甜菜幼苗光合生理对干旱胁迫的响应
2022-02-03尹希龙石杨李王胜兴旺
尹希龙 石杨 李王胜 兴旺
(1黑龙江大学国家甜菜种质中期库,150080,黑龙江哈尔滨;2黑龙江大学现代农业与生态环境学院,150080,黑龙江哈尔滨)
甜菜(Beta vulgaris L.)是苋科甜菜属2年生草本植物,为我国重要的经济作物和糖料作物之一[1]。华北、东北和西北地区是甜菜种植的优势产区,甜菜主要种植在新疆、黑龙江、内蒙古等地[2-3],这些地区幅员辽阔,昼夜温差大,适合甜菜生长及糖分积累[4]。但是这些地区多处于干旱、半干旱地区,土壤对水资源的承载能力较低,环境脆弱[5]。甜菜在生长时需要大量水分,特别是在苗期对水分亏缺十分敏感[6]。干旱胁迫不仅影响甜菜正常的生长发育,而且会造成产量和品质的损失,成为限制甜菜产量的重要因素[7]。如何培育出更加耐旱的甜菜种质资源,以便更好地在干旱、半干旱区扩大甜菜种植及保持甜菜稳产已成为重要研究内容[8]。
植物在生长过程中会遇到各种非生物胁迫,如高温[9]、冷害[10]和干旱等[11]。水分是植物生长的重要条件,在作物生长过程中如果水分亏缺,最先响应的是根系[12]。植物根系伸长受到抑制,吸收水分受阻,会造成作物生长缓慢。植物的叶片状态直接反映出植株受到干旱胁迫的影响[13],干旱胁迫不仅会影响叶绿素含量,还会影响光合作用[14]。干旱胁迫降低植物叶片气孔导度(Gs),抑制对CO2的吸收,影响净光合速率(Pn)和蒸腾速率(Tr)[15]。水是光合作用的原料,光合作用是植物生长发育的重要代谢过程,是植物必不可少的生命活动[16]。在甜菜的非生物胁迫研究方面,大多集中在干旱胁迫对甜菜的生长和生理生化影响的研究,对干旱胁迫下甜菜表观光合指标的研究有一些报道[17-18],研究了甜菜在干旱胁迫下根膨大期光合光响应特性和干旱胁迫对甜菜叶丛期光合作用的影响等。
本试验研究干旱胁迫下耐旱型品种甜菜和干旱敏感型甜菜生长发育和光合生理变化,有助于从生理层面探讨不同甜菜种质对于干旱胁迫响应的差异,探究不同耐旱性甜菜种质对干旱胁迫的响应。丰富甜菜抗旱机制方面研究内容,为甜菜抗旱育种及稳产提供理论依据。
1 材料与方法
1.1 试验材料
试验在黑龙江大学国家甜菜种质中期库进行。供试材料为耐旱甜菜品种依安一号(V1)和干旱敏感甜菜品种92011/1-6/1(V2)。二者均由国家甜菜种质中期库(国家甜菜种质资源平台)提供。
1.2 试验设计
将甜菜种子用75%酒精消毒30s,冲洗干净,在2‰的福美双液体中浸泡12h。蛭石提前烘干,在出苗盆(高25.00cm,宽20.00cm,长35.00cm)中放入适量蛭石,将甜菜种子均匀撒在蛭石中,保持种子萌发过程中水分充足。人工气候室培养10d后,待甜菜长出2片子叶,移入Hoagland营养液中培养。人工气候室培养条件为320μmol/(m2·s),温度27℃,光/暗周期14h/10h[19]。培养至第3对真叶完全展开,选取长势一致的甜菜幼苗,进行干旱胁迫。干旱胁迫采用PEG-6000溶液(浓度为9%)模拟(DS),正常生长条件为对照组(CK),每个试验时间点取样设置3次重复,即每次取长势一致的3株甜菜幼苗。干旱胁迫第2、5、7天,测定甜菜幼苗生长指标、叶片总叶绿素含量和表观光合指标。
1.3 测定项目与方法
茎粗:采用S071012数显游标卡尺(杭州巨星钢盾工具有限公司)测定茎粗。
株高:用直尺测定甜菜幼苗地上部自然高度。
根长:用直尺测定甜菜幼苗地下部自然长度。
叶鲜重:采用ALC210.2电子天平(北京赛多利斯仪器系统有限公司)测定去除叶柄的甜菜幼苗叶片质量。
根鲜重:用电子天平测定甜菜幼苗根质量。
叶干重:用电子天平测定在OGS干燥箱(赛默飞世尔科技公司)烘干的去除叶柄的甜菜幼苗叶片质量。
根干重:用电子天平测定在干燥箱烘干的甜菜幼苗根质量。
甜菜幼苗总叶绿素含量:称取不同处理的甜菜幼苗叶片0.50g,充分研磨,用乙醇提取,离心,取上清液,用比色法测定[20]。
