嗜水气单胞菌外膜蛋白AHA1182 的原核表达、纯化与免疫原性研究
2022-02-02简思杰
简思杰,晁 嘉,孙 薇,陈 锐,丁 锐,陈 琛,5,刘 祥
(1. 陕西理工大学 生物科学与工程学院/中德天然产物研究所,陕西 汉中 723001;2. 阜阳师范大学 生物与食品工程学院,安徽 阜阳 236037;3. 陕南秦巴山区生物资源综合开发协同创新中心,陕西 汉中 723001;4. 陕西理工大学 秦巴生物资源与生态环境省部共建国家重点实验室(培育),陕西 汉中 723001;5. 陕西省资源生物重点实验室,陕西 汉中 723001)
嗜水气单胞菌(Aeromonas hydrophila)属弧菌科、气单胞菌属,革兰氏染色呈阴性,其广泛存在于淡水、土壤、污水以及人体粪便中,是一种广泛流行的“人-畜-水生动物”共患病病原菌[1]。我国是水产大国,2020 年,我国淡水产品总量3234.64 万t[2]。目前,嗜水气单胞菌感染的水生动物病例呈上升趋势,以淡水鱼类为主,其是鱼类的常见致病菌[3]。而且嗜水气单胞菌也是我国大规模流行淡水养殖鱼类暴发性疾病的主要病原菌[4]。嗜水气单胞菌已严重危害我国淡水养殖业,对此病原菌的防治急需加强。传统防治手段以抗生素防治及化学药品防治为主,但长期使用抗生素会造成严重后果。如滥用抗生素会导致病原菌耐药性增强,已经出现耐青霉素、四环素等菌株[5];抑制吞噬细胞的吞噬功能,导致鱼类自身免疫力低下;抗生素的滥用更在一定程度上破坏了生态平衡;也可随着食物链的富集对人类的健康产生危害。因此,亟须寻找生态友好型抗菌剂,其中基于外膜蛋白的亚单位疫苗是一种理想选择。
外膜蛋白(Outer membrane protein,OMP),是革兰氏阴性菌细胞的表面分子,在维持细胞结构、生命活动方面发挥重要作用,其暴露的抗原决定簇也可使外膜蛋白极易被免疫系统识别,从而引起机体的特异性免疫[6]。目前,已有基于外膜蛋白免疫原性的鱼类疫苗研究,如TANG 等[7]将鱼肠弧菌的5 个外膜蛋白进行原核表达,发现外膜蛋白T 具有良好的保护性;SUN 等[8]成功表达纯化荧光假单胞菌外膜蛋白ExbB,通过被动免疫保护评价,发现ExbB 对荧光假单胞菌感染的保护率可达54.0%,同时对嗜水气单胞菌感染具有交叉免疫保护作用,保护率为38.4%;LIU 等[9]表达的荧光假单胞菌外膜蛋白PF1380同样具有交叉被动免疫保护作用,对荧光假单胞菌感染的保护率为57%,对嗜水气单胞菌感染的保护率为50%;毛然然等[10]将嗜水气单胞菌外膜蛋白OprM 原核表达并进行免疫保护作用评价,发现外膜蛋白OprM 的保护率高达89.47%;荣娜等[11-12]成功表达纯化了嗜水气单胞菌外膜蛋白P5、TolC,这些外膜蛋白均具有免疫原性。综上所述,外膜蛋白具有一定免疫保护作用,同时也具有良好的应用前景,但嗜水气单胞菌外膜蛋白AHA1182的研究未见报道。为此,表达嗜水气单胞菌外膜蛋白AHA1182并探究其免疫原性,以期为嗜水气单胞菌疫苗的研制奠定基础。
