山东田横岛海域长牡蛎性腺发育及生化成分周年变化❋
2022-01-13吕敏,李琪,2❋❋
吕 敏, 李 琪,2❋❋
(1.海水养殖教育部重点实验室(中国海洋大学), 山东 青岛 266003; 2. 青岛海洋科学与技术国家试点实验室 海洋渔业科学与食物产出过程功能实验室, 山东 青岛 266237)
海洋双壳贝类性腺发育与环境因子(温度、食物)及内在生理代谢密切相关。温度调控配子发生的时间和速度,食物丰歉调控配子发生的强度,配子重吸收循环是性腺发育额外营养来源[1]。海洋双壳贝类能量储存和生化成分的季节性变化与生殖活动密切相关[2]。糖原、蛋白质、脂肪是软体动物体内的主要能源供应形式。不同物种配子发生期间所消耗能源类型存在差异,Liu等[3]研究发现滑顶薄壳鸟蛤(Fulviamutica)配子发生期的能源是脂质;Kang等[4]研究发现韩国海湾扇贝配子发生期的能源是糖原;Thompson[5]对巨扇贝(Placopectenmagellanicus)研究发现其配子发生期的能源来自近期摄入的食物和之前的储备。根据双壳贝类配子发生动员的能量类型不同,Bayne[6]将其总结为两种不同繁殖策略:(1)配子发生利用之前储存于各组织中能量的物种称为保守种,如褶牡蛎(Crassostreaplicatula)、中国蛤蜊(Mactrachinensis);(2)配子发生利用最近摄入的食物作为能量的物种称为机会种,如坚固马坷蛤(Spisulasolida)[7-9]。
长牡蛎(Crassostreagigas)又称太平洋牡蛎,是我国主要的养殖经济贝类。随着养殖面积与养殖密度的扩大,养殖环境恶化、种质退化、死亡率上升等问题逐年增加,对养殖海区长牡蛎生长与繁殖发育产生较大影响。Li等[7-8,10]报道了日本宫城长牡蛎、山东乳山沿海中国蛤蜊和褶牡蛎性腺发育周期及生化成分季节变化,柯巧珍等[11]报道了山东北部沿海四角蛤蜊性腺发育年周期,栖息海域的不同往往引起双壳贝类繁殖习性发生变化[12]。田横岛位于山东省青岛市即墨东部海域,自然条件良好,长牡蛎等贝类资源较为丰富。然而有关其生殖周期与海区环境因子关系的研究,目前还没有报道。本研究调查田横岛海域长牡蛎生殖发育周期与环境因子、各组织生化成分周年变化的关系,旨在为保护田横岛海域长牡蛎种质资源,开展人工养殖提供重要的参考资料。
1 材料与方法
1.1 样品采集
调查样品采于青岛市田横岛牡蛎养殖海区,自2019年4月—2020年3月,每月中旬采集长牡蛎40~60个体,使用多参数水质分析仪(YSI proplus)现场测定水温(WT)和盐度(S)。同时,取表层水样固定,测定叶绿素a(Chla)浓度,其测定方法参照《海洋监测规范 GB 17378.4—2007》[13]。长牡蛎样品活体运回实验室后,每3个个体为一组,解剖获取外套膜、鳃、闭壳肌及性腺-内脏团组织,-80 ℃冷冻保存。
1.2 组织学分析
每月取长牡蛎30个个体,鉴别雌雄及发育状况,解剖取厚度为5 mm左右的性腺组织,置于 Bouin氏液固定24 h后,70%酒精梯度脱水,石蜡渗透和包埋,组织切片,爱氏苏木精-曙红(HE)法染色,在显微镜(Olympus BX53)下辨别雌雄和性腺发育状况。每月随机选取10张雌性性腺切片,每张切片测量50个卵母细胞的直径(穿过核仁的最长轴),计算平均值以确定性腺发育程度[14]。
1.3 生化成分测定
将外套膜、鳃、闭壳肌及性腺-内脏团4种组织冷冻干燥48 h,研磨成粉末状,80目筛绢过滤后,储存于干燥器中。采用Wang等[15]构建的近红外(NIR)(Antaris MX, Thermo Fisher)分析模型对各组织中的糖原、蛋白质及脂肪含量进行测定。
1.4 条件指数
每月取10个长牡蛎个体,将软体部及贝壳在80 ℃烘箱内烘至恒重,以平均软体部干质量(g)/平均贝壳干质量(g)计算每月条件指数(CI),确定性腺发育程度[16]。
1.5 数据处理
