南亚热带海岸带沙地次生林群落根系形态特征
2022-01-21卢立国吕宗晋谢伟东义柏芊
卢立国,吕宗晋,谢伟东,义柏芊
(广西大学 林学院,广西南宁 530004)
根系作为植物地上部分的支撑者,能通过提供水分和养分对植物生长与繁殖产生决定性作用,还能通过提高土壤通透性、增加土壤生物活性物质等改善土壤性质,进而影响植物生长的微环境,使植物表现出独特的生物学特性和抵抗不良环境的能力[1-4]。植物在长期的生长发育过程中,根系形态特征的响应与土壤资源利用能力紧密相联,能反映植物对环境的适应策略[5-6]。不同生境的植物根系形态特征研究在根系生态学领域中十分重要。马雄忠等[7]研究阿拉善高原缘红砂(Reaumuria soongari⁃ca)和珍珠猪毛菜(Salsola passerina)对干旱生境的生态适应策略,发现这两种植物根系分支相对简单,但空间拓展能力强,有利于获取营养。黄同丽等[8]研究喀斯特地区灌木植物的根系构型,发现均呈现鱼尾形分支结构,次级分支均较少且分支较简单。杨森霖等[9]对干热河谷区不同坡位引种巨菌草(Pennisetum sinese)的根系生长特性进行研究,发现不同坡位各等级根序根系形态指标存在差异,中坡坡位的巨菌草生长状况最佳。王浩等[10]对砒砂岩区3年生中国沙棘(Hippophae rhamnoides)根系进行研究,发现坡位对沙棘根系构型影响最小,随坡位的上升根冠比表现出先增后减的趋势。
海岸带沙地因特殊的地理环境,具有土质盐渍化、土壤贫瘠和地下水位高的特点;经常受台风、暴雨等灾害天气的影响,水源涵养能力极差,生态环境十分脆弱[11-12]。海岸带沙地环境研究主要集中在群落结构特征[13]、水文生态效益[14]和林地土壤理化性质[15-16]等方面,植物根系形态的研究相对较少。关于沙地植物根系形态的研究主要集中在内陆地区[7,10],海岸带沙地植物根系研究以东部沿海地区人工林的研究为主[17],未见针对南亚热带海岸带原生天然林、次生林群落的报道。天然林作为当地生境的主要建群种和优势种,其根系形态特征可反映根系在南亚热带海岸带沙地特殊生境条件下的生态对策。为了解群落植物根系在海岸带沙地特殊生境中的适应机制,本研究在广西防城港东兴市选取2 个海岸带次生林群落为对象,使用Win-RHIZO 根系分析系统分析不同植物群落的根系形态特征,阐明海岸带植被的根系特征和环境适应性,可为海岸带天然林保护和防护林建设提供参考。
1 材料与方法
1.1 研究区概况
研究区位于东兴市东兴镇竹山村海岸带(107°53'~108°15'E,21°31'~21°44'N),海拔10 m以下,坡度11°~22°。东兴市位于广西南部,北回归线以南,属南亚热带季风气候,全年气候温和湿润,冬短夏长;年均气温23.2 ℃以上,10 ℃以上年积温8 000 ~ 8 200 ℃,最低气温2 ℃;年均遭受0.3 ~ 1.0次寒潮侵袭,年均日照时长1 500 h 以上;年均降水量2 738 mm,春、夏季是降雨高峰期,雨热同期,盛行台风。该区域为滨海平原,地表被海相石英细砂、砂砾和亚粘土覆盖。