1 材料和方法
1.1 材料
1.1.1 菌株及供试动物 嗜水气单胞菌ATCC 7966,大肠杆菌(E.coli)DH5α、BL21 菌株及pET-32a 质粒由陕西理工大学中德天然产物研究所保存;红鲫购自汉中市某水族馆,体表健康,平均体质量约20 g,身长平均约10 cm,试验前在90 L 水箱中饲养14 d,每天换水1/3,饲养用水经过处理,平均水温(20±2)℃,饲料(上海傲滋水族科技有限公司)每日按体质量的1%供给,使用气泵24 h 供给空气。饲养14 d后未见任何感染、死亡。
1.1.2 主要试剂 rTaq 聚合酶、ExTaq聚合酶、T4DNA 连接酶、内切酶BamH Ⅰ及XhoⅠ购自日本TaKaRa公司;细菌基因组提取试剂盒购自北京天根试剂公司;质粒提取试剂盒、胶回收试剂盒购自杭州博日公司;异丙基-β-D-硫代半乳糖苷(IPTG)、二甲基联苯胺(DAB)、DL2000 DNA marker、氨苄青霉素(AMP)购自北京索莱宝公司;TMB 显色液为上海生工公司产品;HRP 标记山羊抗大鼠IgG 购于Sigma 公司;碱性磷酸酶(AKP)、酸性磷酸酶(ACP)测定试剂盒购自南京建成生物工程研究所;弗氏不完全佐剂购自MP 生物医疗公司;引物合成、基因测序由西安奥科生物公司完成;大鼠抗鱼血清为中德天然产物研究所实验室自制。
1.2 方法
1.2.1 生物信息学分析 从NCBI 数据库中下载嗜水气单胞菌、杀鲑气单胞菌、简氏气单胞菌、多样气单胞菌、舒氏气单胞菌、产碱普罗维登斯菌以及爱德华氏菌与AHA1182 相同家族的外膜蛋白氨基酸序列,利用GeneDoc 3.2 软件进行序列对比,采用MEGA 5.0软件绘制系统发育树。
1.2.2 重组质粒构建 NCBI 数据库中查找嗜水气单胞菌ATCC7966 全基因组,根据全基因组设计AHA1182的 扩 增 引 物 ,上 游 引 物 :5′-AGAGGATCCGTGAATAAAGCGTTGAAA-3′,下 游引物:5′-ACCCTCGAGTTAGTTGCTCTTGCTTACC-3′(划线部分为酶切位点);扩增模板为嗜水气单胞菌全基因组;利用r Taq 聚合酶扩增,体系为50-µL。预变性温度、退火温度、退火时间分别为94 ℃、55 ℃、90 s;对PCR 产物进行电泳鉴定并切胶回收;使用BamHⅠ与XhoⅠ对PCR 产物与pET-32a 质粒进行双酶切,对二者分别切胶回收;通过T4 DNA 连接酶将目的片段与载体在16 ℃下连接16~18 h;将连接后的产物转入E.coliDH5α 中,转化成功后提取构建好的重组质粒;利用双酶切鉴定重组质粒是否构建成功;最后将重组质粒转入E.coliBL21中,进行相应蛋白质的原核表达。