数据统计分析采用SPSS 23.0软件处理。采用单因素方差分析(one-way ANOVA)比较各月卵径、条件指数及各组织生化成分含量的差异显著性(P<0.05)。
2 结果
2.1 调查海区环境因子季节变化
青岛市田横岛海区水温具有明显的季节变化,夏季8月温度最高27.3 ℃,随后温度逐月下降,冬季1月温度降至最低5.4 ℃。周年调查海区盐度无明显季节变化,变化幅度介于30.48~32.28。调查海区属于温带季风性大陆气候,夏季高温多雨,盐度在夏季降至全年最低。叶绿素a浓度表现为春秋季节高于夏冬季节,上下半年各有一次高峰,9、10月叶绿素a浓度最高,9月为周年最高峰3.10 μg·L-1,随后逐渐下降,在冬季1月降至最低(1.10 μg·L-1)。春季叶绿素a浓度逐渐升高,在4 月出现次高峰(2.20 μg·L-1)。
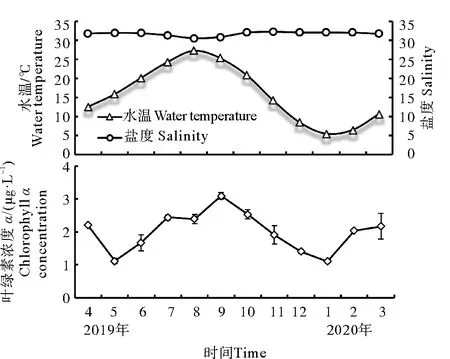
图1 调查海区温度、盐度、叶绿素a浓度周年变化Fig. 1 Seasonal variation of water temperature, salinity and chlorophyll a concentration in the survey area of Tianheng Island
2.2 调查海区长牡蛎性腺发育阶段特征
通过对调查海区长牡蛎性腺外部形态及组织切片的观察,性腺发育周期共分为六个阶段:形成期、增殖期、成熟期、排放期、耗尽期和休止期,各阶段特点如图2所示。
形成期:外观长牡蛎性腺较薄,表面可见内脏团组织。切片可辨雌雄,滤泡较小,结缔组织较多,雌性滤泡壁上卵原细胞开始发育;雄性滤泡壁上有许多精原细胞。
增殖期:外观长牡蛎性腺逐渐增厚,结缔组织减少。雌性卵巢中滤泡变大,滤泡壁上附着大量的卵母细胞,泡内有空隙及少数成熟卵子游离;雄性精巢内滤泡内空隙逐渐缩小,游离精子增多。
陕西关中冬季往往干旱寒冷,梨黑星病菌主要以菌丝和分生孢子在病梢、芽鳞和病枝上,或残存在落叶上越冬。在冬季温暖潮湿年份,分生孢子易丧失生活力而不能越冬,但病菌转而在落叶上形成菌丝团(12月)及子囊壳(1—2月)越冬,翌春再产生子囊孢子进行初次侵染。由于病菌能否形成子囊壳主要决定于冬季的湿度,因此在不同年份、不同地区,甚至同一地区不同小环境中,均可有不同的越冬方式。
成熟期:性腺肥厚饱满,软体部被性腺包裹,几乎没有结缔组织,滤泡间几乎没有间隙。雌性卵巢中滤泡内有大量游离成熟的卵子,呈现出不规则的多边形,少数卵母细胞附着在滤泡壁上;雄性精巢内滤泡中大量成熟的精子密集呈辐射状。
排放期:性腺开始萎缩,表面有明显的生殖导管,结缔组织增多,滤泡缩小,滤泡间间距加大。雌性卵巢中滤泡破裂,大量卵子排出,泡内成熟的卵子数目减少;雄性精巢中滤泡内辐射状精子成流动状向外排出。
耗尽期:性腺变薄呈透明状,结缔组织增多,该阶段属于性腺重吸收阶段,滤泡内仅有少量残留卵子和精子。
休止期:无法辨别雌雄,性腺中充满结缔组织。

(A1、A2:形成期;B1、B2:增殖期;C1、C2:成熟期;D1、D2:排放期;E1、E2:耗尽期。CT:结缔组织;FW:滤泡壁;OO:卵原细胞;DO:未成熟卵子;MO:成熟卵子;RO:残留卵子;SN:精原细胞;SP:精子;RS:残留精子;标尺:50 μm。A1, A2: Early active stage; B1, B2: Development stage; C1, C2: Ripe stage; D1, D2: Spawning stage; E1, E2: Spent stage. CT: Connective tissue; FW: Follicle wall; OO: Oogonia; DO: Developing oocyte; MO: Mature oocyte; RO: Residual oocyte; SN: Spermatogonia; SP: Sperm; RS: Residual sperm; bar: 50 μm.)
