不同耐热基因型芝麻苗期对高温胁迫的生理响应机制
2022-01-10苏小雨高桐梅魏利斌王东勇朱松涛卫双玲
苏小雨,高桐梅,李 丰,魏利斌,田 媛,王东勇,朱松涛 ,卫双玲
(1.河南省农业科学院,芝麻研究中心,河南 郑州 450002;2.河南省特色油料作物基因组学重点实验室,河南 郑州 450002;3.农业部黄淮海油料作物重点实验室,河南 郑州 450002;4.商水县农业农村局,河南 商水 466100)
高温是全球普遍存在的自然灾害,尤其进入21 世纪以来,全球气候变暖引起的高温天气频发以及极端高温,严重限制植物生长发育,是影响作物产量的主要非生物胁迫因素之一[1]。因此,研究作物对高温的响应机制对认知和提升作物抗高温能力具有十分重要的理论和现实意义。

芝麻(SesamumindicumL.)是我国特色油料作物之一,目前,高温灾害是制约我国芝麻生产的重要非生物胁迫因子,关于芝麻对高温胁迫响应研究报道较少,因此,探究芝麻对高温的响应机制十分必要。本研究选用耐热性和热敏感的2个芝麻品种为材料,在苗期进行不同时间的高温胁迫,测定叶片中膜脂过氧化相关指标(相对电导率、丙二醛、ROS)、抗氧化酶(SOD、POD和CAT)活性、光合气体交换参数、叶绿素荧光参数,并进行气孔扫描电镜观察、叶绿体透射电镜观察,分析耐热性差异显著的2个品种响应高温胁迫时生理变化差异,为深入探究芝麻耐热机理和品种改良提供理论依据。
1 材料和方法
1.1 供试材料
选用的郑太芝3号(耐热性品种)和SP19(热敏性品种)芝麻品种由河南省农业科学院芝麻研究中心提供。选取籽粒饱满、大小均一的种子播种在塑料盆(直径16 cm,高度14 cm)中,每个处理3 盆。培养基质为营养土和珍珠岩配制比例3∶1,培养温度为30 ℃,相对湿度为70%。待植株长至2对真叶时,选取长势一致的植株进行处理。
1.2 试验方法
将郑太芝3号和SP19植株分别置于30 ℃(对照,CK)和45 ℃(高温,HT)恒温培养箱中连续处理10 d,培养条件:昼夜14 h/10 h,相对湿度70%,光照350 μmol/(m2·s)。待处理4,6,8,10 d后进行形态指标、相对含水量和光合特性指标测定,收集叶片,在液氮中速冻后保存于-80 ℃超低温冰箱中,用于生理生化指标测定。
1.3 测定项目与方法
1.4 数据处理与统计分析
试验数据利用Microsoft Excel 2010软件进行统计和整理,利用SPSS 16.0进行数据差异显著性分析。