1.2.3 重组蛋白的表达与纯化 挑取单菌落培养8~12 h,按1∶100 比例转接至含AMP 的培养基中培养3~4 h,0.1 mmol/L IPTG 诱导8 h。取1 mL 菌液并离心收菌,加入300µL 2×SDS上样缓冲液后沸水浴2 min,SDS-PAGE 电泳检测目的蛋白的表达。重组蛋白的纯化具体步骤:以1∶100 的比例将过夜培养的表达菌液转接至600 mL LB 培养液(Amp+)中;培养至OD600=0.5时,加入600µL 0.1 mmol/L IPTG,诱导8 h;4 000 r/min离心10 min,收集诱导菌体进行超声破碎,超声结束后,8 000 r/min 离心10 min,收集包涵体;将包涵体反复洗涤后进行SDS-PAGE 电泳;将目的条带切下研磨至无凝胶碎片,用蛋白质浸提液重悬,室温轻摇2 h;4 ℃、8 000 r/min 离心20 min,收集上清,加入4 倍体积预冷丙酮,-20 ℃过夜萃取;将萃取后的蛋白质离心,收集沉淀用无菌水溶解,测量浓度并冻干。
1.2.4 重组蛋白表达条件的优化 采用L9(34)正交试验模型(表1)探究目的蛋白的最佳表达条件。主要步骤:将过夜培养的蛋白质表达菌液以1∶200 的比例接入600 mL 培养液中,37 ℃、220 r/min 培养至设定的OD600值,按照表中要求加入不同浓度的IPTG,每组进行3 次重复。取1 mL 诱导菌液,离心收菌后菌体加入300µL 2×SDS蛋白质上样缓冲液,沸水浴2 min,SDS-PAGE 电泳获得不同诱导条件下的AHA1182 蛋白表达图谱。通过软件Phoretix 1D 1.0.0.1 和SPSS 20.0 分别对表达图谱光密度和不同因子作显著性分析。
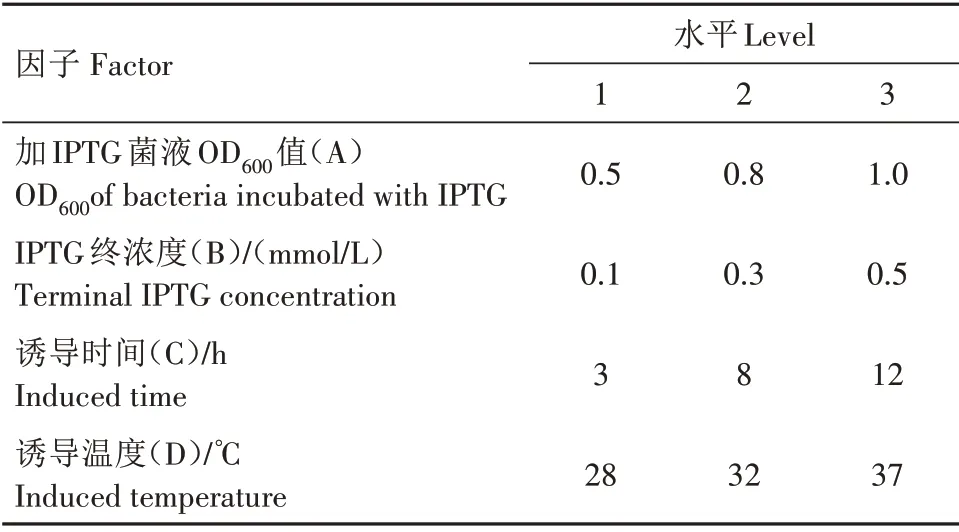
表1 蛋白质表达条件正交试验的因子与水平Tab.1 Factors and levels of orthogonal test for protein expression condition