2.3 性腺发育周期的周年变化
图3显示调查海区长牡蛎性腺发育周期所占比例的周年变化。1月进入形成期的雌雄个数分别占22.2%和9.0%。3月长牡蛎全部处于形成期。4月绝大部分长牡蛎处于增殖期,少部分个体进入成熟期。5月长牡蛎处于成熟期,6月部分个体开始排放配子(雌性17.4 %,雄性33.3 %),配子排放集中在7—8月。随着配子排放,9月进入耗尽期的个体数量逐渐增加,10—12月绝大多数长牡蛎处于休止期。
2.4 长牡蛎卵径与条件指数季节变化
如图4所示,卵径从1月开始逐渐增大,在5月进入成熟期之后卵径达到最大值(51.38 μm),随着配子排放卵径逐渐缩小,各月卵径差异性显著(P< 0.05)。
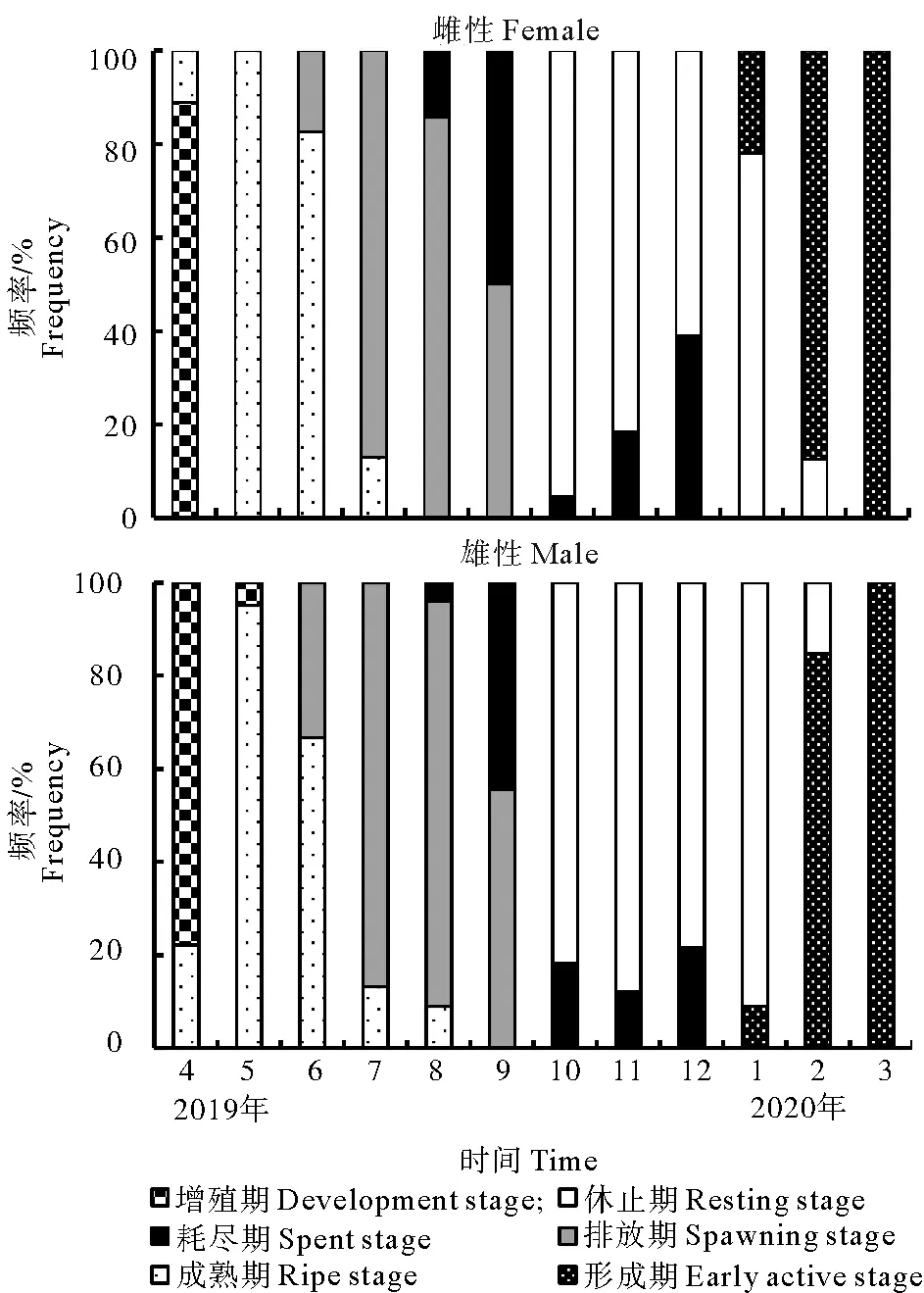
图3 调查海区长牡蛎性腺发育周期的周年变化Fig. 3 Seasonal variation of gonad development cycle of C. gigas in the survey area of Tianheng Island
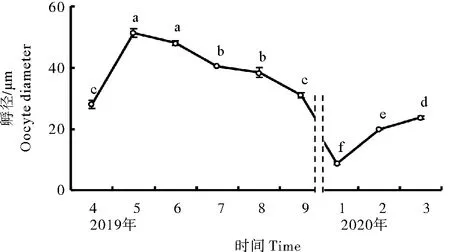
图4 调查海区长牡蛎卵径季节变化Fig.4 Seasonal variation of oocyte diameter of C. gigas in the survey area of Tianheng Island
