ZNF292 suppresses proliferation of ESCC cells through ZNF292/SKP2/P27 signaling axis
2022-01-17WeiGongJianchengXuGuangchaoWangDanLiQiminZhan
Wei Gong ,Jiancheng Xu ,Guangchao Wang ,Dan Li ,Qimin Zhan,2
1State Key Laboratory of Molecular Oncology,National Cancer Center/National Clinical Research Center for Cancer/Cancer Hospital,Chinese Academy of Medical Sciences and Peking Union Medical College,Beijing 100021,China;2Key Laboratory of Carcinogenesis and Translational Research (Ministry of Education/Beijing),Laboratory of Molecular Oncology,Peking University Cancer Hospital &Institute,Beijing 100142,China
Abstract Objective:Increasing evidence has demonstrated that ZNF292 plays a suppressive role in cancer,however,little is known about its function and exact mechanism in esophageal squamous cell carcinoma (ESCC).Methods:Bioinformatic analysis and immunohistochemistry (IHC) were performed to analyze the role of ZNF292 in affecting the prognosis of ESCC.Cell proliferation and colony formation ability assays were performed to analyze cell growth after inferring the expression of ZNF292.Flow cytometry was used to analyze changes in the cell cycle upon the depletion of ZNF292.Quantitative real-time polymerase chain reaction (qRT-PCR) and western blot analysis were used to determine the alteration of cell cycle related RNAs and proteins after knocking down ZNF292.MG-132,cycloheximide (CHX) treatment experiments were performed to analyze the change and half-life time of P27 after knockdown of ZNF292.Chromatin immunoprecipitation (ChIP) and luciferase reporter assays were used to analyze the transcriptional regulation of SKP2 by ZNF292.Results:We report that low expression of ZNF292 is associated with poor prognosis,and ZNF292 emerges to be highly expressed in adjacent and normal tissues rather than tumor tissues in ESCC.Knockdown of ZNF292 significantly boosts cell growth and S phase entry in ESCC cells.ZNF292 depletion will decrease the expression and half-life time of P27,while knockdown of SKP2 will result in elevated expression of P27.ZNF292 can bind to the promoter region of SKP2,and knockdown of ZNF292 will boost the expression of SKP2.Conclusions:Knockdown of ZNF292 mediates G1/S cell cycle procession by activating SKP2/P27 signaling in ESCC cells.ZNF292 knockdown promotes SKP2 expression at the transcriptional level,thereby boosting P27 ubiquitin-degradation,and eventually facilitating the S phase entrance.
Keywords:ZNF292;cell cycle;SKP2;ESCC
Introduction
Esophageal cancer is one of the most malignant tumors in the world,ranking sixth among all the cancer-related death(1).Esophageal cancer has become one of the most malignant cancer types that causes cancer deaths in China(2).Pathologically,esophageal cancer can be divided into two types:esophageal squamous cell carcinoma (ESCC)and esophageal adenocarcinoma (EAC),of which nearly 90% of ESCC patients come from China (3).The lack of an exact mechanism for the occurrence and malignant development of ESCC has severely hindered the exploration of effective targeted therapeutic drugs (4,5).
The occurrence and development of malignant tumors are often accompanied by the activation of oncogenes and the inactivation of tumor suppressor genes.Furthermore,the bulk of the phenotypes that affect the clinical progression and therapeutic responses of patients,are likely to be strongly regulated by transcriptional programs operating within cancer cells (6).DNA-templated reactions,including transcription and replication,as well as DNA repair,largely rely on proteins binding to DNA that regulate these processes locally.Local concentrations of proteins on DNA are vital to exert their functions in a coordinated manner (7).Therefore,the transcriptional functions of tumor cells are mainly regulated by protein-DNA interactions at genomic loci.
In human cells,the largest class of the DNA-binding proteins are zinc finger domain-containing proteins,which are encoded by nearly 5% of all human genes (8).Zinc finger domain-containing proteins play variable roles in maintaining genome integrity,genome stability,and transcriptional regulation (9).Based on the specific C2H2 zinc features,the zinc finger domain-containing proteins exert high binding affinity to DNA sequences with the αhelix of the finger aided by the conserved linkers present between fingers (9).ZNF292 is a growth hormonedependent zinc finger transcription factor that belongs to the kruepel C2H2-type zinc-finger protein family.Several studies have identifiedZNF292as a tumor suppressor gene(10,11).However,the exact underlying mechanism remains unelusive.
In this study,we aimed to identify the role ZNF292 plays in the occurrence and malignant development of ESCC.We found a relatively high expression of ZNF292 in adjacent and normal tissues and a low expression level in tumor tissues.Knockdown of ZNF292 resulted in high proliferation rates,elevated colony formation ability,and cell cycle progression.Mechanically,depletion of ZNF292 promoted the expression of SKP2 at the transcriptional level,thereby stimulating P27 ubiquitin-degradation,and ultimately promoting cell cycle progression.Thus,our findings signalize a novel ZNF292-SKP2-P27 transcriptional regulatory pathway,which deepens our understanding of how ZNF292 works to suppress the proliferation of ESCC cells.
Materials and methods
Cell culture and oligonucleotide transfection
Human ESCC KYSE30 and YES2 cells (kindly provided by Professor Yutaka Shimada of Kyoto University),were cultured in RPMI 1640 (Gibco) with 10% fetal bovine serum (FBS),and HEK293 human embryonic kidney epithelial cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% FBS.All the cells are maintained at 37oC with 5% CO2.A total of 15×104cells were seeded into the 6-well culture plates,and transfected with siRNA with Lipofectamine 2000 (Invitrogen)according to the manufacturer’s instructions.Specific siRNAs targeting ZNF292 and SKP2 were custom designed and provided by RiboBio company and the sequences are indicated inTable 1.

