度分别为3.4%、2.6%、2.5%和0.5%,说明随着盐分增加,的代谢活动会受到抑制,从而减少木质纤维素的降解(图3c)。相比之下,一些中温菌属,如魏斯氏菌属()、明串珠菌属()、链霉菌属()、拟诺卡菌属()、赖氨酸芽孢杆菌()对温度变化的适应性差导致相对丰度较低。
堆肥后期,除之外,的相对丰度也较高,能够产生纤维素酶、半纤维素和木质素分解酶,将木质纤维素转化为HS[42-43]。与T1 ~ T3处理相比,具有较低丰度的T4处理木质纤维素的降解速度慢,抑制了HS形成(图3c&图4a)。随着堆肥的进行,的相对丰度越来越高,说明该物种能适应堆肥环境的改变。
图9为堆肥过程中真菌群落在属水平上的相对丰度,可以观察到所有处理的优势属为念珠菌()、嗜热真菌()、曲霉()和。
堆肥初期,所有处理子囊菌门()中的丰度较高,可以有效降解内分泌干扰物、烷基苯、壬基酚和多环芳烃[44],但是随着堆肥的进行其相对丰度逐渐下降,这与王秀红等[45]在鸡粪堆肥中的结果类似,说明堆肥过程能去除一部分病原菌,减轻对人体和动物的危害。高温阶段(第14天),所有处理的丰度增加。据报道,能够产生耐热纤维素酶,在木聚糖(半纤维素成分)降解过程中发挥重要作用[46]。在堆肥腐熟阶段(第45天)丰度较高,可以产生胞外酶,负责纤维素和木质素的降解[47]。相较于T1处理(50.9%),T2~T4处理的丰度(1.3%~42.3%)较低,尤其是T4处理,说明盐分含量的增加会抑制的生长代谢。隶属于担子菌门()中的毛孢子菌属()具有广泛的底物利用能力,可以将木质纤维素和外源物质转化为脂质[47]。值得注意的是,在T4处理的相对丰度较低,说明了盐添加量为1.5%时,较敏感,活性较低,不利于木质纤维素的转化。此外,伊萨酵母属()、枝顶孢霉属()、金孢子菌属()、丝壳属()等微生物分布在堆肥的各个时期,对有机物分解和腐殖化过程发挥着重要作用[48]。