甜菜幼苗表观光合指标:采用CI-340光合仪(思爱迪生态科学仪器有限公司)在每天上午9:00-11:00进行测定。测定部位为甜菜幼苗第2对完全展开的真叶,每个种质不同处理均测定3株,每株测3次。
1.4 数据处理
采用Microsoft Excel 2019和Origin 2022等统计软件进行相关数据分析统计及作图,采用DPS 18.0软件进行二因素有重复试验统计分析,差异显著性定义为P<0.05,所有数据均为3次重复的平均值±标准差[21]。
2 结果与分析
2.1 干旱胁迫对甜菜幼苗生长的影响
从表1可知,在正常生长条件下,品种V1和V2幼苗的茎粗、株高、根长、叶鲜重、根鲜重、叶干重和根干重随着生长时间的增加而增加。

表1 甜菜幼苗表型指标Table 1 Phenotypic indexes of sugar beet seedlings
在干旱胁迫下,2个甜菜品种幼苗的生长明显受到抑制,随着时间的增加,抑制程度不断增强。V1和V2幼苗诸多生长指标呈下降趋势。其中,在第7天,甜菜幼苗受到干旱胁迫影响最严重。第7天的干旱胁迫使V1和V2幼苗的株高分别下降51.38%和74.65%(DS与CK比较,下同);V1和V2幼苗的根长分别下降到20.33和13.00cm;V1和V2幼苗的叶鲜重下降幅度分别是76.06%和84.89%,V2下降幅度比V1大。干旱胁迫对甜菜幼苗的生长影响较大,干旱胁迫下V1与V2生长指标比较,进一步说明V1的耐旱性比V2好。
2.2 干旱胁迫对甜菜幼苗总叶绿素含量的影响
由图1可知,在正常生长条件下,V1和V2幼苗叶片的叶绿素含量随时间增加逐渐增加。在第2、5、7天,V1和V2幼苗叶片的叶绿素含量没有显著差异。
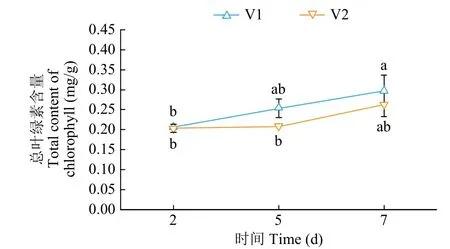
图1 正常水分条件下甜菜幼苗叶片总叶绿素含量Fig.1 Total chlorophyll content of sugar beet seedling leaves under normal water conditions
由图2可知,在干旱胁迫条件下,V1和V2幼苗叶片的叶绿素含量显著降低。V1幼苗的叶绿素含量降低幅度小于V2。

图2 干旱胁迫下甜菜幼苗叶片总叶绿素含量Fig.2 Total chlorophyll content of sugar beet seedling leaves under drought stress
由图3可知,在第2天V1幼苗叶片的叶绿素含量没有显著差异;在第5天,不同处理间V1幼苗叶片的叶绿素含量有显著差异,CK处理叶绿素含量为0.25mg/g,DS处理叶绿素含量为0.18mg/g;在第7天,不同处理间V1幼苗叶片的叶绿素含量有显著差异,含量显著降低,降幅为48.01%。

图3 不同水分条件下V1甜菜幼苗叶片总叶绿素含量Fig.3 Total chlorophyll content of V1 sugar beet seedling leaves under different water conditions
由图4可知,在第2天,不同处理间V2幼苗叶片的叶绿素含量没有显著差异;在第5天,DS处理的V2幼苗叶片叶绿素含量开始显著降低,具有显著差异,下降幅度为30.58%;在第7天,不同处理间V2幼苗叶片的叶绿素含量有显著差异,CK处理叶绿素含量为0.26mg/g,DS处理叶绿素含量为0.11mg/g,下降幅度为58.05%。

图4 不同水分条件下V2甜菜幼苗叶片总叶绿素含量Fig.4 Total chlorophyll content of V2 sugar beet seedling leaves under different water conditions
2.3 干旱胁迫对甜菜幼苗表观光合指标的影响