2 结果与分析
2.1 高温胁迫对芝麻幼苗生长发育和相对含水量的影响
由表1可知,随高温胁迫时间的延长,2种芝麻的株高、叶长、叶宽和相对含水量相比对照均呈下降趋势。高温处理4 d,郑太芝3号和SP19幼苗的株高和叶宽下降不显著(P>0.05),仅SP19幼苗的叶长开始显著下降(P<0.05),较对照减少了15.82%。胁迫6 d,SP19幼苗的株高、叶长和相对含水量均显著下降(P<0.05),分别较对照减少了9.28%,17.32%和8.13百分点。当胁迫8,10 d时,郑太芝3号幼苗的叶长较对照下降了14.42%和14.61%,相对含水量较对照下降了11.07,14.87百分点,差异均达显著水平,其他指标下降不显著;而SP19幼苗的株高下降了8.21%和14.03%,叶长下降了25.79%和31.30%,叶宽下降了10.48%和14.17%,相对含水量下降了18.68,22.07百分点,差异均达显著水平。品种间比较来看,在高温胁迫下,郑太芝3号的株高、叶长、叶宽和相对含水量受抑制程度均低于SP19。
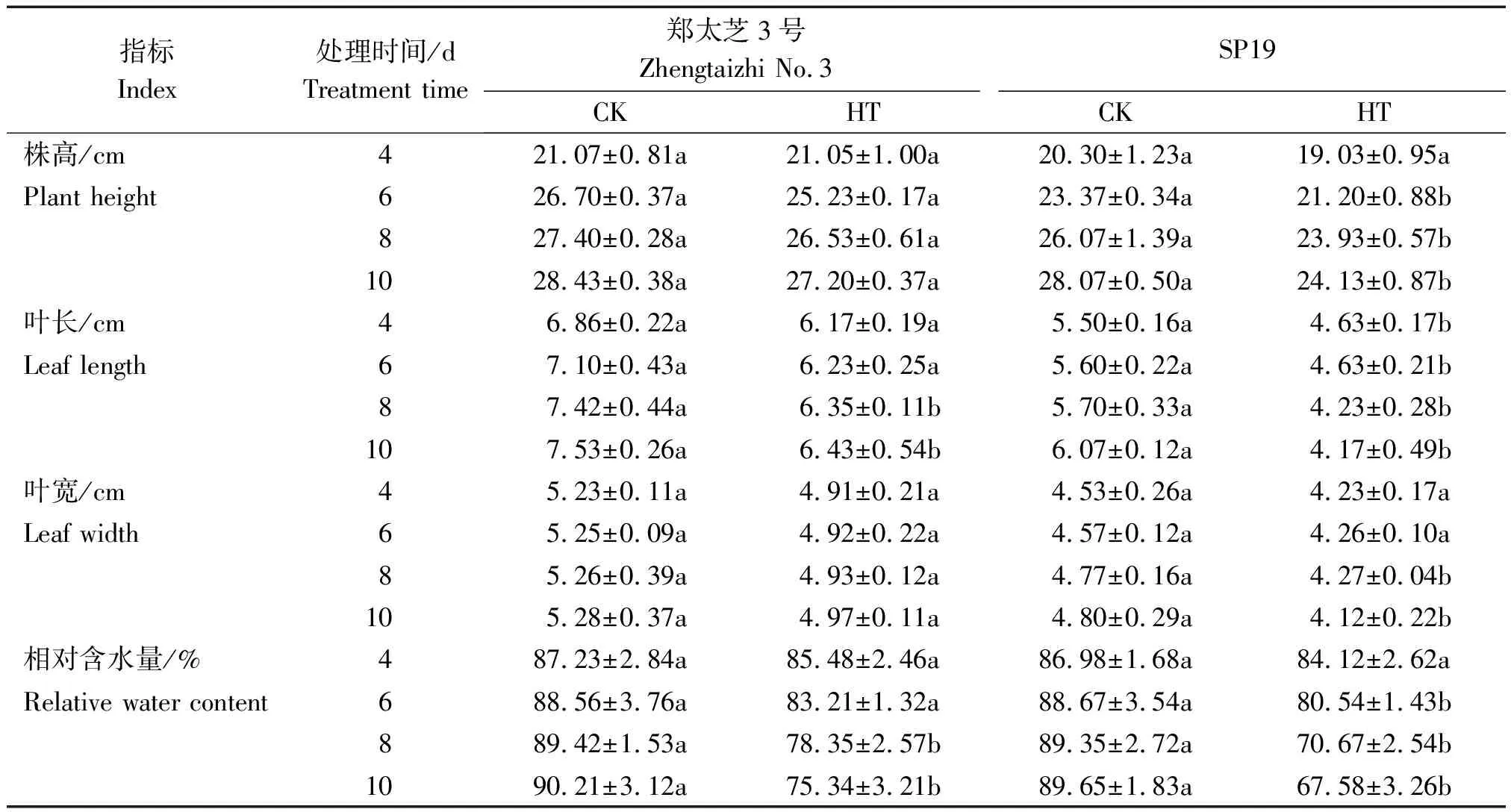
表1 高温胁迫对芝麻幼苗生长特性和相对含水量的影响Tab.1 Effects of heat treatment on growth characteristics and relative water content of sesame seedlings
2.2 高温胁迫对芝麻幼苗相对电导率、丙二醛含量和活性氧水平的影响
由图1可知,高温胁迫不同程度地提高了芝麻幼苗的相对电导率、丙二醛、活性氧含量。郑太芝3号和SP19幼苗的多数相关指标均在高温胁迫6 d开始显著(P<0.05)增加。高温胁迫6 d,郑太芝3号幼苗叶片的相对电导率较对照显著增加了3.03百分点,8,10 d极显著增加了4.50,5.83百分点,丙二醛含量分别增加了22.31%,42.68%和82.47%,过氧化氢含量分别增加了20.11%,32.44%和60.87%,超氧阴离子产生速率分别增加了38.80%,62.42%和77.25%,而SP19相对电导率分别增加了4.33,8.40,13.43百分点,丙二醛含量分别增加了44.85%,70.62%和97.21%,过氧化氢含量分别增加了20.11%,32.44%和60.87%,超氧阴离子产生速率分别增加了48.31%,94.62%和134.35%。品种间比较来看,在高温胁迫下,SP19幼苗的质膜相对透性和氧化损伤程度明显高于郑太芝3号。