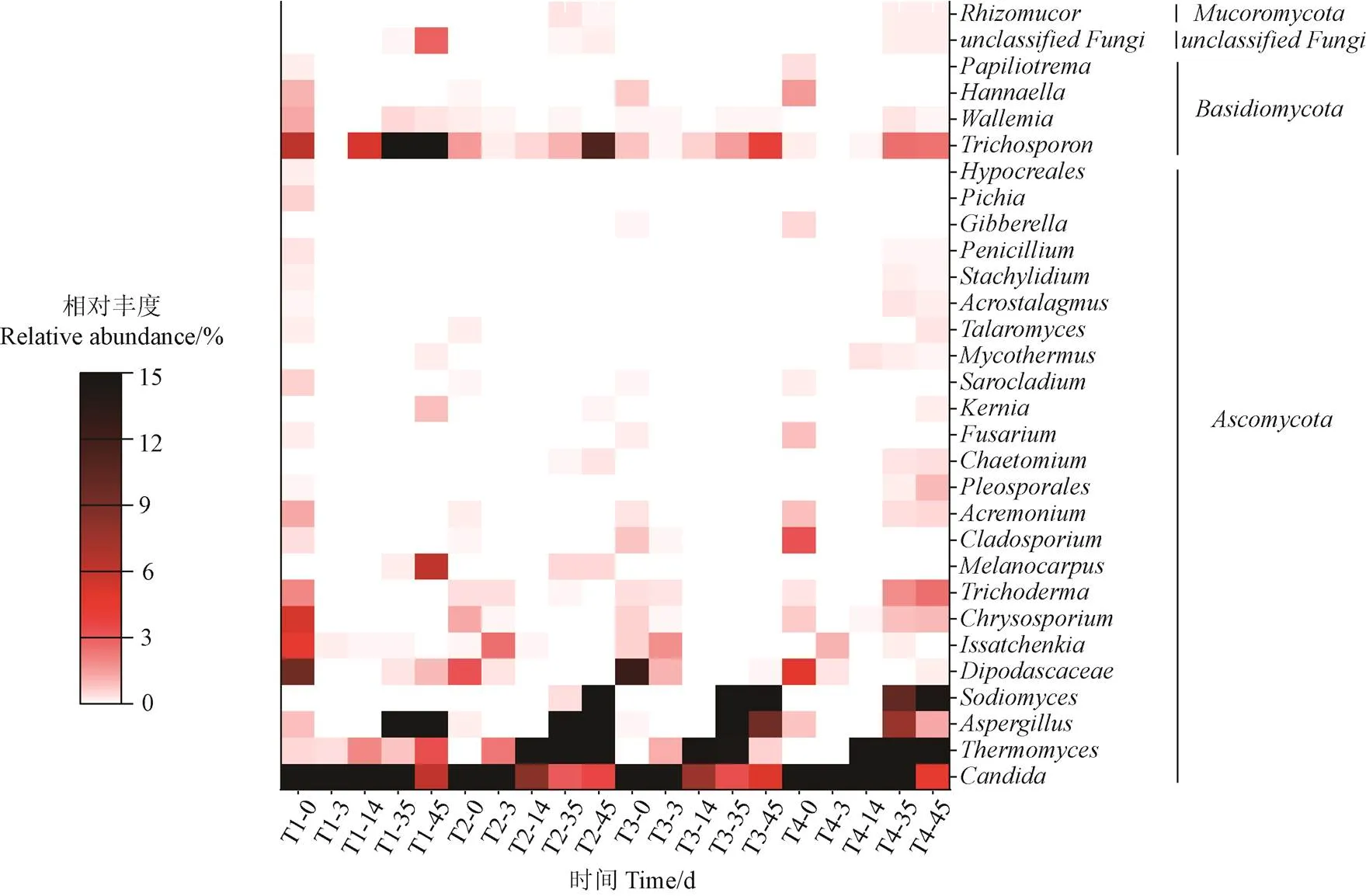
图9 堆肥过程中真菌群落在属水平(前30)上的相对丰度
2.4.3 微生物群落与基本腐熟度指标、有机质、HS组分之间的相关性
冗余分析(Redundancy Analysis,RDA)可进一步确定堆肥过程中微生物群落与基本腐熟度指标、有机质、HS组分之间的相关性。如图10所示,RDA的两个轴共解释了79.4%的细菌群落变化。其中,温度、总糖、HS、pH、EC与细菌群落显著相关(<0.01)。总糖解释了56.4%的细菌群落变化,占比最大。、地芽孢杆菌属()、杆菌属()等细菌与总糖负相关,与HS正相关,说明在缺乏多酚的情况下,这些微生物可以通过降解总糖产生还原糖,并与氨基酸合成HS。Rastogi等[49]指出在城市固体废弃物堆肥中接种能够避免酸化,获得更高HA含量的优质堆肥。温度解释了14.7%的细菌群落变化,嗜热新芽孢杆菌属()、、等细菌与温度存在正相关,说明这些菌属能够抵御高温,在堆肥过程发挥着重要作用[50]。片球菌属()和与pH值存在负相关,说明了和可在酸性条件下生存,即在堆肥初期活性较高(图8)。Tran等[51]在餐厨垃圾堆肥中也发现,这两种微生物可以加速堆肥过程,产生乳酸,促进土著微生物的生长繁殖。研究表明,细菌活性会受理化性质的影响,进而影响HS前驱物的产生和HS形成[32]。在本研究中,、与EC存在负相关,且与总糖、木质纤维素呈负相关关系,与HS、GI呈正相关关系,再次说明了盐含量越高,和的丰度越低,抑制了总糖、木质纤维素的降解,减少了HS前驱物的产生,不利于HS形成,降低堆肥腐殖化效果。
优势真菌属与基本腐熟度指标、有机质和HS组分之间的相关性分析表明(图11),RDA1和RDA2分别解释了38.5%和31.0%的真菌群落变化。其中,GI解释了34.3%的真菌群落差异(<0.01),根毛霉属()、嗜热链球菌属()、、等微生物与GI存在正相关。