如图5所示,条件指数在5月长牡蛎进入成熟期后达到最大6.6%,随着配子排放,条件指数逐渐减小,在9月降到最低3.7%。当长牡蛎性腺处于休止期时,条件指数各月份无显著差异。性腺发育周期重新开始时,条件指数逐步增加,各发育阶段有显著差异(P<0.05)。
2.5 各组织营养成分周年变化

图5 调查海区长牡蛎条件指数季节变化Fig. 5 Seasonal variation of condition index of C. gigas in the survey area of Tianheng Island
闭壳肌(74.27%±2.14%)蛋白质含量高于外套膜(48.18%±4.22 %)、鳃(56.55%±3.87%)和性腺-内脏团(48.74%±4.96%),闭壳肌蛋白质含量各月份变化幅度不显著。在配子发生期间,4—7月性腺-内脏团和外套膜蛋白质含量显著升高;配子排出后,蛋白质含量显著下降。各组织在8—10月缓慢增加,进入冬季后蛋白质含量逐渐下降,但差异不显著(见图7)。
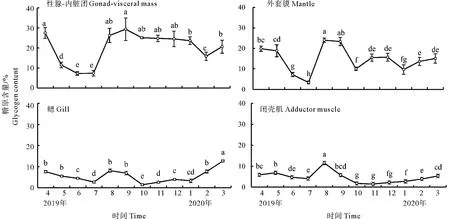
(字母不同表示月间差异显著(P < 0.05)。Different letters denote significant difference between different month (P < 0.05).)

(字母不同表示月间差异显著(P < 0.05)。Different letters denote significant difference between different month (P < 0.05).)