从表2可知,在正常生长条件下,V1和V2幼苗叶片的Pn是随时间增加呈上升趋势。在干旱胁迫下,V1和V2幼苗叶片的Pn均显著低于对照组。干旱胁迫下,甜菜幼苗叶片的Pn呈下降趋势。其中V2幼苗叶片的Pn受到干旱胁迫的影响比V1大,在第7天时降到最低,达到了 0.20μmol/(m²·s),比对照组降低了96.54%。
在未受到干旱胁迫情况下,V1幼苗叶片的Tr逐渐增加,V2幼苗叶片的Tr变化规律和V1相似。干旱胁迫下,V1和V2幼苗叶片的Tr随着时间的增加逐渐下降,V1幼苗叶片的Tr在第5天时显著下降,而V2幼苗叶片的Tr在第2天时显著下降。V1和V2幼苗叶片的Tr在第7天时降至最低值,下降幅度分别是77.73%、89.53%,下降幅度较大的是V2,V1表现出较强的耐旱性。
由表2可知,在没有受到干旱胁迫情况下,V1和V2幼苗叶片Gs是逐渐增加的。在受到干旱胁迫时,与正常生长条件比较,V2幼苗叶片的叶片Gs逐渐下降,而V1幼苗叶片的Gs随时间呈先上升后下降的趋势。第2天时,V1幼苗叶片的Gs从CK处理的70.94mmol/(m²·s)升高到DS处理的111.60mmol/(m²·s),在第5、7天时,甜菜幼苗叶片Gs开始显著下降。而V2甜菜幼苗叶片的Gs在第2天时就开始显著下降,虽然V2的Gs比V1的要低,但是在第5或7天时,V1与V2没有显著差异。

表2 甜菜幼苗叶片表观光合指标Table 2 Apparent photosynthetic index of sugar beet seedling leaves
在正常水分条件下,V1和V2幼苗叶片的胞间CO2浓度(Ci)随干旱胁迫时间的增加呈上升趋势。干旱胁迫下,V1和V2幼苗叶片的Ci随时间的增加逐渐下降,V1幼苗叶片的Ci显著高于V2。V1幼苗叶片的Ci下降幅度分别是48.89%、54.90%和66.56%;V2幼苗叶片的Ci下降幅度分别是50.14%、69.94%和84.49%。V1幼苗叶片的Ci下降幅度比V2的小,受到干旱胁迫影响较小。
3 讨论
3.1 甜菜幼苗生长对干旱胁迫的响应
甜菜苗期耐旱性较弱,由于幼苗没有发育成完善的根系,对水分亏缺较为敏感[19]。干旱胁迫下,植物缺水而细胞水势降低,细胞的收缩和细胞壁的收缩不能同步,导致细胞原生质体破损,叶片出现卷曲或者萎蔫。耐旱性较强的植物根系发达,根冠比大,有较多的根毛数量,能够在干旱胁迫下保持根系的功能,进行正常水分吸收。本研究结果表明,干旱胁迫影响了甜菜幼苗的生长,包括抑制甜菜幼苗的根长、茎粗、株高的生长和降低甜菜幼苗叶鲜重、根鲜重、叶干重及根干重。但是耐旱型甜菜幼苗的各项生长指标均高于干旱敏感型甜菜幼苗,耐旱型甜菜幼苗的降幅比干旱敏感型甜菜降幅小。可能是耐旱型品种根系较为发达,根毛长度和数量高于敏感型,虽然根系长度没有增加,但能维持细胞水分的平衡,进行正常的水分吸收。而敏感型品种根系较弱,在干旱胁迫下不能伸长根系或者生成更多数量的根毛,体内的细胞水势遭到破坏,机体受损,根系结构功能遭到破坏,地上部分表现出萎蔫。随着干旱胁迫时间的增加,2种甜菜幼苗吸收水分能力均逐渐下降,敏感型品种叶片逐渐变黄,出现干枯斑点,但没有出现死亡,而耐旱型品种很少出现这种情况,进一步说明耐旱型品种甜菜和干旱敏感型甜菜生长指标对干旱胁迫的响应。王新军等[22]研究发现,番茄幼苗生长在干旱胁迫下无法吸收到充足的水分,植物器官无法生长发育,致使干物质合成下降。说明干旱胁迫下不仅根系出现响应,植物的器官发育也出现响应。2种甜菜幼苗器官生长发育不能得到充足的水分,幼苗鲜重和干重下降。2种甜菜幼苗的鲜重与干物质量随着时间的增加均呈下降趋势,耐旱型品种干旱胁迫下器官的生长发育虽然接近正常代谢水平,但代谢合成还是会减少,鲜重和干物质的重量也自然下降,而敏感型品种对水分变化敏感,器官功能不能正常运行,代谢合成能力急剧下降,鲜重和干物质重量显著降低。干旱胁迫下,甜菜幼苗的根系首先做出响应[23],甜菜幼苗失去的水分超过了根系的吸收水分,水分平衡受到破坏。甜菜幼苗器官发育由于水分的减少,物质合成减少,鲜重减少,干物质量也趋于下降[24]。