1.2.5 红鲫抗血清特异性检测 红鲫免疫组与对照组各35条,免疫组与对照组均采用弗氏不完全佐剂乳化后进行免疫。免疫组按照2µg/g的剂量进行免疫,红鲫腹腔注射免疫AHA1182 蛋白,14 d 后进行第2 次免疫,7 d 后进行第3 次免疫,操作同上;对照组免疫无菌水。7 d后尾静脉取血。应用Western blot夹心法验证红鲫抗血清特异性,步骤如下:SDSPAGE 电泳嗜水气单胞菌全蛋白;80 V、1 h 电转于硝酸纤维素(NC)膜上,置于5%脱脂牛奶粉中充分封闭;将封闭好的NC 膜与不同稀释倍数(1∶100、1∶200、1∶400、1∶800、1∶1 600、1∶3 200)的红鲫抗血清、阴性对照血清在37 ℃中孵育40 min,TBST 溶液洗涤3 次,每次10 min;再与1∶400 的大鼠抗鱼血清孵育,具体步骤同上;最后与1∶3 000 山羊抗大鼠抗体孵育;洗涤后进行DAB显色,确定抗原-抗体的结合情况。
1.2.6 ACP、AKP 活性测定 采用南京建成生物工程研究所试剂盒进行ACP、AKP 测定,严格按说明书步骤进行。
1.2.7 体外细胞吞噬试验 吞噬细胞是机体免疫系统中重要的细胞成分之一,吞噬微生物和其他异物颗粒,在机体免疫应答中起重要作用。具体步骤:取免疫AHA1182 蛋白的红鲫血浆200 µL 与1%甲醛生理盐水灭活的金黄色葡萄球菌(6×108cfu/mL)1∶1 混合,25 ℃水浴60 min(每10 min 振荡混匀一次);以免疫无菌水的红鲫血浆为对照。反应结束后制作血涂片,用快速姬姆萨染色试剂盒(上海生工)进行染色。
1.2.8 体外模拟抗AHA1182 蛋白抗体与嗜水气单胞菌的相互作用 利用酶联免疫吸附(ELIAS)法[13]检测免疫红鲫血浆与嗜水气单胞菌的相互作用。具体步骤:嗜水气单胞菌培养至OD600约1.0后,将菌体离心并洗涤;将菌体OD600调整至1.0(菌浓度1.0×109cfu/mL),分装菌体每管150µL,再与不同稀释倍数的血浆(1∶100、1∶200、1∶400、1∶800、1∶1 600、1∶3 200)37 ℃条件下孵育90 min,对照为阴性血浆;再与1∶400 的大鼠抗鱼血清孵育90 min;最后与1∶3 000山羊抗大鼠抗体孵育1 h。将洗涤好的菌体与20 µL PBS 混合后加入酶标板,再加入200 µL 的TMB 显色液;最后用50µL 2 mol/L H2SO4终止反应,450 nm读数。
1.2.9 病理切片观察 以免疫结束的红鲫肾脏、脾脏、肠为组织病理学切片材料。病理切片主要有3个步骤,即组织包埋、组织切片与H&E染色:(1)组织包埋:将组织立即浸泡在改良的Davidson’s[14]固定液中,固定18~24 h 后转入10%甲醛溶液中继续固定24 h;将固定好的组织于梯度体积分数(80%、90%、95%、100%)的乙醇中脱水,再放入二甲苯中透明;将透明好的组织于60 ℃的石蜡中充分浸蜡,再包埋于折叠好的模具中。(2)切片:将包埋好的组织修正成约1 cm3的立方体,石蜡切片机切成厚度为4µm 的切片,并将其附着于载玻片上,烘干。(3)H&E 染色:使用苏木素与伊红进行染色。首先将烘干的切片放入二甲苯中脱蜡;再于梯度体积分数的乙醇中复水,复水结束后苏木素染色30 s,使细胞核上色,伊红染色20 min,使细胞质着色。继续于梯度体积分数的乙醇中脱水,在二甲苯中透明,最后用中性树脂封片即可。