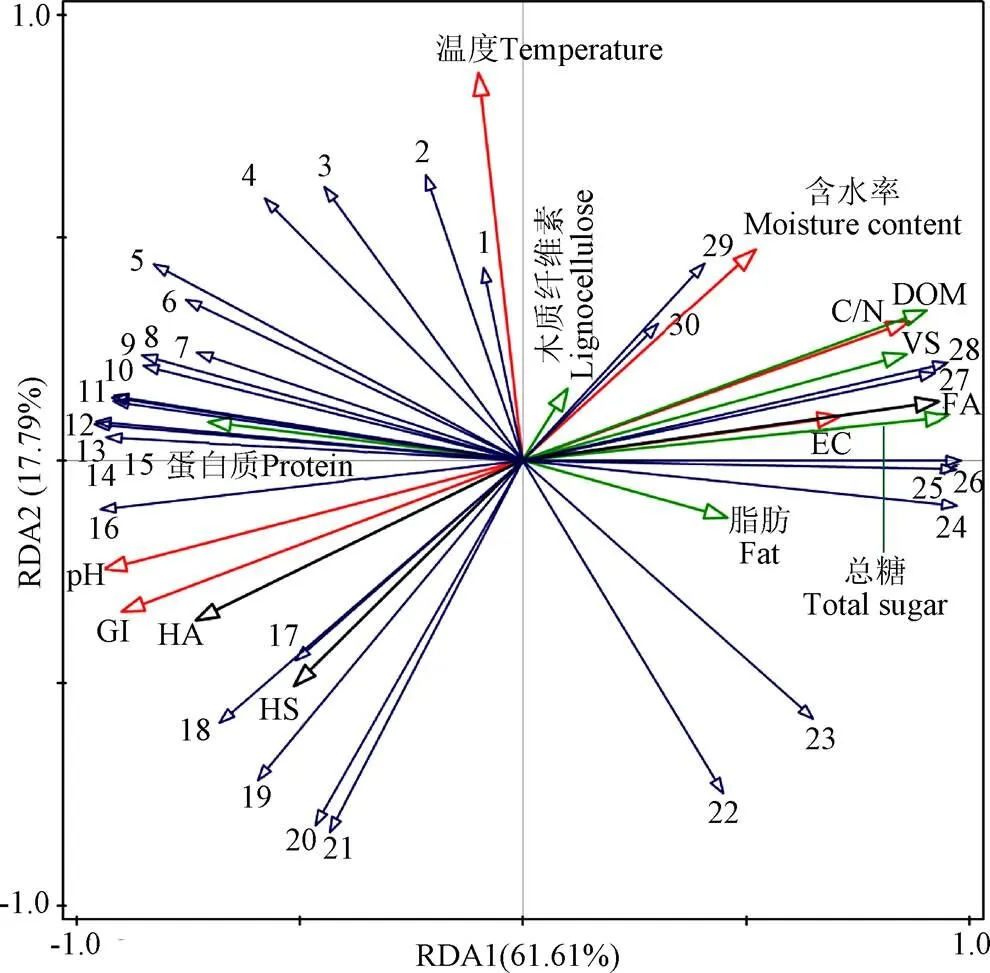
注:数字1~30分别代表Ornithinibacillus、Paenibacillus、Brevibacillus、Ureibacillus、Bacillaceae、Bacillus、Geobacillus、Planifilum、Saccharomonospora、Limnochordaceae、Bacilli、Thermobifida、unclassified Bacillaceae、Oceanobacillus、Novibacillus、Bacillales、Gracilibacillus、Fodinicurvataceae、Truepera、Georgenia、JG30-KF-CM45、Glycomyces、Nocardiopsis、Leuconostoc、Pediococcus、Lactobacillus、Weissella、Streptomyces、Lysinibacillus和Saccharopolyspora。
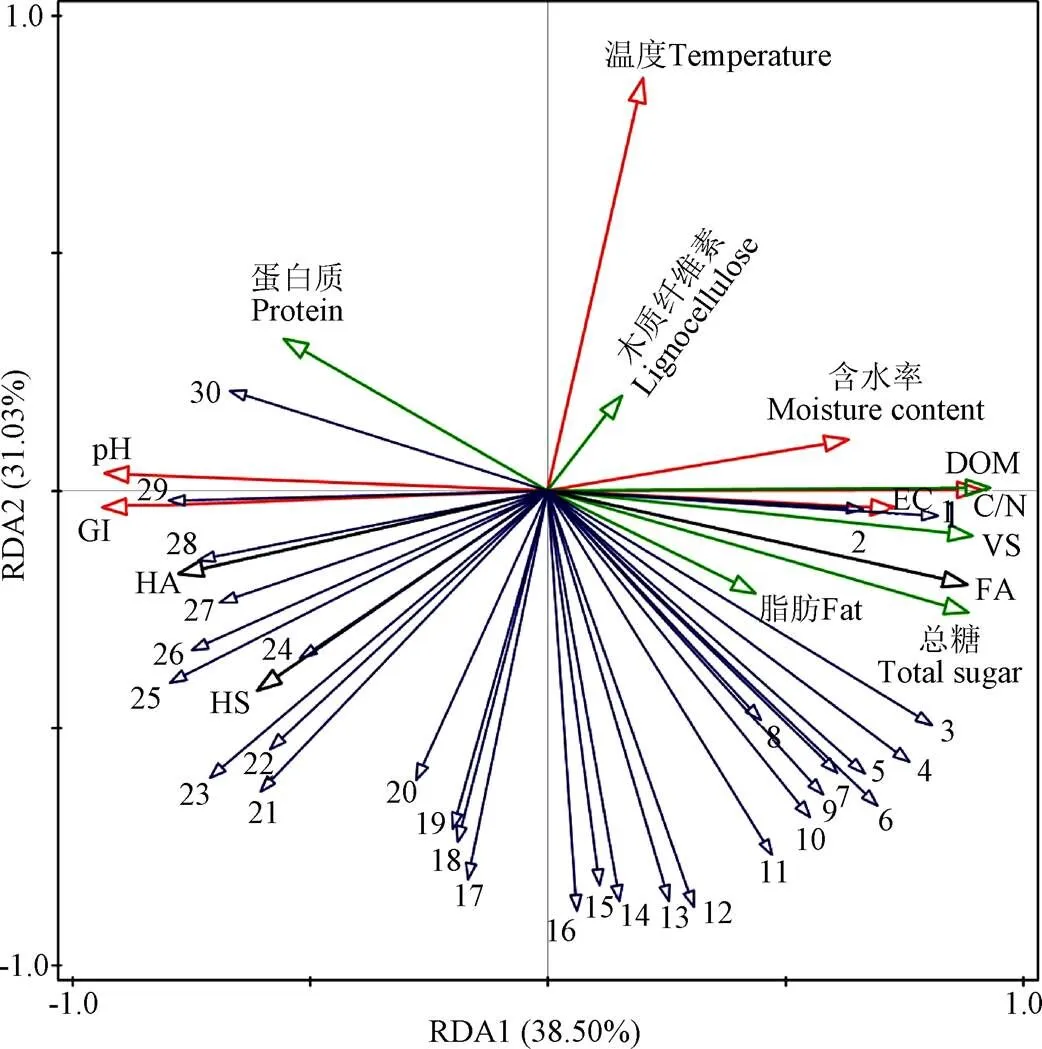
注:数字1 ~ 30分别代表Candida、Issatchenkia、Papiliotrema、Hannaella、Fusarium、Sarocladium、Cladosporium、Pichia、Gibberella、Dipodascaceae、Hypocreales、Chrysosporium、Acremonium、Talaromyces、Trichoderma、Penicillium、Stachylidium、Acrostalagmus、Wallemia、Pleosporales、Chaetomium、Kernia、Aspergillus、Trichosporon、unclassifiedFungi、Sodiomyces、Melanocarpus、Mycothermus、Rhizomucor和Thermomyces。