不同小写字母表示同一时期不同处理存在显著差异(P<0.05);不同大写字母表示存在极显著差异(P<0.01)。图2-5同。Different lowercase letters indicate that there are significant differences (P<0.05) between different treatments in the same period;Different big letters indicate a very significant difference(P<0.01). The same as Fig.2-5.
2.3 高温胁迫对芝麻幼苗抗氧化酶活性的影响
由图2可知,随高温胁迫时间的延长,郑太芝3号和SP19幼苗的SOD和CAT活性均呈先增加后降低的变化趋势,而POD活性均逐渐升高。在胁迫8 d时,郑太芝3号和SP19幼苗的SOD活性均增至最高值,较对照分别增加了2.93,2.63倍,差异均达极显著水平。郑太芝3号和SP19幼苗的CAT活性分别在高温胁迫8,6 d增至最高值,分别增加了1.10,0.99倍,差异均达极显著水平。在胁迫10 d时,郑太芝3号和SP19幼苗的POD活性较对照分别增加了1.86,1.52倍,差异达极显著水平。品种间比较来看,郑太芝3号SOD、POD和CAT活性均高于SP19。
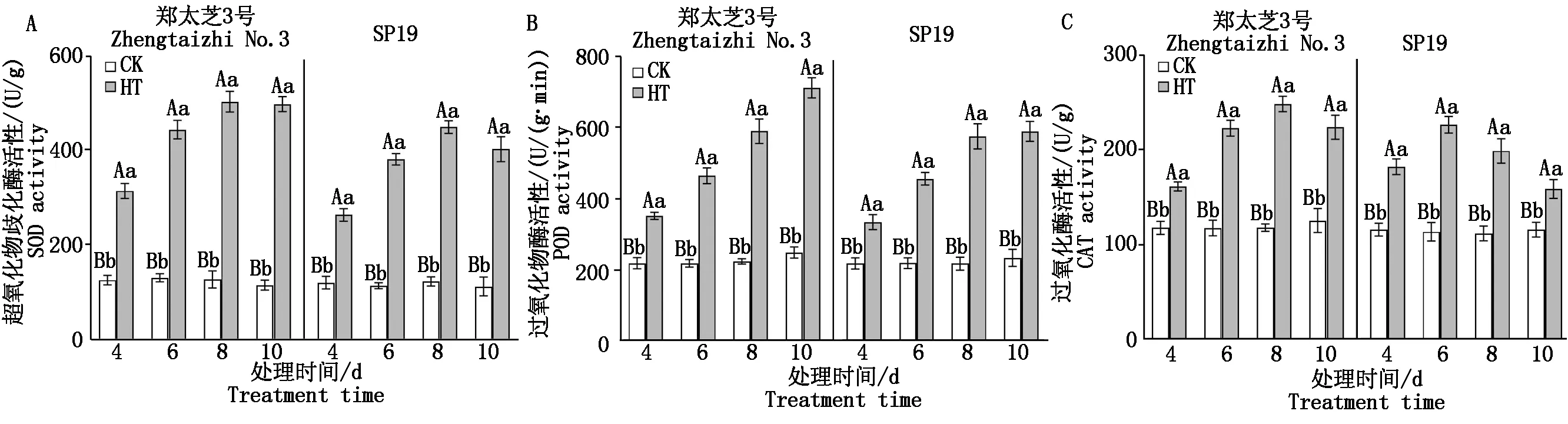
图2 高温胁迫对芝麻幼苗抗氧化酶活性的影响Fig.2 Effects of heat treatment on antioxidant enzyme activities in the seedling leaves of sesame
2.4 高温胁迫对芝麻幼苗叶绿素含量的影响
由图3可知,随高温胁迫时间的延长,郑太芝3号和SP19幼苗的叶绿素含量(SPAD值)呈不断下降趋势,从高温胁迫 6 d郑太芝3号和SP19幼苗的叶绿素含量均开始显著下降。胁迫至 10 d,郑太芝3号和SP19幼苗的叶绿素含量分别极显著降低了14.37%和27.55%。
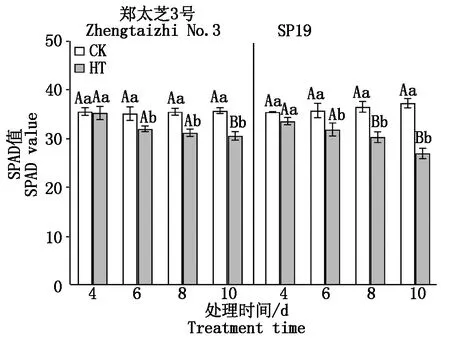
图3 高温胁迫对芝麻幼苗叶绿素含量的影响Fig.3 Effects of heat treatment on chlorophyll content in the seedling leaves of sesame