Table 1 Sequences of siRNAs and primers
Bioinformatics analysis
To analyze the gene expression of ESCC from The Cancer Genome Atlas (TCGA),we obtained the RNA-seq fragments per kilobase million (FPKM) values from the TCGA genomic data commons (GDC) portal,and then thelog2 transformed values were calculated.The survival information of 81 primary ESCC patients was obtained from University of Calfornia Santa Cruz (UCSC) Xena(https://tcga.xenahubs.net).
Reverse transcription polymerase chain reaction (PCR)and quantitative real-time PCR (qRT-PCR)
TRIzol Reagent (15596018,Invitrogen,Carlsbad,USA)was used to extract total RNA.cDNA synthesis was performed by using a reverse transcription kit (Promega).qRT-PCR was performed with a SYBR Premix Ex TaqTM II kit (TaKaRa).The mRNA levels of the target genes were normalized to β-actin.Primers were provided by Invitrogen and are indicated inTable 1.
Chromatin immunoprecipitation (ChIP) assay
ChIP assays were performed using a Pierce™ Magnetic ChIP Kit (Thermo Scientific™,Waltham,USA) according to the manufacturer’s instructions.Immunoprecipitation was performed with anti-ZNF292.Specific sequences were quantified by qRT-PCR using the primers indicated inTable 1.
Western blot analysis
Radioimmunoprecipitation assay (RIPA) buffer was used to lyse cells.Total proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE),and then transferred to polyvinylidene fluoride (PVDF)membranes.After blocking with 5% milk,membranes were incubated overnight at 4oC.Antibodies against ZNF292(1:1,000,Abcam,ab111438,Cambridge,USA),SKP2(1:500,Proteintech,15010-1-AP),P27 (1:500,Proteintech,25614-1-AP,Rosemont,USA),and β-actin (1:3,000,Santa Cruz Biotechnology,sc-58673,Dallas,USA) were purchased as noted in the brackets.The next day,after incubation with secondary antibodies,enhanced chemiluminescence detection reagent was used to visualize specific bands respectively.
Proliferation assay
The xCELLigence Real-Time Cell Analyzer (RTCA)-MP system (Acea Biosciences/Roche Applied Science) was used to test the proliferation rates of tumor cells.The culture medium (50 μL) was added to each well of an E-Plate 96(Roche Applied Science) to obtain equilibrium.Transfected cells were incubated in 6-well culture plates for 24 h,and then 2,000 cells in 100 μL culture medium were seeded in E-Plate 96.E-Plate 96 was locked in a RTCA-MP device at 37oC with 5% CO2.The cell index was read automatically every 15 min,and the recorded curve is shown as.
Colony formation assay
A total of 1,000 transfected cells were seeded into 6-well culture plates and incubated at 37oC with 5% CO2for approximately 10 d.After a wash with PBS,cells were fixed with precooled methanol for 20 min,and then stained with crystal violet for 15 min.
Flow cytometric and cell cycle analyses
A total of 15×104YES2 and KYSE30 cells were seeded into 6-well culture plates.After an appropriate time,cells were harvested,and then were fixed with 75% ethanol at -20oC overnight.The next day,the cells were washed with PBS twice,and incubated with PI solution at 37oC for 30 min.Then flow cytometry was used to analyze the cell cycle distribution.For cell cycle distribution assay after knocking down ZNF292,KYSE30 and YES2 cells treated with si-ZNF292 or si-con were maintained at 37oC with 5% CO2for 48 h,and the following steps were performed as described above.
Ethynyl deoxy uridine incorporation assay
A total of 15×104YES2 and KYSE30 cells were seeded into 6-well culture plates.After siRNA transfection for 48 h,5-ethynyl-2’-deoxyuridine (EdU) labelling was performed according to the manufacturer’s instructions provided in the BeyoClickTMEdU-488 cell proliferation assay kit(Beyotime,Shanghai,China).
Luciferase reporter assay
Genomic DNA fragments of the humanSKP2gene upstream (2,000 bp) and downstream (500 bp) from the initiation sites were inserted into the pGL3-Basic vector(denoted as pGL-SKP2).Combined with theRenillaluciferase plasmid and si-ZNF292,the luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s protocol.
Tissue microarray and immunohistochemistry (IHC)assay
An ESCC tissue microarray (CGt No:HesoS090PG01)was obtained from Shanghai Outdo Biotech Company with the approval of the Institutional Review Board.Thirty paired ESCC samples and adjacent normal tissues were collected by the National Human Genetic Resources Sharing Service Platform (2005DKA21300) in Zhejiang Taizhou Hospital.This tissue microarray contains 30 primary tumor dots,30 adjacent dots,and 30 normal dots.Patient information is shown inTable 2.Heat-mediated antigen retrieval was performed in citrate buffer,and then blocked with 3% hydrogen peroxide.After blocking,the dots were incubated with anti-ZNF292 (1:100,Abcam,ab111438) at 4 °C overnight.The next day,after incubation with secondary antibody,diaminobenzidine was used as a chromogenic substrate.Hematoxylin was used to counterstain these dots.At last,these dots were dehydrated and cover slipped.Based on the intensity,the expression of ZNF292 was visualized.Intensity of staining was scored as— means negative,+means weakly positive,++means moderately positive,and +++means strongly positive.