由此可以推断,在堆肥过程中接种这些微生物可提高GI值,获得较高的腐熟度。温度解释了25.9%的真菌群落差异(< 0.01)。唐聪[52]也指出温度显著影响真菌群落组成。、与温度存在正相关,说明了这两种微生物能够在高温环境下存活,与其丰度相对应(图9)。木霉属()与温度存在负相关,反映了在堆肥高温阶段其活性可能会受到抑制,这与Jiang等[53]在污水污泥堆肥中的研究结果一致。木质纤维素解释了7.0%的真菌群落差异(<0.01),节担菌属()、笋顶孢霉属()、指轮枝孢属()等微生物与木质纤维素存在负相关,与HS存在正相关,说明了这些微生物能够参与木质纤维素的降解,以各种形式聚合形成HS。值得注意的是,本研究中的、与EC、总糖、木质纤维素均呈负相关,与GI呈正相关,同样证实了高盐环境会抑制和的生长繁殖,减少总糖、木质纤维素等有机物腐殖化过程,影响HS产生。
3 结 论
1)餐厨垃圾与玉米秸秆联合堆肥过程,添加盐分由0.5%增加至1.5%时,高温期缩短至13 d,增加堆体电导率值,从而抑制种子发芽,种子发芽指数值降低至65.5%,不利于堆肥腐熟。
2)盐分增加会抑制蛋白质、总糖、木质纤维素和脂肪降解,限制有机质腐殖化过程,不利于腐殖质、胡敏酸的合成。光谱学分析表明,盐分含量增加会降低腐殖质的芳香化结构和稳定性。
3)高盐含量(添加1.5%)降低细菌属、和真菌属、的相对丰度,抑制总糖、木质纤维素等有机物腐殖化过程,影响后续腐殖质形成。
[1] 杨召峰,苑宏英,叶际亮,等. 剩余污泥和餐厨垃圾制取生物柴油现状及前景分析[J]. 天津城建大学学报,2021,27(6):431-435.