1.3 数据分析
试验结果以平均值±标准差表示,采用SPSS 20.0 软件进行单因素方差分析,并用GraphPad Prism 8.0.1软件绘图。
2 结果与分析
2.1 不同菌株间外膜蛋白AHA1182同源性与系统发育分析
利用GeneDoc 3.2 软件对不同菌株与嗜水气单胞菌外膜蛋白AHA1182的氨基酸序列进行比对,结果显示,气单胞菌间具有高度的同源性,分别为98.8%(杀鲑气单胞菌)、73.5%(舒氏气单胞菌)、72.9%(多样气单胞菌)、90.4%(简氏气单胞菌),而不同属间菌株也具有一定的同源性(图1)。以MEGA 5.0 软件构建的进化树显示,2 种不同的气单胞菌亲缘关系较近(嗜水气单胞菌、杀鲑气单胞菌),且不同种菌株间也有一定的亲缘关系(图2)。因而,动物免疫AHA1182蛋白后产生的抗体可能对不同种属的细菌有交叉免疫的作用。
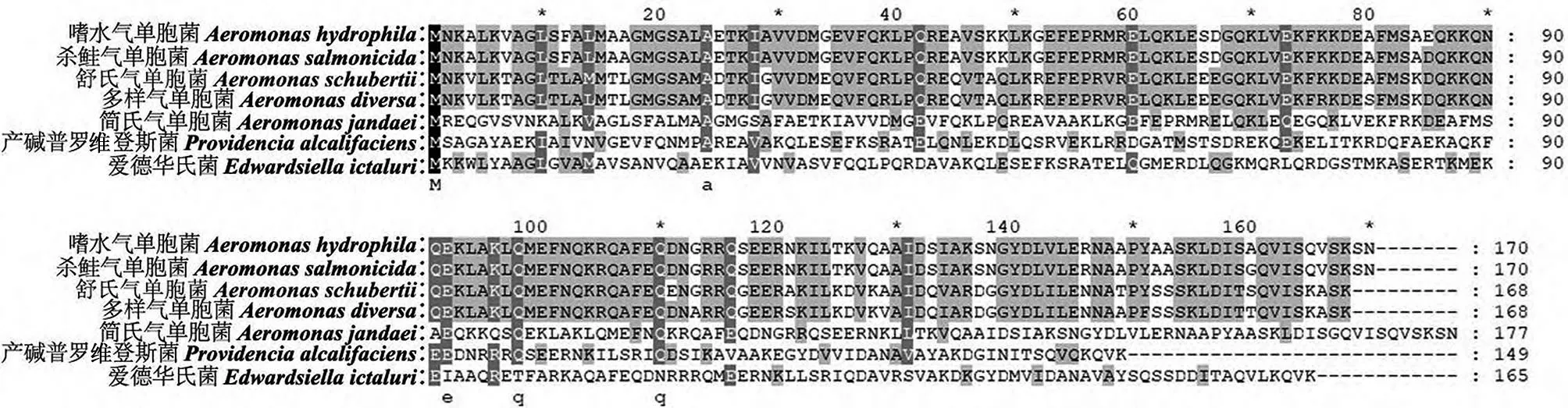
图1 不同菌株间外膜蛋白AHA1182氨基酸序列比对结果Fig.1 Alignment results of amino acid sequences of outer membrane protein AHA1182 among different strains
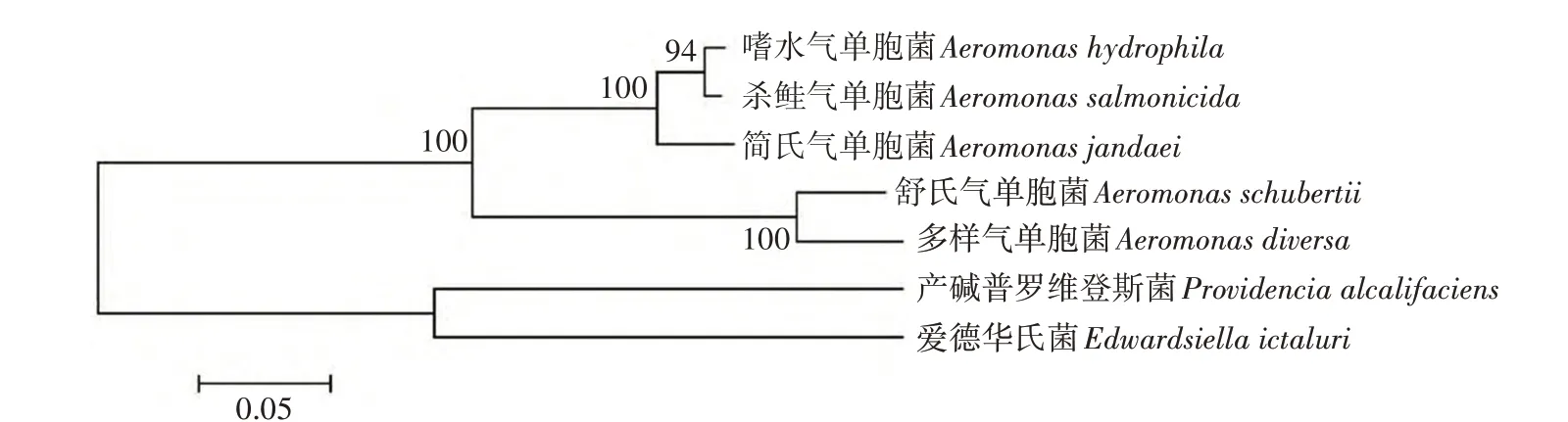
图2 不同菌株间外膜蛋白系统发育树Fig.2 Phylogenetic tree of outer membrane proteins among different strains
2.2 外膜蛋白AHA1182的表达纯化
利用PCR 扩增AHA1182基因,结果如图3A 所示,在约500 bp 处有单一且清晰的条带,与理论值一致;重组质粒pET-32a-AHA1182经过BamH Ⅰ与XhoⅠ双酶切鉴定,在约500 bp处得到一条带,与理论值相同,重组质粒构建成功(图3B)。重组质粒经转化E.coliBL21,IPTG诱导后,通过SDS-PAGE电泳可得1条约35 ku的条带,表示目标蛋白AHA1182表达成功(图3C);利用包涵体洗涤、SDS-PAGE 切胶纯化后获得的目的蛋白如图3C所示。
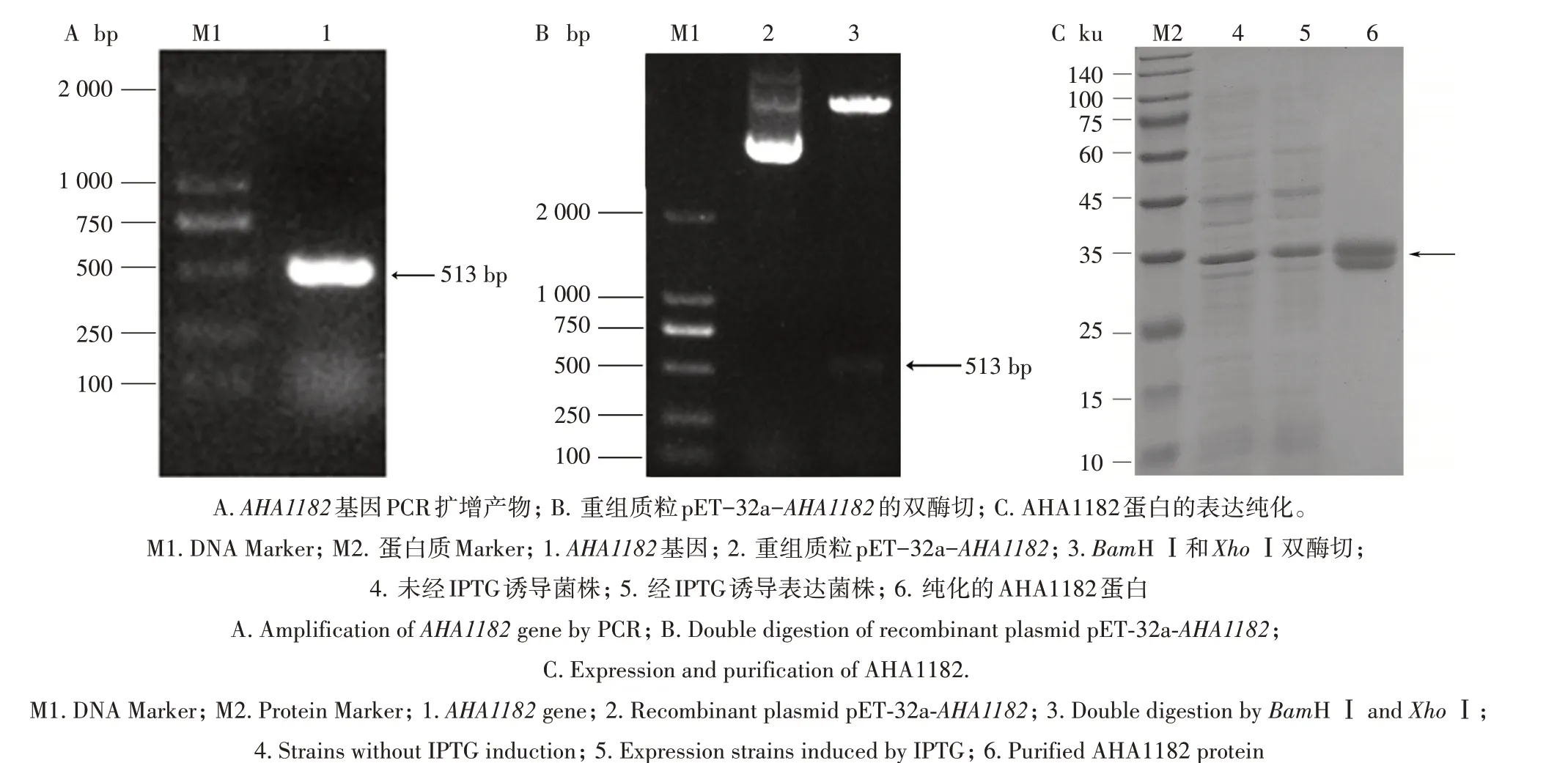
