贵州草海湿地不同植物群落表层土壤氮素的赋存特征
2022-01-11吴云杰张明意潘青春苏春花
王 硕, 吴云杰,2*, 张明意, 潘青春, 田 鑫, 苏春花
(1.贵州民族大学生态环境工程学院, 贵阳 550025; 2.贵州民族大学喀斯特湿地生态研究中心, 贵阳 550025)
在湿地生态系统氮循环过程中,氮素含量高低直接影响湿地土壤的养分供给状况及可利用水平[1]。植物群落表层土壤是总碳、总氮等营养元素较为丰富的土层[2],但可被植物直接吸收利用的氮素不足土壤总氮的2%[3],95%以上氮素以不能直接被植物吸收利用的有机氮形式存在[4],需要经过微生物的矿化作用将其转化为铵态氮和硝态氮形式的有效氮[5]。植物对不同形态氮素具有选择性吸收的特征,高原湿地生态系统中,植被对无机氮均能吸收,但由于氮素形态及时间空间分布特征的差异导致吸收效率有所不同[6],同时不同形态氮素对湿地生态系统碳循环过程有着重要作用,氮输入可能会影响其固碳潜势,进而减弱湿地生态系统的碳汇功能[7]。除此之外,不同形态氮素含量会对湿地土壤pH[8]、土壤硝化和反硝化[9]、功能性微生物群落[1]及生物活性[10]产生影响。土壤阳离子交换量(cation exchange capacity,CEC)是体现土壤肥力的重要指标之一,能直接反映土壤保蓄、供应和缓冲阳离子养分的能力[11],CEC的空间分布具有自相关性,与土壤pH、有机质、黏粒和电导率在不同滞后距离上存在不同的空间相互关系[12]。同时,张莹等[13]认为会仙岩溶湿地系统中CEC是土壤有机碳与总氮积累的控制因子之一,且土壤CEC与总氮呈极显著正相关[14]。吴统贵[15]在杭州湾滨海湿地植被群落演替研究中指出土壤含盐量是影响植被群落分布的主要环境因子之一,并发现4种盐基阳离子(K+、Ca2+、Na+、Mg2+)对该区域植被群落分布丰富度指数、综合多样性指数均呈现逐渐降低的过程。湿地植被演替特征和土壤氮素循环的关系是生态学研究的重要领域之一,目前研究多为针对湿地水生植物多样性及群落演替[16]、运用土壤生态化学计量学对湿地不同演替阶段植物特征的研究[17],还包括典型高原喀斯特湿地植被土壤各碳氮含量空间分布特征[18-19]及微生物活性[20]的研究,不同土地利用方式对典型喀斯特地区土壤团聚体分布及团聚体碳氮含量的探究[21]等,但针对高原喀斯特地区自然演替条件下植物群落-土壤微环境氮素含量的分布特征及其原因的研究鲜有报道。