Table 2 Patient information
Statistical analysis
All data presented in this study were.SPSS software package (Version 17.0;SPSS Inc.,Chicago,IL,USA) was used to analyze these data.Statistical differences were assessed using two-tailed Student’sttest. In all experiments,differences were considered to be significant when P was less than 0.05.All assays were repeated at least 3 times.
Results
Low expression of ZNF292 in ESCC indicates a worse prognosis
To explore the potential role played by ZNF292 in the development of ESCC,we obtained clinical information for ZNF292 in ESCC from the TCGA database.The association of ZNF292 expression with overall survival was analyzed via Kaplan-Meier analysis.Overall survival analysis using the log-rank test indicated that a lower ZNF292 expression rate was associated with reduced overall survival in ESCC patients (P=0.028) (Figure 1A).We also analyzed the mRNA expression profile of ZNF292 in TCGA RNAseq database.The results showed that the expression of ZNF292 in adjacent normal tissues was slightly higher than that in ESCC tumor tissues,but the difference was not statistically significant (possibly due to the limitation of the low number of normal tissues) (Figure 1B).
To present the different expression profiles of ZNF292 in tumor samples and corresponding adjacent normal tissues,we performed an IHC assay in a tissue microarray within 30 primary tumor dots,30 adjacent dots,and 30 normal dots.As shown inFigure 1C,most ESCC samples exhibited lower protein expression of ZNF292 than adjacent or normal tissues.Overall,these results suggest that ZNF292 may act as a tumor suppressor in ESCC.
Depletion of ZNF292 promotes ESCC cell proliferation by promoting G1/S phase transition
To unveil the underlying mechanism of ZNF292 in depth,we first tested the expression profile of ZNF292 in ESCC cell lines and selected YES2 and KYSE30,which have relatively high expression levels of ZNF292,for subsequent experiments (Figure 2A).We knocked down ZNF292 in YES2 and KYSE30 cells to analyze the change in proliferation rates as well as the colony formation ability.The results showed that depletion of ZNF292 significantly boosted the proliferation rates and colony formation of ESCC cell lines (Figure 2B,C).Consistently,a clonogenic soft agar assay demonstrated that knockdown of ZNF292 significantly promoted the clonogenic growth of ESCC cell lines (Figure 2D,E).We also analyzed the change in cell cycle after manipulating ZNF292,and the results showed that knockdown of ZNF292 significantly accelerated S phase entry in both YES2 and KYSE30 cells (Figure 2F,G).Similarly,EdU assays demonstrated that ZNF292 deprivation significantly accelerated cell proliferation(Figure 2H,I).These data indicated that depletion of ZNF292 might boost ESCC cell proliferation by promoting cell cycle S phase entry.
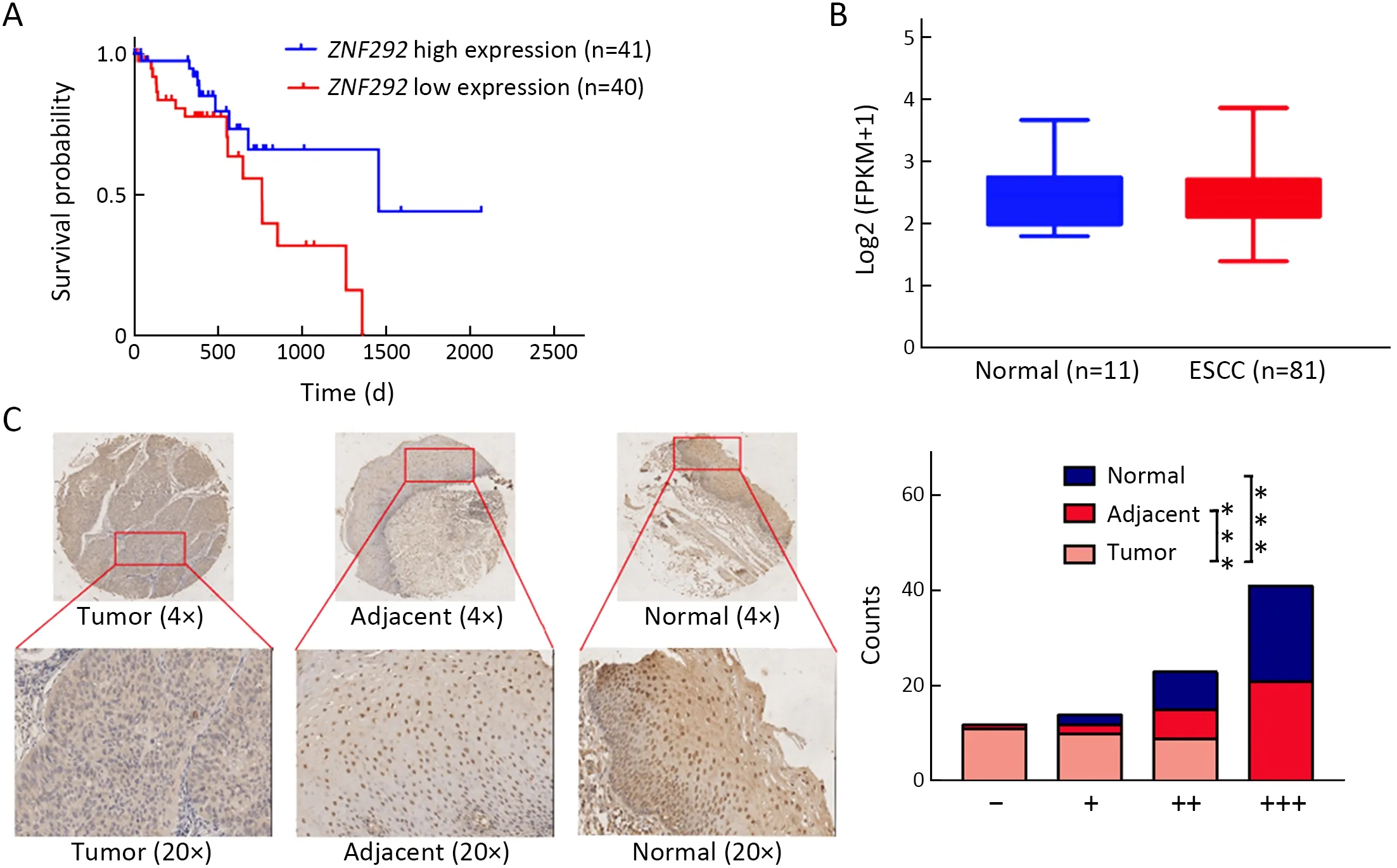
Figure 1 Low expression of ZNF292 in ESCC indicates a worse prognosis.(A) Kaplan-Meier survival analysis of ESCC patients’ overall survival based on the ZNF292 mRNA level (P=0.028);(B) mRNA level of ZNF292 in normal tissue and ESCC tissue in TCGA RNA-seq database;(C) IHC assays were performed to analyze the expression profile of ZNF292 in a tissue microarray within 30 primary tumor dots,30 adjacent dots,and 30 normal dots.ESCC,esophageal squamous cell carcinoma;TCGA,The Cancer Genome Atlas;IHC,immunohistochemistry.***,P<0.001.