图3 AHA1182重组质粒的构建及蛋白质纯化检测Fig.3 Construction of recombinant plasmid and protein purification of AHA1182
2.3 AHA1182蛋白表达条件的优化
诱导菌液经SDS-PAGE 电泳获得不同诱导条件下的表达图谱,图谱光密度值分析如表2所示,所得数据极差和方差分析如表3、表4 所示。综合分析,AHA1182蛋白的最佳表达条件为A3B3C2D1,即菌液浓度OD600=1.0,IPTG 终浓度为0.5 mmol/L,在28 ℃下诱导8 h;通过显著性分析可知,IPTG 浓度对AHA1182蛋白的高效表达有重要作用。

表2 AHA1182蛋白表达图谱光密度分析Tab.2 Optical density analysis of AHA1182 expression map
表3 AHA1182表达图谱光密度值的极差分析Tab.3 Range analysis of optical density of AHA1182 expression map

表4 AHA1182表达图谱光密度值的方差分析Tab.4 Variance analysis of optical density of AHA1182 expression map
2.4 AHA1182蛋白抗血清的特异性与效价检测
Western blot试验结果(图4)显示,约在23 ku处出现特异性条带,阴性血清无条带,表明红鲫血清中存在抗体与嗜水气单胞菌特异性结合。说明AHA1182具有免疫原性,并正确激活了红鲫的特异性免疫,血清效价为1∶100(图4)。

图4 AHA1182蛋白抗血清的特异性与效价检测Fig.4 Detection of specificity and titer of AHA1182 protein serum
2.5 免疫AHA1182蛋白红鲫血清的ACP、AKP活性测定结果
红鲫在免疫AHA1182 蛋白后,与对照组相比,血清的ACP、AKP 均极显著性(P<0.01)升高(图5),表明外膜蛋白AHA1182 可激活红鲫体内的非特异性免疫。
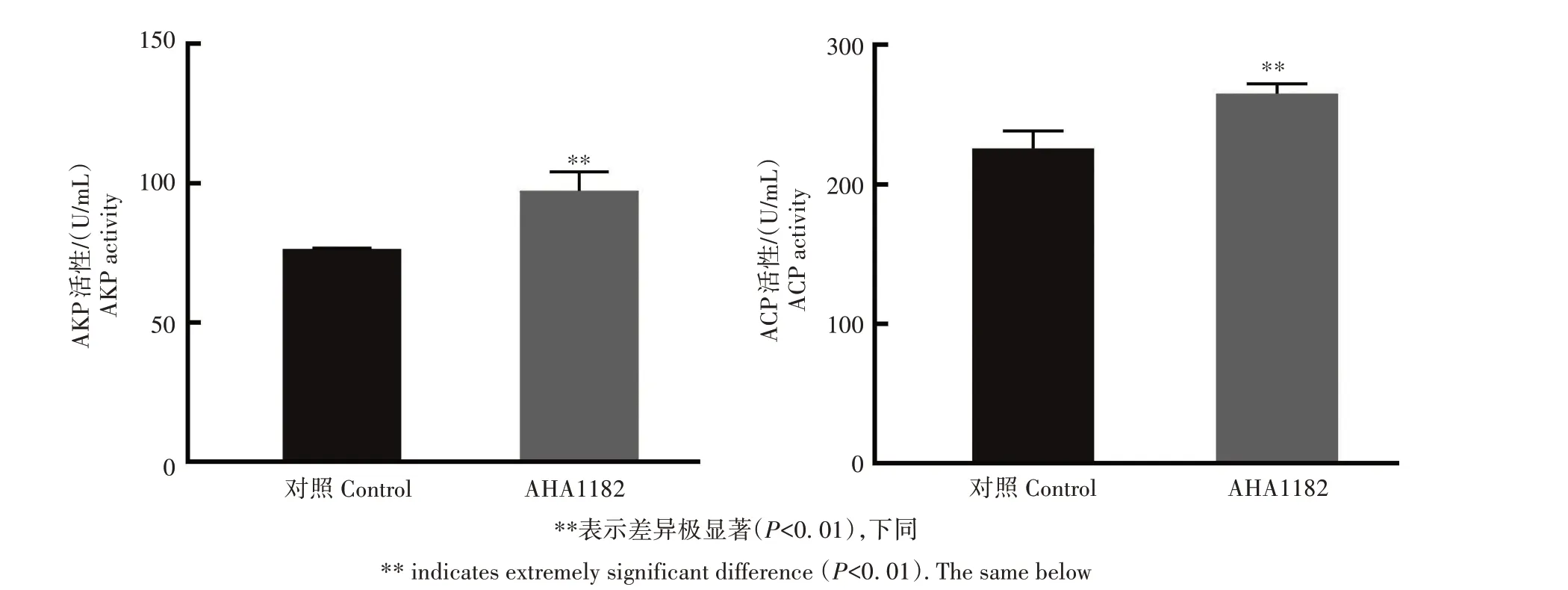
图5 AHA1182蛋白抗血清的AKP、ACP活性Fig.5 Activity of AKP and ACP in serum of C.auratus immunized with AHA1182 protein
2.6 免疫AHA1182蛋白红鲫血浆的体外细胞吞噬作用
免疫AHA1182 蛋白红鲫血浆的吞噬作用结果如表5 所示。与对照组相比,免疫AHA1182 蛋白红鲫血浆细胞吞噬作用明显升高:吞噬百分数升高,且吞噬指数极显著升高(P<0.01)。