现以贵州高原湿地-草海湿地自然演替下湿地植物群落表层土壤作为研究对象,探讨4种典型湿地植物群落表层土壤氮素含量以及不同粒径土壤团聚体中湿地表层土壤氮素的空间分布特征。以助于揭示该区典型自然演替下湿地植被空间扩展过程中植被对氮素竞争的养分机制。同时由于湿地植物群落土壤电荷和营养元素分配与植物群落自然演替之间关系的研究报道较少,可望为研究草海湿地植物群落自然演替和反馈机制提供新的研究方向。
1 材料与方法
1.1 研究区概况
贵州草海国家级自然保护区位于贵州省威宁县城西部 (26°47′32′′~26°52′52′′N,104°10′16′′~104°20′40′′ E),毗邻云贵高原乌蒙山,是长江水系金沙江支流横江洛泽河的源头湿地湖泊,也是一个完整、典型的高原喀斯特湿地生态系统[22]。草海国家级自然保护区总面积9 600 km2,属于贵州高原上最大的天然淡水湖泊,海拔约为2 171.7 m,属于亚热带湿润季风气候,年均降雨量为950 mm,其中 88% 集中在5—10月,无霜期 208.6 d,年日照时数约 1 805.4 h,挺水植物群落多为李氏禾、芦苇和水葱等群落[16],岸边多为一年生草本植物,以香薷、牛筋草等为典型。作为中国典型高原喀斯特地区湿地生态系统,草海自然保护区以其独特的生态性质、地理位置与人文景观具有突出的代表意义[23]。
1.2 供试材料
2018年8月,沿草海湿地设置12个采样点,分别选择牛筋草(NJC)、李氏禾(LSH)、香薷(XR)和芦苇(LW)4种典型植物群落各3处,具体如图1所示,每个采样点设置一个10 m×10 m植物群落样方,每个样方内用五点法取深度0~20 cm土壤,随后将所取土样放入密封袋并混合均匀,低温保存备用,用来测定氮素指标。采集样品经过湿筛法获得4种粒径团聚体粒径,标记后放在阴凉干燥处储存备用。
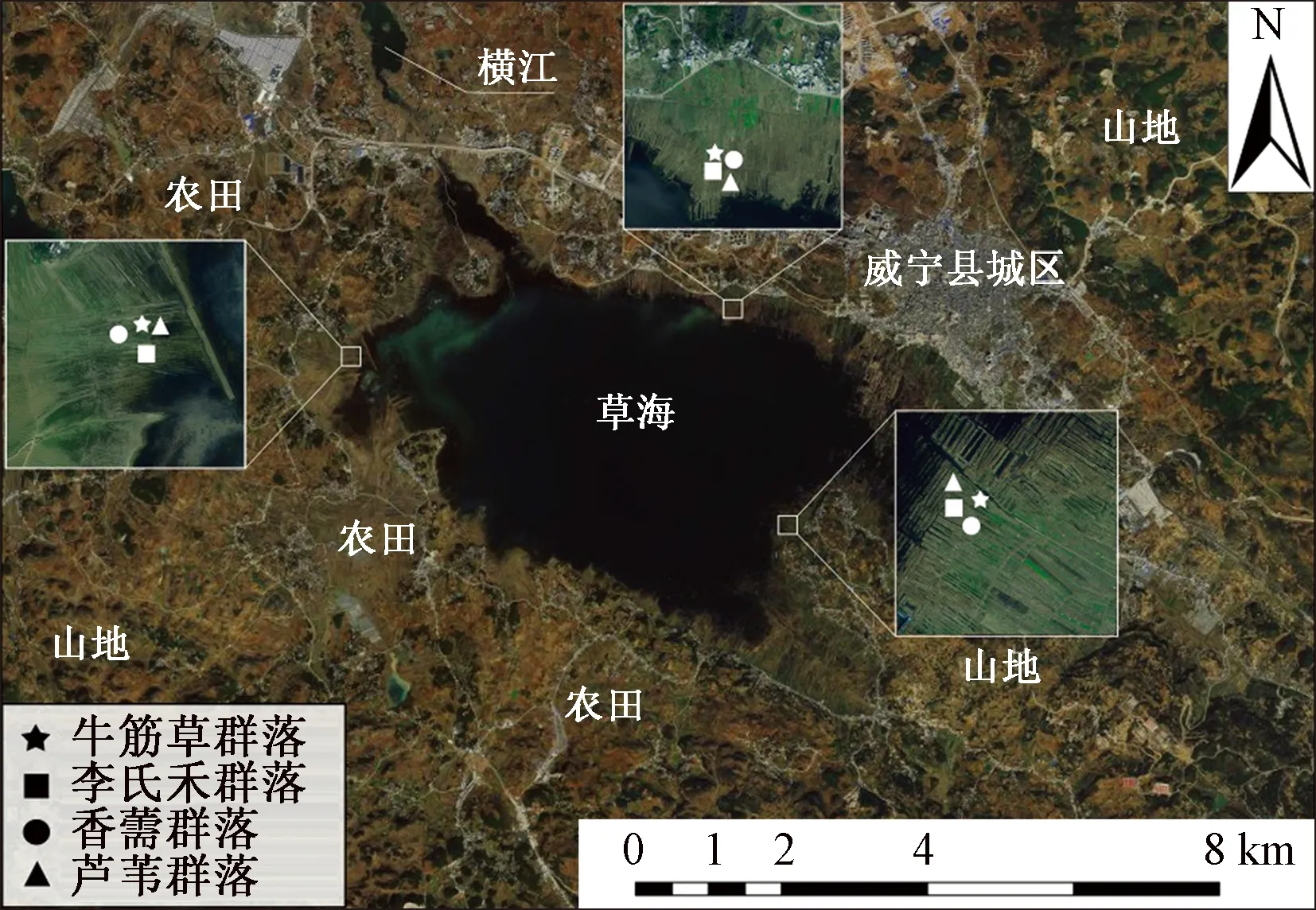
图1 草海湿地取样点示意图Fig.1 Schematic diagram of sampling points in Caohai Wetland
1.3 研究方法
研究采用氯化钡缓冲液法测定土壤CEC[24]。硝态氮测定方法为紫外分光光度法;铵态氮测定方法为2 mol/L氯化钾-靛酚蓝比色法;速效氮测定方法为碱解扩散法;总氮测定方法为半微量开式法, 上述实验均重复3次,具体操作方法参见《土壤农业化学分析方法》[25]。
1.4 数据处理
研究采用了数理分析方法,选用Excel 2010记录试验数据、Origin9.0制图,运用SPSS 20.0分析各态氮素与土壤CEC的差异显著性和相关性。
2 结果与分析
2.1 不同植物群落湿地土壤CEC的分布特征