Knockdown of ZNF292 promotes P27 ubiquitindegradation
To identify the mechanism of ZNF292 in cell cycle regulation,we knocked down ZNF292 and analyzed the expression of S phase entry-related proteins at the RNA and protein levels.As depicted inFigure 3A,B,knockdown of ZNF292 increased the RNA levels of both cyclins and cyclin-dependent kinases (CDKs),as well asKIP1(P27),which are related to S phase entry.At the protein level,different proteins appeared to be variable.As depicted inFigure 3C,knockdown of ZNF292 resulted in elevated expression of CDK2,CDK6,and Cyclin E,but unchanged expression of Cyclin A,Cyclin D1 and CDK4.Interestingly,we found that depletion of ZNF292 reduced the expression level of P27,which was opposite to the RNA level.In view of the central role of P27 in cell cycle G1/S phase transition and its expression pattern in cell cycle,we hypothesized that knockdown of ZNF292 would increase the P27 ubiquitin-mediated degradation,and subsequently boost S phase entry.To verify that the decreased protein expression of P27 was caused by degradation,IP assay was performed to detect the expression of ubiquitin-bound P27.As shown inFigure 3D,MG-132,which is characteristic of blocking protein degradation,significantly boosted the ubiquitination of P27 in ZNF292-knockdown cells.In addition,cycloheximide (CHX),which blocks protein translation,significantly reduced the half-life of P27 in ZNF292-knockdown cells (Figure 3E). These data suggested that knockdown of ZNF292 might result in an increase in P27 ubiquitin-mediated degradation and subsequent cell cycle S phase entry.
Knockdown of ZNF292 suppresses expression of P27 by promoting SKP2 expression
To explore the potential mechanism underlying the regulation of P27 by ZNF292,we first focused on SKP2,which has been reported to be involved in P27 ubiquitindegradation (12,13).We tested the regulatory role of SKP2 on P27 in ESCC cell lines.As shown inFigure 4A,B,knockdown of SKP2 significantly upregulated the expression of P27,indicating the direct regulation of SKP2 to P27 in ESCC cell lines.Based on this,we speculated that whether ZNF292 might regulate the expression of SKP2.Western blotting and qRT-PCR were then performed to test the expression level of SKP2 after knockdown of ZNF292.The results showed that depletion of ZNF292 significantly stimulated both the mRNA and protein levels of SKP2 (Figure 4C,D).In addition,rescue experiments demonstrated that ZNF292 knockdown accelerated the degradation of P27 by inhibiting SKP2 expression (Figure 4E),indicating the existence of a regulatory axis of ZNF292/SKP2/P27.
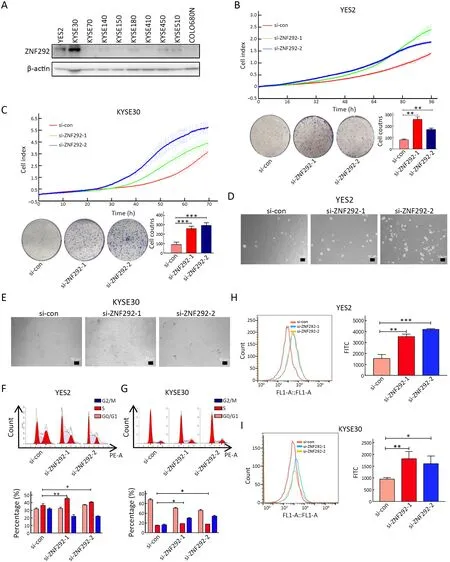
Figure 2 Depletion of ZNF292 promotes ESCC cell proliferation by promoting cell cycle G1/S phase transition.(A) Western blot analysis was used to detect the ZNF292 expression profile in ESCC cell lines;(B) Cell proliferation rates and colony formation ability analyses after knocking down ZNF292 in YES2 cells;(C) Cell proliferation rates and colony formation ability analyses after knocking down ZNF292 in KYSE30 cells;(D,E) Images of representative soft agar plates,revealing accelerated clonogenic growth of ZNF292-knockdown cells compared to control cells in YES2 (D) and KYSE30 (E) cells;(F,G) Cell cycle analysis after knocking down ZNF292 in YES2 (F) and KYSE30 (G) cells;(H,I) EdU dye assays were used to monitor cell proliferation after depletion of ZNF292 in YES2 (H) and KYSE30 (I)cells.ESCC,esophageal squamous cell carcinoma.*,P<0.05;**,P<0.01;***,P<0.001.