表5 免疫AHA1182蛋白后红鲫血浆的细胞吞噬作用Tab.5 Leukocyte phagocytosis of C.auratus plasma after immunization with AHA1182
2.7 免疫AHA1182蛋白红鲫血浆与嗜水气单胞菌的相互识别作用
利用ELISA 进行体外相互作用模拟,结果显示,免疫AHA1182蛋白后红鲫血浆与嗜水气单胞菌相互作用强度随血浆的稀释度增大而减小,可证明抗AHA1182 蛋白抗体与嗜水气单胞菌在体外具有相互识别作用,且滴度可达1∶3 200(图6)。
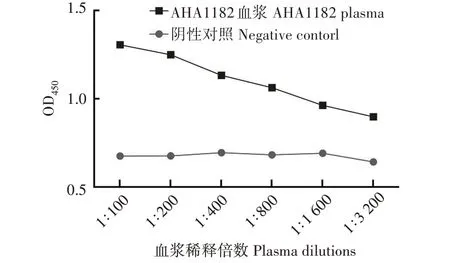
图6 抗AHA1182蛋白抗体与嗜水气单胞菌体外相互作用Fig.6 AHA1182 protein antibody interacts with Aeromonas hydrophila in vitro
2.8 AHA1182蛋白对红鲫内脏的影响
对红鲫肾脏、脾脏、肠组织进行组织病理学切片(图7),对照组肾脏组织可见细胞排列紧凑,肾小管、肾小球无明显损伤(7a),脾脏结构紧凑,淋巴细胞数量相对较多(7b),肠组织杯状细胞排列整齐,结构完整(7c);AHA1182 蛋白免疫组与对照组相比内脏组织无明显区别,表明免疫外膜蛋白AHA1182对红鲫内脏无影响。
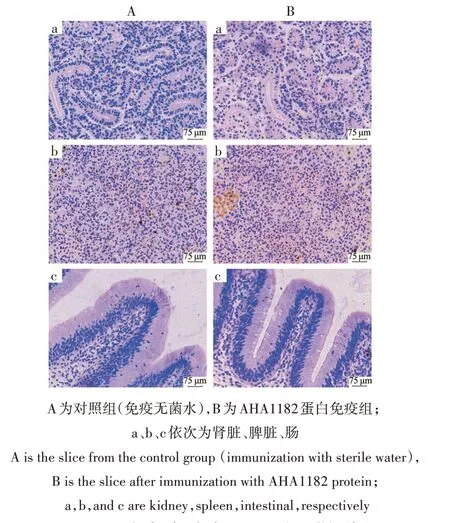
图7 红鲫肾脏、脾脏、肠组织病理学切片Fig.7 Pathological sections of kidney,spleen and intestine of C.auratus
3 结论与讨论
嗜水气单胞菌是广泛存在于各种水体中的病原菌,其对渔业养殖的危害性已引起公众的注意,且目前的防治方法均存在缺陷。长期使用抗生素防治会使病原菌产生耐药性;饲料中维生素与中草药的添加虽会提高鱼类的抗菌力,但防治效果不明显;益生素的使用能够提高一部分鱼类的自身免疫力,但适用范围并不广泛[15]。因此,本研究选取嗜水气单胞菌外膜蛋白AHA1182进行主动免疫评价,为研制嗜水气单胞菌渔用疫苗寻找新的候选抗原。
本研究成功构建了AHA1182原核表达菌株,正交试验确定了最佳表达条件:菌液浓度OD600=1.0,IPTG 终浓度为0.5 mmol/L,在28 ℃条件下诱导8 h。显著性分析表示,IPTG 浓度对诱导表达有极显著的影响。有研究表明,低温培养有助于蛋白质的可溶性表达,可减少包涵体的形成[16],本研究结果(28 ℃)印证了这一结论。