土壤CEC指土壤溶液在酸碱度恒定时,单位质量干土所吸收的全部交换性阳离子的物质的量[11]。土壤交换性阳离子的吸附与解吸作用以及养分的固持与淋失作用是影响土壤肥力的主要因素[26]。图2所示为本研究中不同植物群落湿地土壤CEC的分布特征,4种植物群落土壤中CEC芦苇群落含量最高为(362.96±61.48) mg/kg(Mean±SD,下同),牛筋草群落含量最低为(270.33±41.63) mg/kg,其他两种群落含量分别为(303.43±32.87) mg/kg和(332.68±26.18) mg/kg。忽略4种植物群落生境的细微差别,牛筋草群落与芦苇群落土壤CEC差异性显著(p<0.05),而二者与李氏禾、香薷群落无差异。
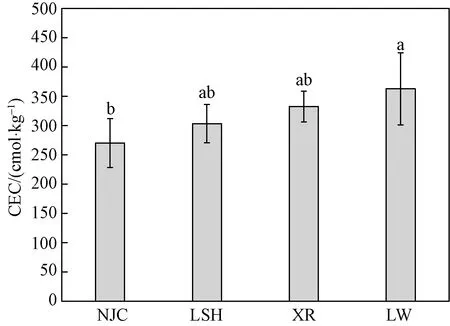
不同字母表示植物群落之间土壤CEC含量的差异性图2 4种湿地植物群落土壤中CEC的含量分布Fig.2 The distribution of cation exchange capacity in soil of four wetland plant communities
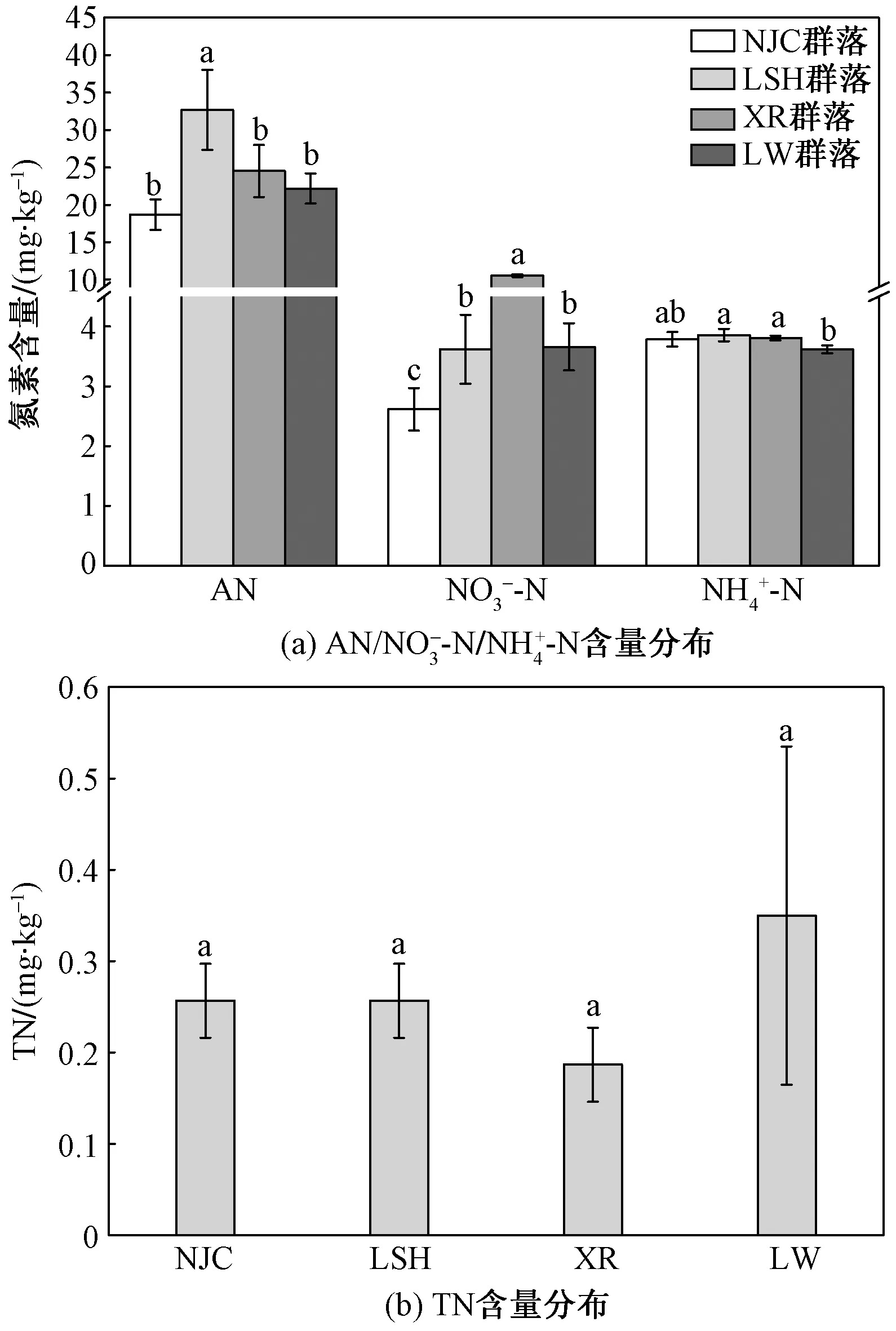
不同字母表示植物群落之间土壤各态氮含量的差异性图3 4种湿地植物群落土壤中及TN各态氮含量分布Fig.3 The distribution of nitrogen and TN) in the soil of four wetland plant communities
2.2 不同植物群落湿地土壤各态氮的分布特征
图3所示为不同植物群落湿地土壤各态氮的分布特征,李氏禾群落下土壤速效氮(available nitrogen,AN)含量差异显著(p<0.05),且李氏禾群落下土壤速效氮含量为4种植物群落最高,达到了(32.67±5.35 )mg/kg,而牛筋草群落土壤速效氮含量最低,为(18.67±2.02) mg/kg;铵态氮和硝态氮作为速效氮中两种重要的无机氮,二者含量在不同植物群落下各不相同,香薷群落土壤硝态氮含量较其他3种植物群落较高,达到了(10.52±0.18) mg/kg,4种植物群落土壤硝态氮含量差异极显著(p<0.01);在4种植物群落土壤中铵态氮含量分布范围为3.55~3.93 mg/kg,较硝态氮含量相比,各植物群落土壤铵态氮含量差异较小(p>0.05)。总氮(total nitrogen,TN)含量在不同植物群落土壤中在数值上差异较大,但总氮含量与不同植物群落差异性并不显著(p>0.05),其中芦苇植物群落土壤中总氮含量最高,达到了(350.00±185.20) mg/kg,香薷群落土壤总氮含量最低,为(186.67±41.41) mg/kg。