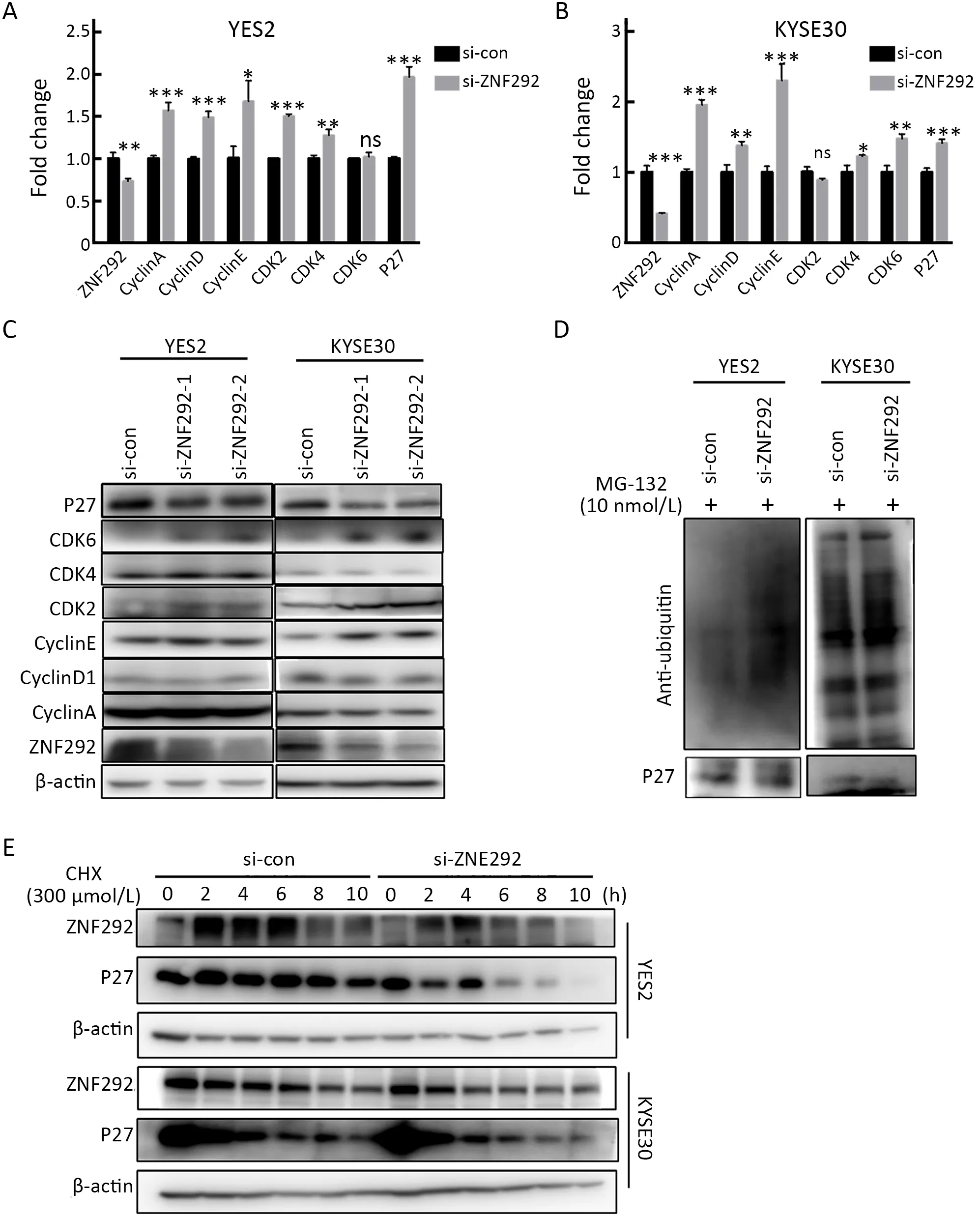
Figure 3 Knockdown of ZNF292 promotes P27 ubiquitin-degradation.(A) qRT-PCR was performed to analyze change in S phase entrance-related genes after knocking down ZNF292 in YES2 cells;(B) qRT-PCR was performed to analyze change in S phase entrancerelated genes after knocking down ZNF292 in KYSE30 cells;(C) Western blot analysis was used to detect the change in S phase entrancerelated proteins after knocking down ZNF292 in YES2 and KYSE30 cells;(D) IP assays were performed to analyze the expression of ubiquitin-bound P27 after knocking down ZNF292 in YES2 and KYSE30 cells;(E) Culture medium containing 300 μmol/L CHX was added to cells after knocking down ZNF292 for 24 h,and then cells were harvested every 2 h.Western blotting was used to detect change in P27 at different culture times.qRT-PCR,quantitative real-time polymerase chain reaction;IP,immunoprecipitation;CHX,cycloheximide;ns,not significant.*,P<0.05;**,P<0.01;***,P<0.001.