2.5 高温胁迫对芝麻幼苗气体交换参数的影响
由图4可知,随着胁迫时间的延长,郑太芝3号幼苗的Pn、Tr、Ci和Gs 均呈现逐渐下降趋势。胁迫4 d 时,郑太芝3号幼苗的Pn较对照略有升高,但差异不显著,Tr、Ci和 Gs较对照增加了30.97%,10.40%和8.9%,差异均达显著水平,胁迫6 d,郑太芝3号幼苗的Pn显著下降了11.26%,而Tr、Ci和Gs下降不显著,胁迫 8 d,郑太芝3号幼苗的Pn较对照显著下降了11.58%,10 d极显著下降了18.41%,而Tr较对照分别下降了21.31%和28.49%,Ci分别下降了18.78%和30.45%,Gs分别下降了16.10%和23.31%,差异均达极显著水平。

图4 高温胁迫对芝麻幼苗气体交换参数的影响Fig.4 Effects of heat treatment on gas exchange parameter in the seedling leaves of sesame
SP19幼苗的Pn、Tr、Gs随着胁迫时间的增加也均呈现逐渐下降趋势,而 Ci 在处理 6 d时降至最低值后呈现上升趋势。胁迫 4 d 时,SP19幼苗的Ci和Gs较对照略有升高,但差异不显著,Pn略有下降,Tr较对照增加了30.36%,差异达极显著水平,胁迫6 d 时,SP19幼苗的Pn下降了21.87%,Tr下降了11.85%,Ci下降了10.74%,Gs下降了9.37%,差异均达显著水平,在胁迫8 ,10 d 时,SP19幼苗的Pn较对照分别下降了37.90%和56.98%,Tr较对照分别下降了28.94%和40.60%,Ci较对照分别提高了5.62%和17.49%,Gs分别下降了26.75%和38.02%,除8 d Ci外差异均达显著水平。
2.6 高温胁迫对芝麻幼苗叶绿素荧光参数的影响
如图5所示,2种供试芝麻品种幼苗叶片的Fv/Fm、φPSⅡ和ETR(Ⅱ)均随着高温胁迫时间的延长而呈现不断下降趋势,郑太芝3号幼苗的Fv/Fm、φPSⅡ和ETR(Ⅱ)均在胁迫8 d开始显著下降,而SP19幼苗的相关指标在胁迫6 d开始呈现显著下降。胁迫6 d SP19幼苗的Fv/Fm、φPSⅡ和ETR(Ⅱ)较对照分别下降了12.30%,11.04%和9.52%。高温胁迫8 d,10 d郑太芝3号幼苗的Fv/Fm较对照分别下降了14.29%和20.92%,φPSⅡ较对照分别下降了17.61%和31.62%,ETR(Ⅱ)分别下降了11.10%和22.72%;而SP19幼苗的Fv/Fm较对照分别下降了24.02%和37.15%,φPSⅡ较对照分别下降了32.68%和54.90%,ETR(Ⅱ)分别下降了22.55%和34.94%。