诱导时间因素对蛋白质表达具有重要影响,有研究表明,诱导时间过长会导致蛋白质被蛋白酶降解,不利于蛋白质的表达[17],这与本研究结果(8 h)一致。IPTG 具有一定的毒性,过高浓度的IPTG 会抑制细菌生长[18],适度的IPTG 浓度对蛋白质的表达量具有重要作用。本研究结果表明,IPTG 浓度对蛋白质表达有极显著的影响,其最佳浓度为0.5 mmol/L。最佳诱导条件的确定,为AHA1182蛋白的批量纯化并进行免疫学研究奠定了基础。
免疫原性指能够刺激特定的免疫细胞,使免疫细胞活化、增殖、分化,产生抗体从而引起免疫应答的抗原。本研究通过体外模拟红鲫血清与嗜水气单胞菌的识别作用及Western blot 检测AHA1182 的免疫原性。体外模拟试验结果表明,抗血清与细菌具有相互作用,且效价可达1∶3 200;Western blot 检测结果表明,红鲫抗血清具有特异性。这与前人研究结果基本一致[19-21]。抗原-抗体结合需要相应的抗体[22],而体外模拟的结果说明了红鲫血清可识别嗜水气单胞菌,并通过抗原呈递作用,将体内病原菌清除。本研究利用纯化后的外膜蛋白AHA1182免疫红鲫,发现AHA1182 蛋白具有一定的免疫原性,为渔用疫苗的开发奠定了生物学基础。
非特异性免疫是鱼类免疫病原微生物的主要方式,吞噬作用与免疫相关的酶类是非特异性免疫的重要组成部分。ACP、AKP作为鱼类主要解毒酶,广泛存在于鱼类体液与脏器中[23-24]。因此,吞噬细胞活性、AKP、ACP 可作为非特异性免疫的评价指标。本研究发现,对红鲫免疫AHA1182 蛋白后,红鲫血清中AKP、ACP 均显著升高;同时,红鲫血浆中细胞吞噬作用的指标值均升高。可见,AHA1182蛋白可激活红鲫的非特异性免疫。此外,SUN 等[8]通过生物信息学分析,发现荧光假单胞菌外膜蛋白ExbB进化保守,且具有交叉免疫保护作用。本研究选取6 株具有AHA1182 相同家族外膜蛋白的鱼类致病菌,并与嗜水气单胞菌外膜蛋白AHA1182进行氨基酸序列比对、系统发育树构建,发现外膜蛋白AHA1182 进化保守,因此,蛋白AHA1182 也可能具有交叉免疫保护作用,有望成为多价疫苗的候选抗原。
组织病理学切片观察可直观反映组织、细胞的损伤情况。张海彬等[25]利用病理学组织切片观察感染嗜水气单胞菌的乌鳢内脏损伤情况;兰蕾等[26]对艾烟染毒后大鼠的心脏、肝脏、脾脏、肺等组织进行组织病理学切片观察,发现染毒后均有损伤。本研究利用组织病理学切片观察,发现AHA1182蛋白的免疫对红鲫内脏无明显影响。
综上所述,AHA1182 蛋白具有较强的免疫原性,能够激活红鲫非特异性免疫,且对红鲫内脏无明显影响。本研究证明了嗜水气单胞菌外膜蛋白AHA1182可作为一种保护性抗原,为嗜水气单胞菌疫苗的开发奠定了理论基础。