1.2 研究方法
1.2.1 样地设置
在研究区选取相邻两个天然次生林群落进行根系采样。两个群落分别是假苹婆(Sterculia lan⁃ceolata)+ 红鳞蒲桃(Syzygium hancei)-长花龙血树(Dracaena angustifolia)+ 黄脉九节(Psychotria stra⁃minea)-华山姜(Alpinia chinensis)群落(简称假苹婆群落,下同)和鸭脚木(Heptapleurum heptaphyllum)+红鳞蒲桃-罗伞树(Ardisia quinquegona)+ 长花龙血树-沿阶草(Ophiopogon bodinieri)群落(简称鸭脚木群落,下同)(表1)。
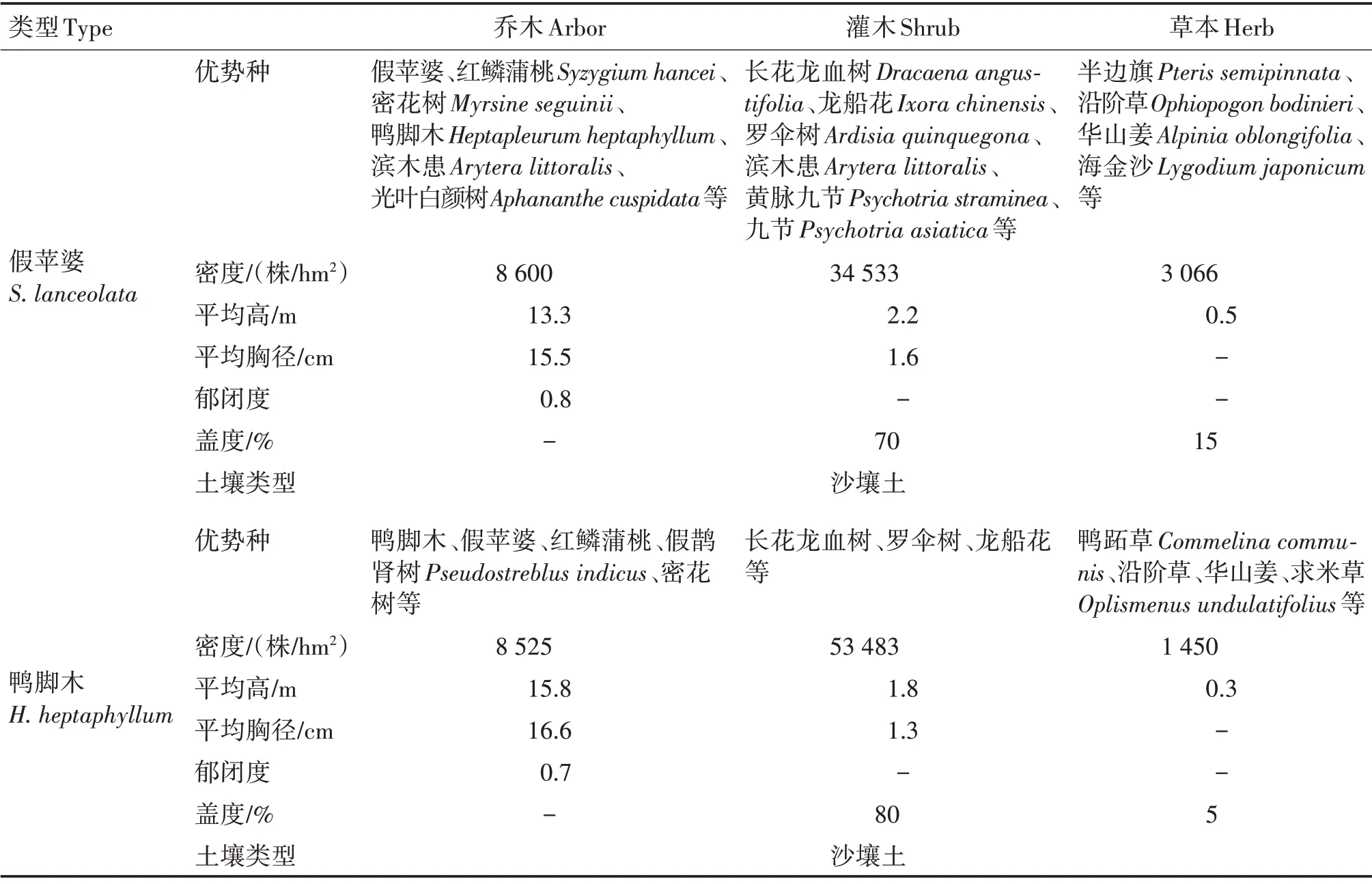
表1 群落基本概况Tab.1 General situation of community
1.2.2 根系采样方法
采用挖土方的方法采集根系,在两个群落的上、中和下坡分别沿水平方向相距10 m设置3个20 m×20 m的样方,在各样方对角线的交叉点用平铲挖取1个20 cm×20 cm×20 cm的土块,每群落每坡位3个处理,共27个处理。分别收集土块中的所有根系,小心清理根系上粘附的土壤和杂质,称重后装入封口袋并贴上标签,保湿带回实验室,在冰箱中保存。
1.2.3 根系形态指标测定及分级
将根系样品用水浸泡,去除土壤和杂质,随后放入60 目筛网,用水反复冲洗,将干净的根系放入塑料袋内并保持湿润,30 min内测定根系形态。
采用根系分析系统(Win-RHIZO 5.0)对根系样品进行扫描,分析10个径级(0 mm <d1≤0.5 mm、0.5 mm <d2≤1.0 mm、1.0 mm <d3≤1.5 mm、1.5 mm <d4≤2.0 mm、2.0 mm <d5≤2.5 mm、2.5 mm <d6≤3.0 mm、3.0 mm <d7≤3.5 mm、3.5 mm <d8≤4.0 mm、4.0 mm <d9≤4.5 mm和4.5 mm <d10≤5.0 mm)细根的根长、根表面积、根尖数和分叉数,计算根尖密度、分叉密度、细根长度百分比和细根表面积百分比。
根系分级采用Pregitzer 等[18]的根序分级方法,将每个采样点的根系样品分成5级根序。最远端的具有根尖的根或根系最末梢的根定义为1级细根,1级细根的母根定义为2级细根,2级细根的母根定义为3 级细根,依次类推直至区分完5 级细根,5 级细根的母根及之后或大于5 mm 的根为粗根。测量各级细根根序的根长和根表面积。用镊子取下每一级别细根,分袋标号后放入烘箱,80 ℃连续烘24 h至恒重,用电子天平称量根干质量(精确至0.001 g),计算各级细根的生物量、比根长和比表面积。