3.2 甜菜幼苗总叶绿素含量对干旱胁迫的响应
植物叶片中的叶绿素含量和植物的光合作用强弱有关,干旱胁迫下叶绿素合成减少[25]。干旱胁迫下气孔关闭,CO2吸收下降。叶绿体被膜和基质片层在干旱胁迫下皱缩,组织破损,光系统活力下降,电子传递和光合磷酸化活动降低。本研究结果显示,2种甜菜幼苗的叶绿素含量随干旱胁迫时间的增加,整体呈下降趋势,干旱敏感型甜菜幼苗的降幅比耐旱型甜菜幼苗的降幅大。说明在干旱胁迫下,叶绿体片层结构遭到破坏,吸收光能下降,光合作用下降,总叶绿素合成降低。李剑威等[26]研究显示,薄壳核桃幼苗在干旱胁迫下叶绿素含量下降。Wang等[27]研究表明,在干旱胁迫下,植物幼苗叶绿素含量显著下降,均低于对照组。前人研究结果与本研究结果相同。但曾继娟等[28]研究表明,胡枝子幼苗在干旱胁迫下,叶绿素的含量呈现先上升后降低的趋势。水分降低,植株的叶面积减小,叶绿素在一定程度上被压缩,先导致叶绿素含量升高,随着干旱胁迫程度的加深,叶片的水分缺失进一步加深,叶片功能遭到损害,叶绿素合成受限,叶绿素含量降低[29]。干旱胁迫下耐旱型品种并没有出现叶绿素含量先上升再降低的情况,可能是叶片面积没有减少,叶绿体结构破坏程度不深,组织结构较为完整,能够保持一定的光合作用,维持叶绿素的合成。敏感型品种在干旱胁迫下光系统活力快速下降,氧化程度增加,光合作用过程效率低下,总叶绿素含量迅速降低[30-31]。
3.3 甜菜幼苗表观光合对干旱胁迫的响应
水分的缺失会影响植物的光合生命活动,Pn、Tr、Gs和Ci常作为衡量植株光合作用的强弱[32-33]。本研究中,干旱胁迫下甜菜幼苗的Pn、Tr、Gs和Ci趋于下降。耐旱型品种甜菜幼苗的表观光合指标显著高于干旱敏感型种质,耐旱型甜菜幼苗Pn、Tr、Gs和Ci降低的幅度比干旱敏感型甜菜幼苗降低的幅度小。说明在干旱胁迫下甜菜幼苗水分缺失,光合作用下降,直接造成Pn降低[34]。Tr是植物在一定时间内单位叶面积蒸腾的水量,蒸腾作用反映出植株生命活动的强弱,随着干旱胁迫程度的增加,甜菜幼苗水分不足,生命活动强度降低[35]。气孔是水分和CO2进出的重要门户[36],在干旱胁迫下,甜菜幼苗叶片气孔保卫细胞和周围叶肉细胞水势不一致,导致Gs变化,叶片Gs张开变化和光反应一致[37]。植株体内CO2浓度不足以支持植物生命活动,甜菜幼苗需要从外界吸收CO2,而气孔的关闭会导致吸收CO2减少,Ci降低。陈芳等[38]研究显示,干旱胁迫下辣椒的Pn、Tr、Gs和Ci均降低。吴海霞等[39]研究结果显示,在干旱胁迫情况下,甜菜幼苗的气孔阻力不断增大,Gs随之变小。曹兵等[40]研究表明,随干旱胁迫的加剧,臭椿苗期Pn、Tr和Gs明显下降。前人研究结果与本研究结果一致。耐旱型品种生命活动较强,水势变化导致Gs和Ci减小;而敏感型品种生命活动较弱,光合作用效率下降,Gs迅速减小。
4 结论
干旱胁迫下,2种甜菜幼苗根系水分吸收减少,生长发育受到抑制,器官发育缓慢甚至破损,干物质合成下降。CK与DS处理比较,第7天时的V2和V1幼苗根长分别下降到13.00和20.33cm;V1和V2幼苗的叶鲜重下降幅度分别是45.05%和64.06%。叶绿体基质破坏,光合电子传递和氧化磷酸化活动降低,总叶绿素含量降低。随着干旱胁迫时间的增加,甜菜幼苗叶片Gs减小,吸收CO2能力下降,Tr和Pn降低。敏感型品种Pn在第7天时降到最低,为 0.20μmol/(m²·s),比对照组降低了96.54%。耐旱型品种甜菜幼苗的各项指标表现优于干旱敏感型种质,降幅总体上小于干旱敏感型种质,进一步说明2种甜菜种质耐旱性差异,能够更好地探究甜菜幼苗光合生理对干旱胁迫的响应。
为全面掌握不同耐旱型甜菜品种对干旱的应答机制,后续将从细胞学层面探讨甜菜不同种质对于干旱胁迫响应的差异及对相关作用的重要差异表达基因和代谢通路等进行转录组分析,丰富甜菜抗旱机制的研究内容,丰富甜菜幼苗对非生物胁迫响应的研究内容,为甜菜稳产及育种提供理论依据。