图5 高温胁迫对芝麻幼苗叶绿素荧光参数的影响Fig.5 Effects of heat treatment on chlorophyll fluorescence parameters in the seedling leaves of sesame
2.7 高温胁迫对芝麻幼苗叶片气孔形态的影响
通过利用扫描电镜对2种芝麻叶片的气孔形态进行观察,如图6所示,在正常环境下,2种芝麻品种的气孔开放程度较高,气孔周围下表皮细胞表面较为平滑,且蜡质纹饰不明显,而在45 ℃高温处理10 d后,郑太芝3号和SP19的叶片气孔发生明显改变,气孔周围的表皮细胞呈现凹凸不平,大部分气孔处于关闭或半关闭状态并且保卫细胞皱缩变形,陷于周围的表皮内。品种间比较来看,高温胁迫下,郑太芝3号叶片气孔不完全闭合,气孔周围的表皮细胞趋于平整光滑,保卫细胞皱缩变形不明显,而SP19叶片气孔处于完全闭合状态,保卫细胞皱缩变形严重,且周围的表皮细胞凹凸不平严重。

图6 高温胁迫10 d对芝麻幼苗气孔形态的影响Fig.6 Effects of heat treatment for 10 d on stomatal morphology in the seedling leaves of sesame
2.8 高温胁迫对芝麻幼苗叶片叶绿体结构的影响
如图7所示,常温下,芝麻叶绿体呈月牙状,紧贴细胞膜,膜结构完整,基粒片层规则排列且清晰,均匀分布在叶绿体中,叶绿体中存在着淀粉粒和少量嗜锇颗粒。高温处理10 d后,芝麻叶绿体体积变大,形状趋于圆型,叶绿体膜有解体现象,基粒片层堆叠增厚,淀粉粒消失,叶绿体移动至细胞内部。同时,在SP19叶绿体上观察到大量嗜锇颗粒,基粒片层部分已经断裂甚至解体,尽管郑太芝3号叶绿体上观察到嗜锇颗粒有所增加但明显少于SP19,基粒片层断裂较少。这些结果表明,高温下郑太芝3号叶绿体结构的稳定性高于SP19。
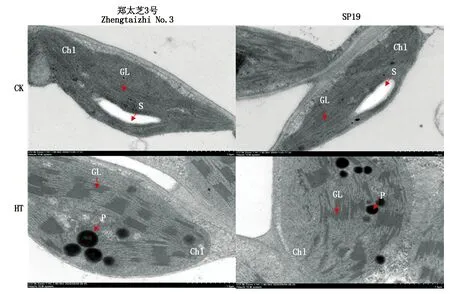
Chl.叶绿体;S.淀粉粒;P.嗜锇颗粒;GL.基粒片层。Chl.Chloroplast;S.Starch grain;P.Plastoglobule;GL.Grana lamellae.