1.2.4 根系指标计算
采用下列公式计算根系各指标[19-20]。

1.3 数据处理
采用Excel 软件整理数据;采用SPSS 软件进行LSD 单因素方差分析和差异显著性检验;采用Ori⁃gin软件作图和计算。
2 结果与分析
2.1 根系形态特征
两个群落的各项根系形态指标差异均不显著,鸭脚木群落的各项根系形态指标均大于假苹婆群落(表2)。假苹婆和鸭脚木群落的总根长分别为5 442和5 924 cm,总表面积分别为140.94 和156.71 cm2,根尖数分别为21 680.67 和35 725 个,分叉数分别为2 724.33 和3 780 个,根尖密度分别为3.93 和5.44 个∕cm,分叉密度分别为0.50和0.56个∕cm。

表2 根系形态指标Tab.2 Root morphological indexes
2.2 根序分级与数量特征
2.2.1 根系比根长
细根比根长在一定程度上反映细根吸收水分和养分的能力。两个群落的各级细根比根长和占比均表现为1 级>2 级>3 级>4 级>5 级(表3)。两个群落的1 级细根比根长分别为2 327 和1 913 cm∕g,占比分别为50.48%和55.03%。两个群落的同一根序比根长间差异不显著,1、2 级细根的比根长明显大于其他根序。
2.2.2 根系比表面积
假苹婆群落各级细根比表面积和占比均表现为2 级>3 级>1 级>4 级>5 级,鸭脚木群落表现为2级>1 级>3 级>4 级>5 级(表3)。随根序级别增加,两个群落的各细根比表面积呈先上升后下降的趋势,2 级细根的比表面积均最大。两个群落的细根比表面积均以1 ~ 3 级细根为主,占比分别为95.22%和92.46%;两个群落同一根序的比表面积间差异不显著。
2.2.3 根系生物量
假苹婆群落各级细根生物量和占比均表现为5级>4 级>1 级>2 级>3 级,鸭脚木群落表现为5级>3 级>1 级>4 级>2 级,假苹婆群落总生物量显著大于鸭脚木群落(P<0.05)(表3)。两个群落同一根序的生物量均差异不显著;5 级细根生物量最大,占比分别为71.60%和48.98%。
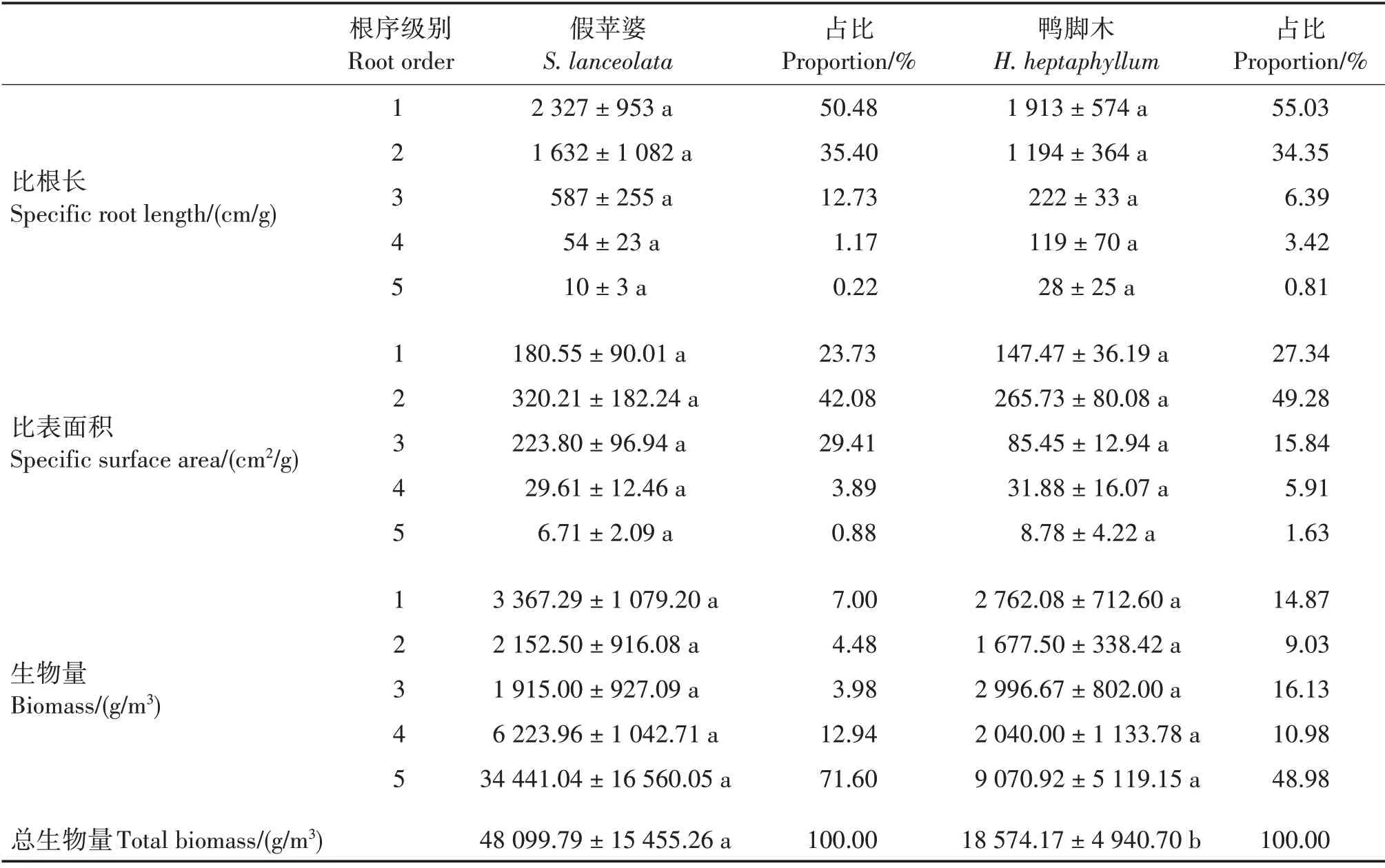
表3 不同根序的细根比根长、比表面积和生物量及其占比Tab.3 Species root length,specific surface area,biomass and their proportions of fine roots in different root orders