性腺-内脏团(8.67%±2.33%)的脂肪含量高于闭壳肌(2.55%±1.05%)、外套膜(4.15%±0.92%)和鳃(3.53%±1.42%)。闭壳肌和性腺-内脏团变化趋势一致,在配子发生时(1月),脂肪含量逐渐上升,在5月性腺成熟期达到最大;随着配子排放,脂肪含量下降,进入休止期后含量逐渐上升。外套膜和鳃脂肪含量周年变化平缓,各月份无显著差异(见图8)。
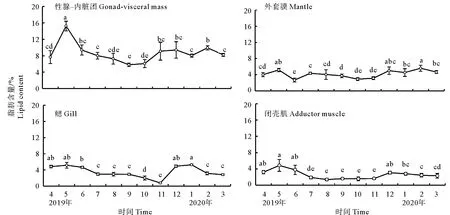
(字母不同表示月间差异显著(P<0.05)。Different letters denote significant difference between different month (P < 0.05).)
3 讨论
3.1 长牡蛎性腺发育周期与环境因子关系
双壳贝类繁殖周期与内源因素(营养和激素)和外源因素(水温、盐度、食物)密切相关[6, 17]。配子发生周期及产卵持续时间受地理纬度特别是水温的影响显著[18]。配子形成是缓慢过程,水温被认为是决定贝类配子发育和产卵最重要的环境因素。一方面温度能有效控制生殖细胞的有丝分裂,控制性腺发育程度;另一方面温度升高能提高浮游植物丰度,为繁殖期间双壳贝类提供能量来源[2,14,19]。Mann[20]研究表明双壳贝类的繁殖发育有相应的生物学零度和诱导产卵的最低绝对温度,而且同一物种在不同地区的配子发育和产卵温度不同[21]。Li等[10]研究发现日本女川湾的长牡蛎配子开始发育温度为8~10 ℃。本研究结果显示,调查海区长牡蛎少部分个体的配子自1月开始发育,此时海区温度为5.4 ℃。Drummond等[22]研究发现短暂的升温刺激有可能促进配子的发生。长牡蛎诱导产卵的最低绝对温度为18 ℃[20],调查区17.4%雌性牡蛎配子于6月开始排放,此时水温为20.0 ℃。
温度影响双壳贝类的性腺发育,饵料丰度影响其繁殖力与配子质量[19]。浮游植物是海洋的初级生产者,是海洋贝类的主要食物来源,叶绿素a相对浓度高低反映水体浮游植物丰度[23-24]。Ruiz等[25]研究表明西班牙海域长牡蛎第二个产卵高峰发生在浮游植物丰度最大、温度较低的10月,由此可见温度并不是刺激牡蛎产卵的唯一环境因素,浮游植物丰度也会刺激双壳贝类产卵。Starr等[26]研究表明充足的浮游植物可以刺激海胆(Strongylocentrotusdroebachiensis)和紫贻贝(Mytilusedulis)产卵。刘文广等[27]研究表明饥饿状态下长牡蛎存在发育迟缓、配子不排等现象。本研究测定叶绿素a浓度周年变化,通过组织学观察该海区长牡蛎配子发生在温度较低、饵料含量相对丰富的春季。这与滑顶薄壳鸟蛤(Fulviamutica)和魁蚶(Scapharcabroughtonii)的配子发育起始时间相似[3,28]。随着温度不断升高,成熟配子集中在7—8月排放,此时叶绿素a含量达到高峰,为长牡蛎幼体的发育提供充足的食物供应,以维持幼体的存活。
条件指数反映牡蛎性腺成熟程度,因为配子产生和释放在组织重量上表现出增加或减少[29]。Kim等[30]研究指出条件指数会随着性腺发育程度变化,性腺成熟过程中数值逐渐增大,排放后数值逐渐下降。本研究结果显示,配子发育初期条件指数不断上升,性腺成熟时达到最大值,随着配子排出,条件指数逐渐下降,在休止期处于平稳状态。卵径变化也是判断配子发生的一种方法,能反映牡蛎性腺发育程度[1],调查海区长牡蛎在卵子成熟过程中卵径不断增大,排卵后卵径逐渐缩小。条件指数与卵径变化相结合可以更准确的判断长牡蛎性腺的发育程度。