Yang Zhaofeng, Yuan Hongying, Ye Jiliang, et al. Analysis on the present situation and prospect of biodiesel production from waste activated sludge and kitchen waste[J]. Journal of Tianjin Chengjian University, 2021, 27(6): 431-435. (in Chinese with English abstract)
[2] 郝云东,周学锋. 农村生活垃圾分类处理问题及对策研究[J]. 农村经济与科技,2022,33(1):214-216.
Hao Yundong, Zhou Xuefeng. Study on problems and countermeasures of classified treatment of rural domestic waste[J]. Rural Economy and Science-Technology, 2022, 33(1): 214-216. (in Chinese with English abstract)
[3] Xu Z, Ma Y, Zhang L, et al. Relating bacterial dynamics and functions to gaseous emissions during composting of kitchen and garden wastes[J]. Science of The Total Environment, 2021, 767: 144210.
[4] 满吉勇,林永锋,李丛林,等. 餐厨垃圾堆肥对水稻生长、产量及土壤养分含量的影响[J]. 中国农业大学学报,2021,26(11):165-179.
Man Jiyong, Lin Yongfeng, Li Conglin, et al. Effects of kitchen waste compost on the yield and quality of rice and soil nutrient content[J]. Journal of China Agricultural University, 2021, 26(11): 165-179. (in Chinese with English abstract)
[5] 聂永丰. 固体废物处理工程技术手册[M]. 北京:化学工业出版社,2013.
[6] 朱慧. 餐厨垃圾及组分对几种常见微生物生长的影响[D]. 南京:南京大学,2019.
Zhu Hui. The Effect of Kitchen Waste and Its Components on the Growth of Several Common Microbes[D]. Nanjing: Nanjing University, 2019.(in Chinese with English abstract)
[7] Zhao J, Zhang C, Wang D, et al. Revealing the underlying mechanisms of how sodium chloride affects short-chain fatty acid production from the co-fermentation of waste activated sludge and food waste[J]. Acs Sustainable Chemistry and Engineering, 2016: 6b-816b.
[8] Wang P, Wang H T, Qiu Y Q, et al. Microbial characteristics in anaerobic digestion process of food waste for methane production: A review[J]. Bioresource Technology, 2018, 248: 29-36.
[9] 任连海,黄燕冰,王攀. 含盐量对餐厨垃圾堆肥理化特性变化规律的影响[J]. 重庆大学学报,2014,37(7):104-109.
Ren Lianhai, Huang Yanbing, Wang Pan. Effect of salt content on the change of physicochemical characteristics of restaurant garbage during aerobic composting[J]. Journal of Chongqing University, 2014, 37(7): 104-109. (in Chinese with English abstract)
[10] Wu Z, Yin B, Song X, et al. Effects of salinity on earthworms and the product during vermicomposting of kitchen wastes[J]. International Journal of Environmental Research and Public Health, 2019, 16(23): 4737.
[11] 于兆友,闫海冰,张慧芳,等. 不同盐分胁迫对皂荚种子萌发及幼苗生理特征的影响[J]. 东北农业大学学报,2020,51(10):28-35.
Yu Zhaoyou, Yan Haibing, Zhang Huifang, et al. Effect of different salt stresses on seed germination and seedling physiological characteristics of Gleditsia sinensis[J]. Journal of Northeast Agricultural University, 2020, 51(10): 28-35. (in Chinese with English abstract)
[12] 周海宾, 刘娟, 丁京涛, 等. 采用EEM-FRI方法研究黑曲霉对牛粪堆肥腐熟及纤维素降解影响[J]. 农业工程学报, 2022,38(1):276-286.
Zhou Haibin, Liu Juan, Ding Jingtao, et al. Effects ofon maturity and cellulose degradation of cow manure composting using EEM-FRI method[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2022, 38(1): 276-286.(in Chinese with English abstract)
[13] 迟凯威. 盐碱胁迫下玉米秸秆堆肥对玉米生长发育的影响及转录组分析[D]. 哈尔滨:东北林业大学2021.
Chen Kaiwei. Effects of Maize Straw Composting on Maize Growth and Development and Transcriptome Analysis Under Saline-alkali Stress[D]. Harbin: Northeast Forestry University, 2021. (in Chinese with English abstract)
[14] Van Soest P J, Robertson J B, Lewis B A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition[J]. Journal of Dairy Science, 1991, 74(10): 3583-3597.