2.3 细根分级与数量特征
2.3.1 不同径级细根的长度和表面积分布特征
两个群落0 mm<d1≤0.5 mm径级的细根根长占比最大,分别为55.14%和58.79%;其次为0.5 mm<d2≤1.0 mm 径 级,分 别 为25.28% 和24.31%;d>2 mm 径级的细根根长占比较小,分别为8.04%和8.92%(图1)。随着径级增大,细根根长占比下降。两个群落各径级细根根长的比例相近。

图1 不同径级细根长度百分比Fig.1 Percentages of fine root length in different diameter classes
两个群落0.5 mm<d2≤1 mm 径级的细根表面积占比最大,分别为25.64%和26.94%;其次为0 mm<d ≤0.5 mm 径级,分别为19.57%和23.89%(图2)。d>2 mm径级的细根表面积占比较小。

图2 不同径级细根表面积百分比Fig.2 Percentages of fine root surface area in different diameter classes
2.3.2 不同坡位细根形态特征
假苹婆群落中,上坡的比根长、比表面积和生物量最大,均显著大于中、下坡(P<0.05),下坡最小(表4)。鸭脚木群落中,中坡的比根长和比表面积最大,与上、下坡差异显著(P<0.05);下坡的生物量最大,与上坡差异不显著。同坡位下,假苹婆群落的总生物量均大于鸭脚木群落;假苹婆群落上坡的比根长和比表面积远大于鸭脚木群落上坡,中、下坡则小于鸭脚木群落。

表4 不同坡位细根形态特征Tab.4 Root morphological characteristics of fine roots in different slope posotions
3 讨论与结论
3.1 根系形态指标
植物根系具有很强的可塑性,为了适应生存环境,根系形态和空间分布会产生一定的变化[21]。本研究中两个群落根系的分叉密度(0.50 和0.56 个∕cm)小于刘佳等[19]和胡慧等[22]的研究结果,但根尖密度大于两者,原因可能与土壤结构、水分和养分的分布有关;与内陆植被相比,海岸带降水较多,表层土壤水分可利用率较高,植物根系受到的环境胁迫较小,不需要增加根系分支即可满足自身生长[11]。由于两个群落的地理位置、气候条件和土壤理化性质基本一致,根分叉少,各项根系形态指标均差异不显著。
3.2 各根序的细根形态特征