2.3 不同植物群落湿地土壤各态氮与土壤团聚体粒径的关系
4种湿地植物群落(NJC、LSH、XR、LW)土壤中硝态氮、铵态氮、速效氮及总氮在不同土壤团聚体粒径(d=0.250、0.177、0.147、0.125 mm)的含量分布,如图4所示。牛筋草群落与香薷群落,各态氮素在不同粒径土壤团聚体中含量差异均不明显(p>0.05),香薷群落中各粒径土壤团聚体中硝态氮含量整体分布与其他3种植物群落相比均较高,当土壤团聚体粒径为0.25 mm时,硝态氮含量最高达(8.43±1.24) mg/kg,当土壤团聚体粒径d≤0.147 mm时,硝态氮含量分布趋于稳定;李氏禾群落不同粒径土壤团聚体中硝态氮、速效氮和总氮含量不具有显著差异(p>0.05),硝态氮与速效氮含量分布基本趋势一致,速效氮与总氮与其他3种植物群落相比整体含量较高;铵态氮在不同粒径土壤团聚体中含量均较为稳定,均未超过4.0 mg/kg,其中在土壤团聚体粒径d≥0.177 mm时,铵态氮含量具有显著差异(p<0.05)。芦苇群落不同粒径土壤团聚体中速效氮与总氮含量分布不具有显著差异(p>0.05),硝态氮含量分布整体较其他3种植物群落相比含量最低,当土壤团聚体粒径为0.177 mm时,硝态氮含量与土壤团聚体粒径为0.125 mm时具有显著差异(p<0.05),与铵态氮分布差异显著性一致。
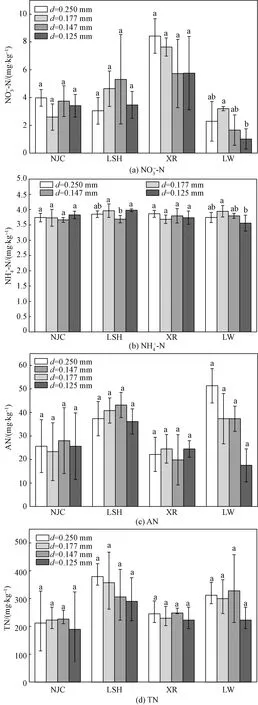
不同字母表示植物群落内不同粒径土壤团聚体各态氮含量的差异性图4 4种湿地植物群落土壤及TN在不同粒径土壤团聚体中含量分布Fig.4 The distribution of nitrogen AN and TN) in soil aggregates with different particle sizes of four wetland plant communities
3 讨论
3.1 不同植物群落对湿地土壤各态氮含量的影响
各群落下土壤总氮平均含量分布趋势为TNLW>TNNJC≈TNLSH>TNXR,这与张珍明等[27]研究草海沼泽草地总氮含量基本一致。不同植物群落下土壤总氮含量差异反映了植物群落的固氮能力和初级生产力差异,白红军等[3]研究认为,湿地植物种类及其自身对元素的选择性吸收、滞留作用会使土壤中不同元素含量形成较大差异。