[15] 蒋志伟. 赤泥对堆肥中木质纤维素降解和腐殖质形成及功能微生物群落演替的影响[D]. 南宁:广西大学,2020.
Jiang Zhiwei. Effects of Red Mud on the Degradation of Lignocellulose and Humus Formation and Succession of Functional Microbial Communities During Composting[D]. Nanning: Guangxi University, 2020. (in Chinese with English abstract)
[16] 佟影影. 畜禽粪便堆肥过程溶解性有机质组成与重金属变化研究[D]. 合肥:安徽农业大学, 2020.
Tong Yingying. Study on the Changes of Dissolved Organic Matter and Heavy Metals in Compost[D]. Hefei: Anhui Agricultural University, 2020.(in Chinese with English abstract)
[17] 欧阳兆威. 高盐高油废水处理新工艺技术研究[D]. 长沙:湖南农业大学,2018.
Ouyang Zhaowei. Study on New Process Technology of High Concentration Salt and Oil Wastewater Treatment[D]. Changsha: Hunan Agricultural University, 2018.(in Chinese with English abstract)
[18] Yang F, Li G, Yang Q, et al. Effect of bulking agents on maturity and gaseous emissions during kitchen waste composting[J]. Chemosphere, 2013, 93(7): 1393-1399.
[19] Xu Z, Xu W, Zhang L, et al. Bacterial dynamics and functions driven by bulking agents to mitigate gaseous emissions in kitchen waste composting[J]. Bioresource Technology, 2021, 332: 125028.
[20] Jiang J, Wang Y, Yu D, et al. Combined addition of biochar and garbage enzyme improving the humification and succession of fungal community during sewage sludge composting[J]. Bioresource Technology, 2021: 126344.
[21] Li X, Wang P, Chu S, et al. Short-term biodrying achieves compost maturity and significantly reduces antibiotic resistance genes during semi-continuous food waste composting inoculated with mature compost[J]. Journal of Hazardous Materials, 2021: 127915.
[22] Meng X, Yan J, Zuo B, et al. Full-scale of composting process of biogas residues from corn stover anaerobic digestion: Physical-chemical, biology parameters and maturity indexes during whole process[J]. Bioresource Technology, 2020, 302: 122742.
[23] Chandna P, Nain L, Singh S, et al. Assessment of bacterial diversity during composting of agricultural byproducts[J]. BMC Microbiology, 2013, 13(1): 99.
[24] Gao X, Xu Z, Li Y, et al. Bacterial dynamics for gaseous emission and humification in bio-augmented composting of kitchen waste[J]. Science of The Total Environment, 2021, 801: 149640.
[25] Mavi M S, Marschner P, Chittleborough D J, et al. Salinity and sodicity affect soil respiration and dissolved organic matter dynamics differentially in soils varying in texture[J]. Soil Biology and Biochemistry, 2012, 45: 8-13.
[26] Wei Y, Wu D, Wei D, et al. Improved lignocellulose-degrading performance during straw composting from diverse sources with actinomycetes inoculation by regulating the key enzyme activities[J]. Bioresource Technology, 2019, 271: 66-74.
[27] Liu L, Ye Q, Wu Q, et al. Effect of biochar addition on sludge aerobic composting and greenbelt utilization[J]. Environmental Technology & Innovation, 2021, 21: 101279.
[28] Wu J, Qi H, Huang X, et al. How does manganese dioxide affect humus formation during bio-composting of chicken manure and corn straw?[J]. Bioresource Technology, 2018, 269: 169-178.
[29] Barje F, El-Fels L, El-Hajjouji H, et al. Molecular behaviour of humic acid-like substances during co-composting of olive mill waste and the organic part of municipal solid waste[J]. International Biodeterioration & Biodegradation, 2012, 74: 17-23.
[30] Zhang Z, Zhao Y, Yang T, et al. Effects of exogenous protein-like precursors on humification process during lignocellulose-like biomass composting: Amino acids as the key linker to promote humification process[J]. Bioresource Technology, 2019, 291: 121882.
[31] Dias Bruno O, Silva Carlos A, Higashikawa Fábio S, et al. Use of biochar as bulking agent for the composting of poultry manure: Effect on organic matter degradation and humification[J]. Bioresource Technology, 2010, 101(4): 1239-1246.
[32] Wu J, Zhao Y, Zhao W, et al. Effect of precursors combined with bacteria communities on the formation of humic substances during different materials composting[J]. Bioresource Technology, 2017, 226: 191-199.