树木根系在长期进化过程中形成分支结构,使根系分工明确,比根长和比表面积是重要的根系形态指标,表示根系生理功能的情况,反映植物对不同生境的适应特征[23]。有关学者研究我国亚热带和小兴安岭多个树种的根系形态,结果显示1 级根单个根长最短、比根长最大,5 级根相反[24-26]。师伟等[27]研究帽儿山天然次生林20 个阔叶树种细根形态,发现前3 级根累计根长占前5 级根总长度的80%以上;庄丽燕等[28]发现川西亚高山3 个优势树种根系的比根长和比表面积在1 级根最大,随序级升高而减小,5 级根降到最低。本研究中,比根长和比表面积指标与上述研究结果总体趋势相似,1~3序级的根系是群落根系的主要部分。根序级别小的比根长和比表面积较大,扩大根系与土壤进行水分和养分交换的面积[30]。
根系生物量的分布格局代表根系对土壤资源的获取能力[31]。假苹婆和鸭脚木群落的5 级细根生物量分别占总生物量的71.60%和48.98%,与张咪等[32]和熊德成等[33]研究结果类似,表明两个群落对总生物量的贡献主要集中在根序级别大的根。梅莉等[34]认为根系生物量在养分和水分充足的表层土壤分布较多,在贫瘠的深层土壤中分布较少。干旱条件下,沙地植物会倾向于将更多根系分布在浅层土壤中,以更好地吸收降水[35]。南亚热带滨海沙地植物受土壤水分、养分分布的影响,也表现出浅根性,植物根系集中分布于土壤表层,体现两种天然植物群落对海岸带沙地生境长期适应的结果。
3.3 细根形态特征
不同径级的根系具有不同的功能。Fransen等[36]认为植物根系中直径≤1 mm 的细根能充分吸收和利用土壤中的矿质营养和水分并分泌次生代谢产物。李勇[37]认为≤1 mm 径级的根系对土壤结构和渗透性的改善最为明显。Hendrick 等[38]研究认为80%的细根长度是由直径<1 mm 的细根组成。本研究中,两个群落0<d ≤0.5 mm 径级的细根根长和和0.5<d ≤1 mm 径级的细根表面积对细根总细根长、细根总表面积贡献最大,与权伟等[20]的研究结果相同。直径≤1 mm 的细根在根系动态活动中最为活跃。
植物在不同生境条件下的资源分配策略反映植物生长对不同养分环境的反馈能力[39]。王浩等[10]发现随坡位下降,沙棘根长和根数逐渐增大,根系生物量先增大后减小。本研究中,假苹婆群落的比根长、比表面积和生物量都是在上坡位最大,下坡位最小;鸭脚木群落的比根长、比表面积在中坡位最大,生物量在下坡位最大,这种能量分配形式有利于两个群落在不同坡度的立地条件下加强对土壤和光资源的竞争和利用。
在未来海岸带沙地植被恢复与经营管理中,合理控制群落结构,选用合适的树种,改善土壤结构和理化性质,可促进群落根系水平和垂直方向的生长,有利于植物健康成长。本研究基于群落根系特征进行研究,由于群落的树种组成较为复杂,密度较大、根系交错,没有按照单一植物收集根系,未能探明不同树种间的根系形态差异。今后可按照不同树种纯林或混交林研究海岸带防护林植物根系的形态特征、生境适应性和养分吸收效率,进一步为海滨沙地植被恢复的树种选择与群落配置提供依据。