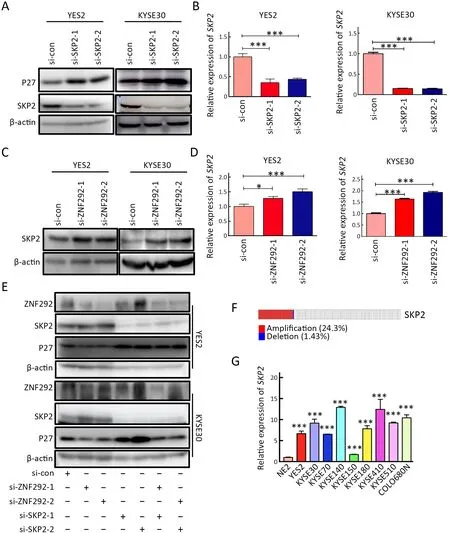
Figure 4 Knockdown of ZNF292 suppresses the expression of P27 by promoting SKP2 expression.(A) Western blot analysis was used to detect the change in P27 after knocking down SKP2 in YES2 and KYSE30 cells;(B) qRT-PCR was performed to detect the knockdown efficiency of si-SKP2 in YES2 and KYSE30 cells;(C) Western blot analysis was used to detect SKP2 expression upon the depletion of ZNF292 in YES2 and KYSE30 cells;(D) qRT-PCR was performed to detect SKP2 expression upon the depletion of ZNF292 in YES2 and KYSE30 cells;(E) A rescue experiment was performed to analyze the expression of p27.Cells were transfected with si-con (1 unit),si-ZNF292 (1 unit),si-SKP2 (1 unit),si-ZNF292 &si-SKP2 (half and half units) in six well culture plates,48 h later,cells were harvested,and western blotting was performed to analyze the expression of SKP2 and p27;(F) Genetic information analysis for SKP2 in ESCC from the TCGA database;(G) qRT-PCR was performed to detect the expression profile of SKP2 in ESCC cell lines.qRT-PCR,quantitative realtime polymerase chain reaction;ESCC,esophageal squamous cell carcinoma;TCGA,The Cancer Genome Atlas.*,P<0.05;***,P<0.001.
SKP2is an oncogene that has been considered as a strong prognostic marker for tumor development (14).We then performed genetic information analysis forSKP2in ESCC from our previous study,and found that the rate of genetic alteration ofSKP2was as high as 24.3% (15) (Figure 4F).In addition,we found that compared to the normal epithelial cell line NE2,SKP2has a remarkably high expression level in ESCC cell lines (Figure 4G),indicating the importance and necessity of SKP2 to the occurrence and malignant development of ESCC.The above results indicate that ZNF292 may inhibit the expression of SKP2,which further implies the importance of the tumor suppressor geneZNF292in suppressing the malignant development of ESCC.
ZNF292 functions to suppress SKP2 expression at transcriptional level
To further confirm the correlation between ZNF292 and SKP2,transcription factor binding prediction (http://cistrome.org) of ZNF292 was performed for its transcription factor characteristics.The results showed that there was a signal at the promoter element ofSKP2(Figure 5A).Furthermore,we applied ChIP assays and confirmed a high binding affinity of ZNF292 in theSKP2promoter region (Figure 5B).To further examine the transcriptional activity of ZNF292 onSKP2,we constructed a luciferase reporter assay system (Figure 5C).We found that the luciferase activity increased significantly upon depletion of ZNF292 in both HEK293 cells and KYSE30 cells (Figure 5D,E).Taken together,these results indicate that ZNF292 can inhibit SKP2 expression by binding directly to the promoter ofSKP2and ultimately lead to P27 accumulation to suppress the proliferation of ESCC cells.
Discussion