[33] Zhu N, Gao J, Liang D, et al. Thermal pretreatment enhances the degradation and humification of lignocellulose by stimulating thermophilic bacteria during dairy manure composting[J]. Bioresource Technology, 2021, 319: 124149.
[34] Haddad G, El-Ali F, Mouneimne A H. Humic matter of compost: Determination of humic spectroscopic ratio (E4/E6)[J]. Current Science International, 2015, 4(1): 56-72.
[35] 崔东宇,何小松,席北斗,等. 牛粪堆肥过程中水溶性有机物演化的光谱学研究[J]. 中国环境科学,2014,34(11):2897-2904.
Cui Dongyu, He Xiaosong, Xi Beidou, et al. Spectrum analysis on the evolution of dissolved organic matter during cattle manure composting[J]. China Environmental Science, 2014, 34(11): 2897-2904.(in Chinese with English abstract)
[36] Chen J, Leboeuf E J, Dai S, et al. Fluorescence spectroscopic studies of natural organic matter fractions[J]. Chemosphere, 2003, 50(5): 639-647.
[37] Li R, Meng H, Zhao L, et al. Study of the morphological changes of copper and zinc during pig manure composting with addition of biochar and a microbial agent[J]. Bioresource Technology, 2019, 291: 121752.
[38] Zhang T, Wu X, Shaheen Sabry M, et al. Effects of microorganism-mediated inoculants on humification processes and phosphorus dynamics during the aerobic composting of swine manure[J]. Journal of Hazardous Materials, 2021, 416: 125738.
[39] Antunes Luciana P, Martins Layla F, Pereira Roberta V, et al. Microbial community structure and dynamics in thermophilic composting viewed through metagenomics and metatranscriptomics[J]. Scientific Reports, 2016, 6(1): 38915.
[40] Li Y, Liu Y, Yong X, et al. Odor emission and microbial community succession during biogas residue composting covered with a molecular membrane[J]. Bioresource Technology, 2019, 297: 122518.
[41] Ge M, Shen Y, Ding J, et al. New insight into the impact of moisture content and pH on dissolved organic matter and microbial dynamics during cattle manure composting[J]. Bioresource Technology, 2022, 344: 126236.
[42] Klenk H P, Held B, Lucas S, et al. Genome sequence of the soil bacteriumtype strain (NA-128(T))[J]. Standards in Genomic Sciences, 2012, 6(2): 220-229.
[43] Zhang L, Zhang H, Wang Z, et al. Dynamic changes of the dominant functioning microbial community in the compost of a 90 m3aerobic solid state fermentor revealed by integrated meta-omics[J]. Bioresource Technology, 2016, 203: 1-10.
[44] Liu C, Liu J, Li J, et al. Removal of H2S by co-immobilized bacteria and fungi biocatalysts in a bio-trickling filter[J]. Process Safety and Environmental Protection, 2013, 91(1): 145-152.
[45] 王秀红,史向远,张纪涛,等. 鸡粪好氧堆肥腐熟度、重金属残留及微生物菌群分析[J]. 山西农业科学,2021,49(9):6.
Wang Xiuhong, Shi Xiangyuan, Zhang Jitao, et al. Analysis of maturity, heavy metal residues and microbial flora of chicken manure aerobic compost[J]. Journal of Shanxi Agricultural Sciences, 2021, 49(9): 6. (in Chinese with English abstract)
[46] 唐小飞. 西藏热泉嗜热真菌分离鉴定、产酶及生物活性初步研究[D]. 汉中:陕西理工大学,2019.
Tang Xiaofei. Preliminary Study on Production Enzyme and Biological Activity Form Isolation and Identification of Thermophilic Fungi in Tibetan Hot Spring[D]. Hanzhong: Shanxi University of Technology, 2019. (in Chinese with English abstract)
[47] Awasthi M K, Pandey A K, Bundela P S, et al. Co-composting of organic fraction of municipal solid waste mixed with different bulking waste: Characterization of physicochemical parameters and microbial enzymatic dynamic[J]. Bioresource Technology, 2015, 182: 200-207.
[48] Wang K, Mao H, Li X. Functional characteristics and influence factors of microbial community in sewage sludge composting with inorganic bulking agent[J]. Bioresource Technology, 2018, 249: 527-535.
[49] Rastogi M, Nandal M, Khosla B. Microbes as vital additives for solid waste composting[J]. Heliyon, 2020, 6(2): e3343.
[50] Wang J, Liu Z, Xia J, et al. Effect of microbial inoculation on physicochemical properties and bacterial community structure of citrus peel composting[J]. Bioresource Technology, 2019, 291(C): 121843.