3 讨论与结论
植物对高温胁迫响应是较为复杂又相互协调的生物学调控过程,其中主要包括分子、细胞、生理生化的响应,导致植物形态发生改变[17]。研究表明,高温胁迫能够引起植物光合作用和氧化应激发生明显改变,造成植物生长发育受到抑制[18]。目前,关于植物对高温胁迫的生理生化响应在较多作物中已作了大量研究,但关于芝麻对高温的生理响应机制尚无报道,本研究对芝麻幼苗在不同胁迫时间下的形态、生理以及显微结构变化进行全面的分析,并比较不同耐热性芝麻品种响应高温胁迫的生理特性差异,旨在明确芝麻幼苗对高温胁迫响应和适应机制。
3.1 不同耐热性芝麻品种生长参数对高温胁迫的响应变化
高温胁迫下,植物生理代谢过程出现紊乱,造成其生长发育受到抑制[19]。本研究表明,随高温胁迫时间延长,不同耐热性芝麻幼苗叶长、叶宽以及相对含水量不同程度下降,热敏感芝麻品种的株高受到显著抑制,而耐热性芝麻品种株高略受抑制,这一结果在其他作物研究中也得到证实[20-21],说明高温胁迫对芝麻的生长发育产生了一系列负面影响。此外,不同耐热性芝麻品种响应高温胁迫时存在着差异,本研究中,胁迫4~10 d时,耐热性品种郑太芝3号的株高和叶宽略有下降但不显著,叶长和相对含水量显著下降的胁迫时间为8 d;而热敏感品种SP19株高和相对含水量均在胁迫时间6 d时开始显著下降,叶宽开始显著下降时间为8 d,叶长为4 d,说明高温胁迫下芝麻幼苗的生长发育抑制程度与不同品种耐热性密切相关,且相比热敏感品种SP19,高温胁迫对耐热性品种郑太芝3号生长的影响程度较小。
3.2 不同耐热性芝麻品种光合特性对高温胁迫的响应变化
光合作用在植物的生长发育中扮演着重要角色,较多研究表明,光合作用对高温胁迫较为敏感,高温胁迫导致植物的光抑制,使光系统破坏,促进叶绿素的降解和光合同化的降低[22-23]。在本研究中,随着高温胁迫的加剧,光合指标 Pn、Gs、Tr、Ci和相对叶绿素含量均降低,表明高温胁迫对芝麻幼苗的光合作用具有显著的抑制作用。此外,研究发现,在胁迫4 d时,2种芝麻品种幼苗的叶绿素含量略微减少,而Pn、Gs、Tr、Ci较对照均有不同程度的提高,这可能是由于在较短时间的高温胁迫下,芝麻通过提高自身的蒸腾作用,启动降温的自身保护机制,诱导气孔的开放,增加了外界二氧化碳的摄入量,从而提高了自身的净光合速率。植物光合作用的抑制主要受气孔和非气孔限制,随着胁迫时间的延长,植物为了减少自身水分大量蒸发,保持体内细胞膨胀压力,诱导气孔关闭,但减少了与外界环境的气体交换,同时叶绿素合成受到抑制,从而造成Pn的下降[24-25],本研究结果也证实这一结论,在高温胁迫6 d时,2种芝麻品种Pn、Gs、Tr和Ci 都呈降低趋势,而郑太芝3号的Fv/Fm、φPS Ⅱ 和ETR 在处理 6 d 时变化较小,仅SP19的Fv/Fm、φPS Ⅱ 和ETR有所下降,表明短期的高温胁迫下芝麻幼苗光合能力受抑制的主要因素为气孔限制。然而在高温胁迫8~10 d,2种芝麻品种的光合指标 Pn、Gs和Tr均不同程度地下降,有趣的是,敏感品种SP19的Ci与郑太芝3号结果相反呈上升趋势。前人研究表明,Ci 和Gs变化趋势是判断Pn下降的气孔或非气孔因素的主要依据,Ci 值随胁迫程度上升,而Gs 却表现为下降趋势时,非气孔限制是Pn下降的主要因素[26- 27]。进一步对高温胁迫10 d 2种芝麻品种幼苗叶片的气孔开放程度进行观察,发现耐热性品种郑太芝3号气孔处于半关闭状态而热敏感品种SP19气孔为完全关闭状态。这些结果表明,在重度高温胁迫时,非气孔限制是芝麻叶片Pn 下降的主要因素。相比较于热敏感品种SP19,耐热性品种郑太芝3号表现为气孔和非气孔限制较低,说明芝麻品种的耐热性与气孔限制密切相关。