In this study,our findings are suggestive of a novel mechanism by which ZNF292 may exert a negative role on ESCC cell cycle progression in ESCC (Figure 6).We demonstrated the regulatory role of ZNF292 onSKP2,which conduces to explaining how ZNF292 inhibits ESCC cell proliferation. Mechanistically,ZNF292,as a transcription factor,suppresses the expression ofSKP2by binding to the promoter element ofSKP2.Consequently,decreased SKP2 attenuates the ubiquitin-mediated degradation of P27,and eventually,the accumulated P27 suppresses the cell proliferation of ESCC cells by blocking cell cycle progression.
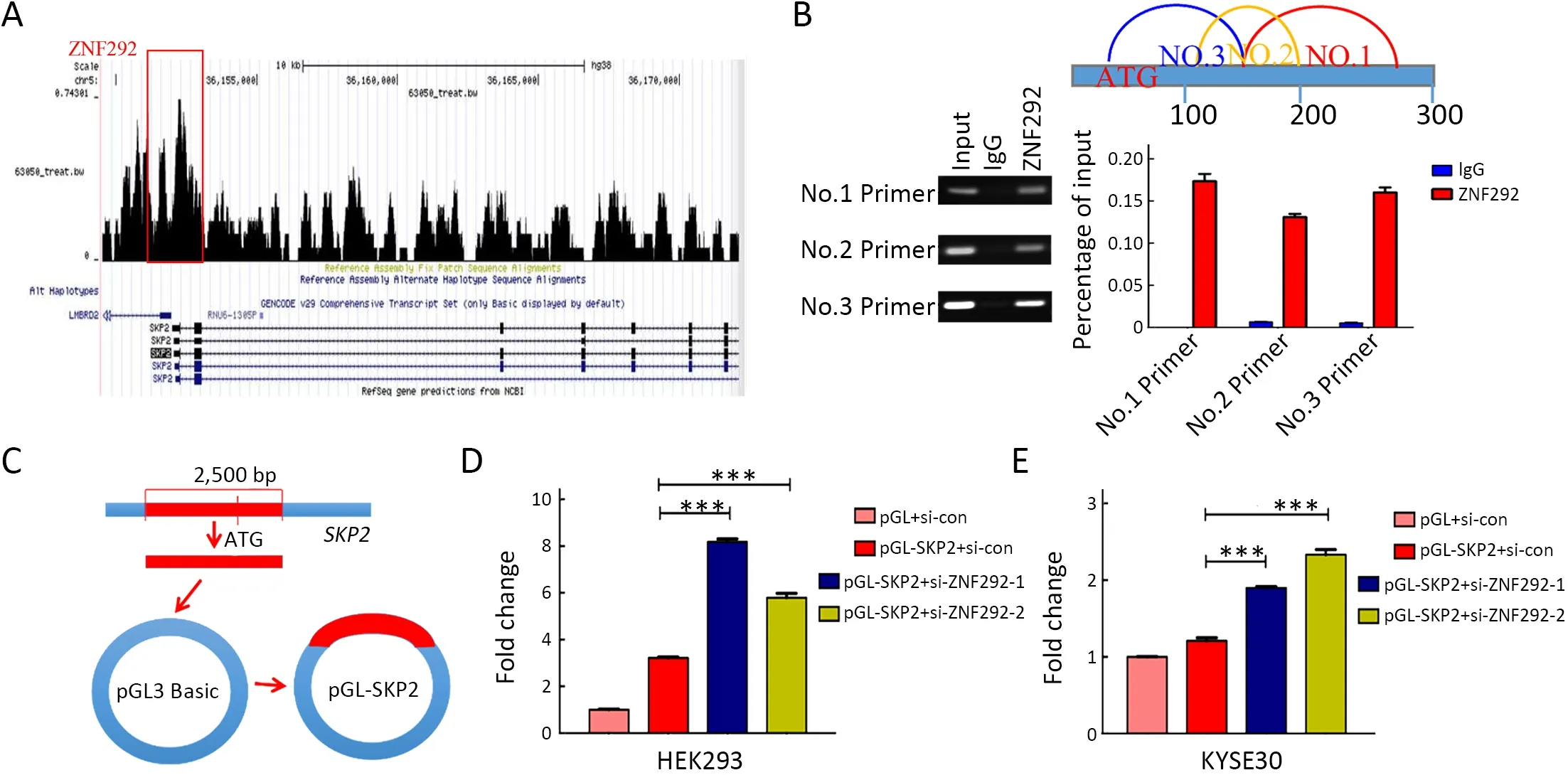
Figure 5 ZNF292 functions to suppress SKP2 expression at transcriptional level.(A) Transcription factor binding prediction for ZNF292 on the promoter element of SKP2;(B) ChIP assay for the binding ability of ZNF292 to the promoter element of SKP2 in KYSE30 cells;(C)Construction model of the luciferase reporter plasmids;(D,E) Luciferase reporter results analyses in HEK293 cells (D) and KYSE30 cells(E).These two cell lines were all transfected with pGL+si-con,pGL-SKP2+si-con,pGL-SKP2+si-ZNF292-1,or pGL-SKP2+si-ZNF292-2.After transfection for 48 h,cells were subjected to dual luciferase analysis.The results were presented as the fold change relative to cells transfected with pGL+si-con after normalization to Renilla activity.ChIP,chromatin immunoprecipitation;IgG,immunoglobulin G.***,P<0.001.
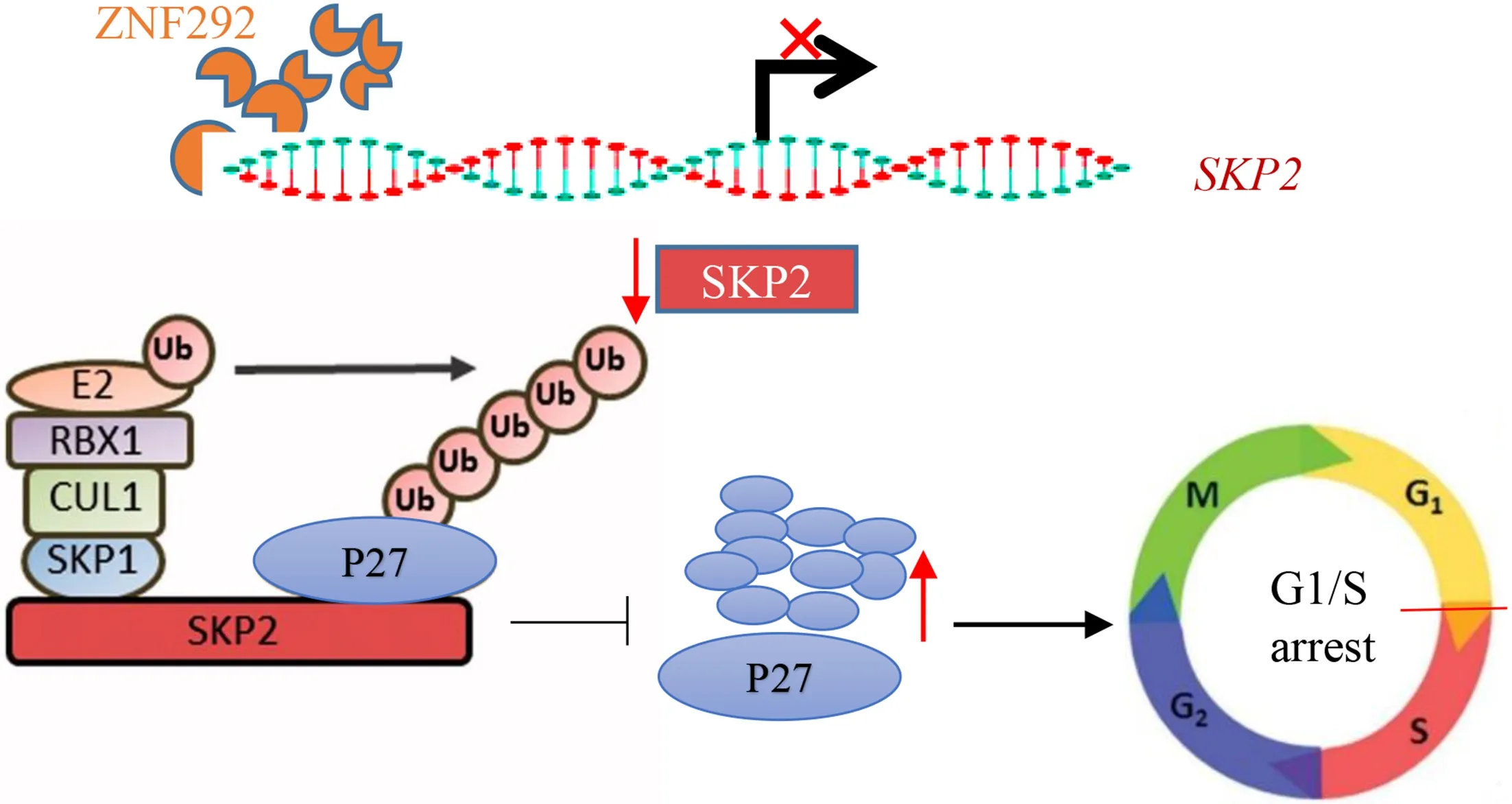
Figure 6 Graphical summary of ZNF292/SKP2/P27 pathway.ZNF292 functions to suppress the expression of SKP2 at the transcription level.The decrease of SKP2 attenuates the ubiquitin-mediated degradation of P27.As a result,the accumulated P27 suppresses cell proliferation of ESCC cells by blocking cell cycle progression.ESCC,esophageal squamous cell carcinoma.
Cell cycle progression is based on the activation of CDKs (16) and the blockage of cyclin-dependent kinase inhibitors (12).Abnormal expression of cell cycle regulators is conductive to malignant progression of tumor cells.Among these regulators,the tumor suppressor protein P27 can be marketed out as being involved in cell proliferation,differentiation,apoptosis,cellular adhesion and growth inhibition (12,17).Previous studies indicated that the absence of P27 expression is associated with a poor prognosis in a variety of cancer types (18,19).P27 functions to block the transition from G1 to S phase of the cell cycle to inhibit cell growth and proliferation.Unlike other tumor suppressor proteins,such as P53 and P16,the decreased expression of P27 in tumor cells is mainly due to the increase in protein degradation rather than genetic mutations that interfere with mRNA synthesis (18-20).The degradation of P27 mainly depends on its phosphorylation,which allows SKP2 binding and subsequent proteasomedependent degradation.