[51] Tran Q M, Mimoto H, Koyama M, et al. Lactic acid bacteria modulate organic acid production during early stages of food waste composting[J]. Science of The Total Environment, 2019, 687: 341-347.
[52] 唐聪. 农业废物好氧堆肥中不同位置微生物群落分布特征的驱动机制响应研究[D]. 长沙:湖南大学,2013.
Tang Cong. The Driving Mechanism of the Spatial Distribution of Bacterial and Fungal Communities within a Pile during Agricultural Waste Composting[D]. Changsha: Hunan University, 2013.(in Chinese with English abstract)
[53] Jiang J, Pan Y, Yang X, et al. Beneficial influences of pelelith and dicyandiamide on gaseous emissions and the fungal community during sewage sludge composting[J]. Environmental Science and Pollution Research, 2019, 26(9): 8928-8938.
Effects of salt content on the humification and microbial community succession of food waste aerobic composting
Liu Juan1,2,4, Shen Yujun1,2,4, Luo Wenhai2, Ding Jingtao1,2,4, Meng Haibo1,4, Zhou Haibin1,2,3,4※, Zhang Xi1,2,4, Cheng Hongsheng1,4, Wang Jian1,4, Xu Pengxiang1,4
(1100125,; 2,100193,; 3.,,832000,;4.,,100125
Food waste is ever increasing in rural areas in recent years, particularly with the rapid economic development and population growth. If disposed of improperly, severe pollution can pose a great threat to the rural living environment. Fortunately, aerobic composting has been widely applied for food waste treatment, due to the high harmlessness and resource utilization rate. It is worth noting that food waste with high salinity can limit the application of aerobic composting. Among them, humification is one of the most important processes in the application of compost. However, only a few studies focused on the effect of salt content on aerobic compost humification. It is still lacking in the influencing mechanisms from the perspective of microbial community structure evolution. Taking food waste as the main raw material, and the maize stover as an auxiliary material, this study aims to explore the influences of salt content on the basic maturity index, organic matter composition, and dynamic change of humus substances during the composting process. The succession of microbial community was combined with the correlation statistical analysis, in order to reveal the microbial action about the effect of salt content on the mature process of food waste aerobic composting. Specifically, the food waste and maize stover were mixed well at the ratio of 4:1, where the initial moisture content was adjusted to about 63%. A composting experiment was carried out in the 60 L composting reactors for 45 days. A total of four experimental treatments were set with the addition of 0%, 0.5%, 1.0%, and 1.5% salt. Among them, the treatment without salt was the control group. The results showed that the temperature, C/N, and organic matter composition of four treatments fully met the requirements of maturity during aerobic composting. However, the salt addition greatly increased the electrical conductivity of the reactor. Particularly, the thermophilic period was shortened to 13 days at the proportion of salt addition of 1.5%. But, the seed growth was inhibited with the seed germination index reducing to 65.5%. Compared with the control treatment without salt, the degradation rate of organic matter was reduced by 6.5%, indicating the restricted humification process. The high throughput sequencing showed that the addition of 1.5% salt inhibited the activities of Thermobifida, Saccharomonospora, Aspergillus, and Trichosporon. In addition, a correlation analysis showed that the high salinity reduced the biodegradation of organic matter, such as total sugar and lignocellulose, thereby limiting the subsequent formation of humus substances. Thus, restricted improvement was found in the humification effect during food waste and maize stover composting. These findings can provide theoretical support for salt regulation in the application of aerobic composting in food wastes.
microorganism; aerobic composting; humification; food waste; salt
10.11975/j.issn.1002-6819.2022.19.021
S141.4
A
1002-6819(2022)-19-0190-12
刘娟,沈玉君,罗文海,等. 盐含量对餐厨垃圾好氧堆肥腐殖化过程及微生物演变的影响[J]. 农业工程学报,2022,38(19):190-201.doi:10.11975/j.issn.1002-6819.2022.19.021 http://www.tcsae.org
Liu Juan, Shen Yujun, Luo Wenhai, et al. Effects of salt content on the humification and microbial community succession of food waste aerobic composting[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2022, 38(19): 190-201. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2022.19.021 http://www.tcsae.org
2021-10-01
2022-05-13
现代农业产业技术体系北京市创新团队项目(BAIC08-2022-FQ03);北京市科技新星计划项目(Z191100001119063)
刘娟,研究方向为废弃物处理与资源化。Email:2528506831@qq.com。
周海宾,高级工程师,博士,研究方向为农业农村废弃物资源化利用。Email:nxzhb66@163.com