非气孔限制主要由于植物叶片叶绿素含量减少,光系统的稳定性和完整性遭到破坏,叶绿体机构遭到破坏,致使光系统活性降低,从而造成Pn 下降[25]。研究表明,在非生物胁迫下,PSⅡ是光系统最敏感的部位,易受到胁迫损伤,而光系统Ⅰ较为稳定[28]。PSⅡ作为光系统反应中心,是调节光能吸收、转化以及电子传递速率的主要部位[29]。本研究发现,2 个芝麻品种随着高温胁迫时间延长,Fv/Fm、φPSⅡ和ETR(Ⅱ)均呈下降趋势,表明高温胁迫下,芝麻幼苗叶片PSⅡ受到损害,活性降低。相比较于热敏感品种SP19,高温胁迫下耐热性品种郑太芝3号Fv/Fm、φPSⅡ和ETR(Ⅱ)下降程度较低,这与前人研究结论相一致[10],表明PSⅡ活性在芝麻耐热性中扮演着重要角色。对高温胁迫10 d 2种品种叶片叶绿体结构观察表明,高温胁迫后2种品种叶绿体结构受到不同程度的破坏,整体表现为叶绿体趋于圆形,向膜内移动,基粒堆积加厚和片层断裂,嗜锇颗粒变大和增多,与前人结果较为相似[30-31],表明高温导致芝麻叶绿体超微结构发生变化从而造成光合机构的破坏。相比较于热敏感品种SP19,耐热性品种郑太芝3号的叶绿体结构较为稳定,表明叶绿体结构的稳定性对提高芝麻的耐热能力起着关键性作用。此外,这些结果进一步证实了重度高温胁迫下,非气孔限制是导致芝麻幼苗Pn 下降的主要原因。
3.3 不同耐热性芝麻品种活性氧和抗氧化酶活性对高温胁迫的响应变化
较多研究表明,高温胁迫打破 ROS产生和清除之间的平衡,诱导其过量积累[32]。高温胁迫导致叶绿体结构的破坏,抑制了 PS Ⅱ 活性,叶绿体作为ROS的主要产生部位,PS Ⅱ 活性降低导致激发能量从天线传递到反应中心受阻,引起能量过剩从而产生过量的ROS,造成氧化损伤加剧膜脂过氧化作用,MDA含量增加,破坏植物细胞内部环境[33]。本研究表明,随着高温胁迫时间的延长,ROS和MDA含量均呈现不同程度的上升趋势,说明高温胁迫下,芝麻幼苗生长受到抑制与ROS的过量积累密切相关,加剧了光合机构和功能的破坏,造成植物受到氧化损伤。相比较于热敏感品种SP19,同一胁迫时间下耐热性品种郑太芝3号ROS和MDA含量增加程度较低,表明维持ROS的平衡和降低膜脂过氧化程度是芝麻具有较高耐热性的关键因素。
综上,本试验通过对2种不同耐热性芝麻品种的生长特性和生理变化分析,发现高温胁迫能够破坏光合系统结构和功能,造成ROS过量积累,显著降低叶绿素含量和净光合速率,明显抑制芝麻幼苗的生长,尤其热敏感品种SP19 中光系统活性中心对高温胁迫较为敏感,不可逆损伤明显高于郑太芝3号,且净光合速率降低在轻度高温胁迫下主要由于气孔限制因素,随胁迫时间延长非气孔限制因素占主导地位。另外,明确了郑太芝3号在高温胁迫下,植株体内抗氧化酶活性较高、膜脂过氧化程度较低、叶绿体结构稳定和叶绿素降解少是其具有高耐热性的关键。这些结果为芝麻耐热性品种选育和改良提供了理论依据。