3.2 长牡蛎各组织生化成分的季节变化
糖原在双壳贝类配子发生的能量代谢过程中起重要作用,其含量的储存与消耗变化与生殖活动密切相关[20,31]。Berthelin等[32]研究法国西海岸长牡蛎与糖原储存相关的囊泡细胞,在配子发生前具有较高的储存能力,随着配子发生储存能力逐渐下降。本研究显示,配子发育成熟前各组织糖原含量逐渐增加,4月达到最大值,随着配子发育成熟,各组织糖原含量下降,7月降至最低,8—10月显著上升。其原因可能包括:夏末秋初食物资源丰富,从外界摄入补充体内糖原的消耗;8—10月长牡蛎性腺发育阶段属于排放后期与耗尽期,性腺中未排出配子开始重新吸收,配子重新吸收是额外的营养补充[1]。春季浮游植物生物量升高,食物充沛,各组织糖原大量积累,为繁殖期做准备[2,30]。在配子发生期间,调查海区长牡蛎主要利用储藏在各组织中糖原作为配子发生的能源。研究结果显示,性腺-内脏团和外套膜糖原含量远高于闭壳肌和鳃,推测性腺-内脏团和外套膜是糖原储存的主要组织。
在海洋双壳贝类中脂质含量能准确反映性腺发育过程[7,33]。性腺-内脏团中脂质含量变化与卵径变化一致。性腺的发育涉及糖原向脂质的代谢过程[34-35]。在配子发生过程中,糖原含量逐渐降低,脂质含量逐渐增加,主要原因是糖原代谢分解过程中会产生甘油三酯,而甘油三酯参与卵母细胞中卵黄脂的合成,而在雄性精子形成过程中脂质(磷脂)是参与细胞膜形成的主要物质[10],所以在性成熟过程中性腺-内脏团脂质含量逐渐增加,随着配子排放,性腺萎缩,脂质含量逐渐降低。外套膜、内脏团和鳃脂质含量远低于性腺-内脏团,且周年稳定无显著差异,表明它们不是贮藏脂质的组织。
蛋白质是卵母细胞的重要组成成分,与脂质相结合形成卵黄蛋白,约占卵细胞的40%~50%[36]。本研究表明,在配子成熟过程中,蛋白质含量在卵巢中不断累积,随着卵子的排放,含量显著下降。蛋白质是主要的能源物质,在配子发生过程中,卵巢中蛋白质含量增加,精巢情况则相反,在糖原含量极低的状况下,蛋白质会为精子产生提供能量[35]。在配子发生结束后,8—10月各组织蛋白质含量显著增加。11月至次年1月各组织蛋白含量持续下降,可能由于季节性食物匮乏,长牡蛎为保证机体正常代谢,蛋白质成为主要利用的能源物质[4]。闭壳肌的蛋白质含量高于其他三个组织,且周年变化相对平稳,表明在长牡蛎繁殖期间闭壳肌的蛋白质可能没有为配子发育提供能量,这与褶牡蛎[7]、魁蚶[28]、缢蛏[37]研究结果相类似。
4 结语
水温和叶绿素a浓度(饵料丰度)是影响田横岛海域长牡蛎性腺发育的主要环境因子。少部分个体在冬季(1月)开始发育,随着温度和叶绿素a浓度升高,6月部分个体进入排放期,在温度较低的10—12月性腺处于休止期。条件指数与卵径相结合可更准确地判断该海区长牡蛎性腺的发育程度。研究结果表明,该海区长牡蛎各组织生化成分周年变化与性腺发育密切相关,储存在各组织中的糖原是性腺发育的主要能量来源;性腺-内脏团中脂质和蛋白质与配子发育密切相关;在饵料相对匮乏季节,蛋白质是维持机体正常代谢的能源物质。本研究探究了田横岛海域长牡蛎繁殖发育周期与环境因子、各组织生化成分周年变化的关系,为保护田横岛海域长牡蛎种质资源,开展人工养殖提供重要的参考资料。