8月草海气温升高,伴随着雨水频次及降雨量增加,芦苇群落处于快速生长期,其光合作用和蒸腾作用均强于其他植物群落,物质循环加快,植物根系大量吸收无机态氮,因此芦苇群落土壤中铵态氮和硝态氮与其他群落差异显著;同时凌敏等[28]研究表明在黄河三角洲湿地植物群落地势对土壤氮的累积有决定性影响,芦苇根部沉水,抑制硝化作用且硝态氮易淋失;芦苇为高大草本植物,大量的植物凋落物及残体经微生物分解后,增加了表层土壤有机质含量[29],同时淹水造成湿地土壤有机碳矿化速率减缓,降低了微生物参与的氮循环速率,综合以上两方面原因,造成芦苇群落土壤中硝态氮和铵态氮含量与其他植物群落有显著差异,且总氮的含量较高[13]。李氏禾群落速效氮较高,由于李氏禾为矮小草本植物,生长在岸边浅水滩涂,合适的温度、水文及溶解氧条件使得土壤中促进微生物参与的氮循环,从而速效氮中其他氮含量较高。土壤腐殖质是土壤保持较高CEC的一个重要影响因子[26],这也解释了芦苇群落表层土壤CEC和总氮含量较高的原因[30]。
4种植物群落对土壤中铵态氮含量分布差异显著,牛筋草群落硝态氮含量低于铵态氮含量,可能是8月植物处于生长旺盛期[31],会大量吸收土壤中硝态氮;另一方面,牛筋草群落土壤中CEC最低,反映了土壤 中有机质含量低,亚硝化或硝化细菌没有充足的碳源用于生命活动,同时8月草海湿地雨水频次及降雨量增加,导致牛筋草群落表层土壤含水量增加,土壤中好氧微生物耗氧速率大于水体复氧速率,对微生物活动特别是硝化作用产生抑制作用,这与王大鹏等[32]研究水分对土壤硝化作用影响的结果一致;同时铵态氮易被土壤胶体吸附,而硝态氮易淋失[33],最终表现为牛筋草群落铵态氮含量略高于硝态氮含量,同时也是速效氮含量低于其他植物群落的主要原因。由于香薷群落自身植物特性,生长地势均高于其他植物群落,丰水期土壤含水率不会发生剧烈波动,王大鹏等[32]认为土壤水分含量为60%持水量时,硝化速率及硝化率最高,良好的水文、温度条件和充足有机质含量利于土壤硝化细菌等微生物活动,促进土壤中硝化作用使铵态氮向硝态氮转化;有研究表明,当湿地土壤可利用性铵态氮无法满足微生物固持需求时,微生物对硝态氮有较高固持量[34];此外植物生长还需吸收无机氮,综合几种因素使得香薷群落硝态氮含量高于铵态氮含量。
3.2 不同植物群落对湿地土壤团聚体氮素含量的影响
土壤团聚体作为土壤结构最基本的单元,通过多重物理化学机制对团聚体中有机碳氮进行有效固持和保护[35]。学者一般以0.25 mm为界对土壤团聚体进行分组,将团聚体分为大团聚体和微团聚体两类[36-37],有研究表明喀斯特地区土壤团聚体主要以大团聚体(>0.5 mm)为主,总量超过80%,在喀斯特地区以封育为土地利用方式下,团聚体粒径为0.054~0.25 mm时,土壤团聚体有机质含量处于较低水平[21],而有机质作为团聚体形成的重要胶结物,微团聚体提供良好的吸附表面[38],起到吸附土壤中各类营养物质的作用。