SKP2is an oncogene and has been considered as a strong prognostic marker for tumor development.The high copy number amplification rate (24.3%) in ESCC patients indicates the pivotal role ofSKP2in the occurrence and development of ESCC.SKP2 is involved in the ubiquitinmediated degradation of P27,while decreased P27 and increased SKP2 are correlated with tumor size,stage,grade and metastasis (21-23).Several studies have demonstrated that the expression of SKP2 can be regulated through different pathways,such as PI3K,ERK (24,25),Akt (26),and MAPK (27).Additionally,the von Hippel-Lindau(VHL) protein can destabilize SKP2 by regulating the ubiquitin ligase activity of SKP2 through its β-domain (28).Here,we found a transcriptional regulatory axis ofSKP2that was executed by ZNF292.
ZNF292 is a growth hormone-dependent zinc finger transcription factor that belongs to the kruepel C2H2-type zinc finger protein family.Several studies have identifiedZNF292as a tumor suppressor gene in many cancer types.Gene mutation of tumor suppressor genes may attenuate their tumor suppressor function.ZNF292mutations have been identified in cancers of liver,colon,bone marrow and stomach (10).In our previous study,we also found mutations ofZNF292in ESCC.However,ZNF292mutations have been observed in many cancers with low to moderate incidences (1.3% in colorectal cancer,4.4% in gastric cancers,and 5.56% in ESCC) (15,29),indicating thatZNF292may be a moderately conserved gene expressed in a variety of human tissues.Therefore,more studies should be incorporated to study the multiple functions of wild-type ZNF292 to develop effective targeted therapeutic drugs against ESCC and even other cancer types.The cell cycle-regulatory illusion of ZNF292 helps to enrich the functions of zinc finger protein family members.
Conclusions
We confirmed a transcriptional regulatory pathway of ZNF292/SKP2,which deepens our understanding of the underlying mechanism of the tumor suppressor geneZNF292.Additionally,these results also corroborate the important role of ZNF292/SKP2/P27 signaling in cell cycle regulation in ESCC.
Acknowledgements
This work is supported by the National Natural Fund of China (No.81988101,81830086 and 81972318) and the Doctoral Innovation Fund of Peking Union Medical College (No.2018071011).
Footnote
Conflicts of Interest:The authors have no conflicts of interest to declare.
杂志排行

Chinese Journal of Cancer Research的其它文章
- Prudent application of single-cell RNA sequencing in understanding cellular features and functional phenotypes in cancer studies
- Development and validation of a questionnaire-based risk scoring system to identify individuals at high risk for gastric cancer inChinese populations
- Associations of centralization with health care quality for gastric cancer patients receiving gastrectomy in China
- Identification of lymph node metastasis by computed tomography in early gastric cancer
- Deep learning model improves radiologists’ performance in detection and classification of breast lesions
- All-trans retinoic acid (ATRA) inhibits insufficient radiofrequency ablation (IRFA)-induced enrichment of tumor-initiating cells in hepatocellular carcinoma