在不同植物群落下各粒径土壤团聚体中铵态氮含量基本呈现无差别分布的特征,这可能由于土壤团聚体粒径位于0.25~0.125 mm时,土壤中有机质的含量较低,无法对铵态氮起到吸附作用或吸附作用力较小。土壤有机碳、硝态氮含量和微生物生物量氮是影响土壤硝化与反硝化潜势的关键共性因子[9],在不同粒径土壤团聚体中,有机质含量随着土壤团聚体粒径变小而降低[21],抑制土壤中铵态氮向硝态氮转化过程。当土壤团聚体粒径为0.125 mm时,不同植物群落下硝态氮含量均表现为比其他粒径团聚体低,这可能受到土壤团聚体中有机质含量的影响。当香薷群落土壤团聚体粒径为0.147 mm时,土壤硝化作用已明显受到影响。
而两种草海湿地典型挺水植物群落——李氏禾与芦苇群落,在不同粒径团聚体中硝态氮含量与香薷群落相比,分布趋势并不一致。8月草海湿地迎来夏季丰水期,李氏禾与芦苇群落淹水较深,抑制土壤中硝化作用,同时植物生长旺盛期对硝态氮吸收作用较为强烈,淹水还对硝态氮有淋失作用,同时还包括植物光合作用及蒸腾作用加快物质循环;硝化作用还受有机碳的数量和质量影响[39],特别是芦苇群落有机质含量高,抑制芦苇群落土壤硝化作用,以上几种因素共同发挥作用,李氏禾群落最终表现为在粒径为0.147 mm土壤团聚体、芦苇群落在0.177 mm土壤团聚体中硝态氮含量较高,但整体水平较低;芦苇群落硝态氮含量整体水平最低。
各植物群落在不同粒径土壤团聚体中总氮和速效氮含量差异性不显著,在芦苇和李氏禾群落不同粒径土壤团聚体中总氮和速效氮含量基本均高于其他植物群落,这也进一步说明了淹水削减土壤有机碳矿化速率从而影响了土壤氮循环;芦苇群落土壤CEC高,总氮含量分布高,牛筋草群落正相反,与陈忠柳等[40]研究结果一致,总氮含量与土壤有机质和CEC含量呈正相关,土壤阳离子交换作用变化(45%)主要受土壤基本理化因素影响。此外草海湿地周围特别是北岸人为活动较为强烈[29],李氏禾群落和芦苇群落为挺水植物,土壤易受到水体中氮素的影响[39],同时有机态氮占总氮的95%以上[4],因此不同植物群落下总氮整体含量差异较为明显。
4 结论
(1)8月植物处于生长旺盛期,同时草海处于丰水期,植物群落表层各氮素含量分布受植物种类及自身凋落物、降雨量和地势、微生物活动、有机质和生长期等因素的影响,土壤硝化作用受到抑制或促进,最终表现为各植物群落铵态氮和硝态氮含量分布有显著差异。
(2)湿地表层土壤氮素含量受水体中外部氮输入及氮素本身在自然土壤中分布结构的影响,因此各植物群落氮素含量表现并不一致,特别是土壤中总氮含量。同时本研究可为后续进一步为草海地区氮输入及其对该地区碳汇和碳储量研究提供基本数据支撑。
