东南太平洋智利竹筴鱼的洄游路线
2022-01-10丁鹏邹晓荣冯超白思琦
丁鹏,邹晓荣,2,3,4,5*,冯超,白思琦
(1.上海海洋大学 海洋科学学院,上海 201306; 2.远洋渔业创新中心,上海 201306; 3.大洋渔业资源可持续开发教育部重点实验室,上海 201306; 4.国家远洋渔业工程技术研究中心,上海 201306; 5.农业农村部大洋渔业资源环境科学观测试验站,上海 201306)
鱼类洄游路线研究和种群划分是渔业资源学和生态学的重点。对鱼类而言,种群既是数量生态学的研究单位,也是渔业资源量的研究单位[1]。经济鱼类往往具有集群和洄游特征,不同种群的洄游,往往具有各自相对固定的时间和路线[2],因而形成种群特有的时空分布格局。鱼类种群划分和洄游路线的研究,对于渔场鱼汛的确定和增殖措施的制定等均具有重要的科学意义[3]。
智利竹筴鱼Trachurusmurphyi被认为是世界上主要的海洋经济鱼种之一,广泛分布于南太平洋水域,其渔获量曾多年位居世界前列[4]。中国大型拖网船队于2000年开始开发利用东南太平洋公海智利竹筴鱼,该海域也迅速成为中国大型拖网渔业最主要的作业渔场。目前,国内外学者对智利竹筴鱼渔场学[5-7]、生物学特性[8-9]、渔业资源状况[10-11]、渔情预报[12]、种群结构[13]等进行了一系列研究,但对该鱼的洄游路线研究较少[14-15]。本研究中,拟利用2000—2003、2007—2009、2016年中国智利竹筴鱼捕捞日志及其生物学数据,采用神经网络模型和频率分布法对智利竹筴鱼的洄游路线进行研究,以期为中国智利竹筴鱼渔船生产调度及布局提供科学依据。
1 材料与方法
1.1 数据来源
智利竹筴鱼渔捞日志及其生物学数据均由中国远洋渔业协会大型加工拖网项目技术组提供,数据来源于大型拖网渔船“开欣”号和“开富”号。“开欣”号船长96.7 m、型深10.2 m、船宽16 m、总吨位4 407 t、主机功率2 567 kW。“开富”号船长91.1 m、型深9.2 m、船宽20 m、总吨位7 671 t、主机功率5 920 kW。时间范围为2000年5—6月、2001年7—10月、2002年8—9月、2003年1—2月、2007年5—8月、2008年5—7月、2009年8—10月、2016年3—10月,共计28个月,共20 096尾叉长数据。2007年5—8月、2008年5—7月共采集耳石样本782对。空间范围为30°~50°S、75°~110°W。
环境数据包括海表温度(sea surface temperature,SST)、海表面盐度(sea surface salinity,SSS)和叶绿素a浓度(chlorophyll a,Chl-a),均来源于ocean-watch网站(http://oceanwatch pifsc.noaa.gov)。
1.2 方法
1.2.1 年龄鉴定 为判断智利竹筴鱼各龄鱼的叉长范围及确定叉长频率分步法的组距,笔者对智利竹筴鱼的年龄进行鉴定,具体步骤如下。
1)将耳石放入体积分数为75%的乙醇中浸泡7 d,然后对耳石进行清洗并去除表面残留的有机物[16]。
2)将耳石放在干燥箱中60 ℃下烘24 h,然后按照精度0.1 mg进行称重[17]。
3)用P1200规格的防水砂纸在研磨台上将耳石两面打磨至0.5 mm厚,然后用70~90 g/L的抛光粉悬浮液将耳石两面抛光,之后在载玻片上用乙醇灯将耳石灼烧至褐色[18]。
4)使用NikonZOOM 645S体式显微镜对耳石放大40倍进行年轮判读和计数。
1.2.2 生长方程及参数估算 Von Bertalanffy生长方程拟合[19-20]为
Lt=L∞[1-e-k(t-t0)],
(1)
生长速度方程为
(2)
其中:Lt为t龄时的叉长(mm);L∞为可能达到的最大叉长(mm);k为相对生长指标;t为年龄(a);t0为理论生长起点年龄(a);dL/dt为叉长的生长速度。
1.2.3 洄游路线研究 每天记录渔船的作业位置,随机从渔获物中取样进行叉长测定,测定标准参照相关国家海洋调查规范[21]。本研究中,时间上以月份为单位,空间上将智利竹筴鱼渔场以经度1°为单位划分成若干个矩形海域定义为经向海区,以纬度1°为单位划分成若干个矩形海域定义为纬向海区,并采用频率分布法逐个海区统计渔获物叉长。
频率分布中组距越小,分的组数越多,所统计的渔获物叉长精度就越高,但数据统计过多则会导致数据分布较为分散。为确保所统计的渔获物叉长既具有高精度又不至于过度分散,本研究中将智利竹筴鱼生长速度最慢时的叉长与生长月份的比值向下取整数定为其组距。据统计,大部分渔区鱼群优势叉长频率在40%左右,故本研究中将优势叉长频率统一以40%为标准进行统计。优势叉长组内叉长平均值分别按照纬向海区纬度中点、经向海区经度中点顺序相连,分析智利竹筴鱼的时空分布。优势叉长组内叉长平均值和组距计算公式分别为
Z=C/(t×12),
(3)
X=∑Cij/I。
(4)
其中:Z为组距(mm);C为叉长生长速度最慢时的叉长(mm);t为年龄(a);X为优势叉长组内叉长平均值(mm);I为优势叉长组内样本数;Cij为i月j渔区优势叉长组中叉长(mm)。
以月份为单位对渔获物叉长进行叉长频率统计,优势叉长组内计算叉长对应的经纬度平均值定义为该月份的洄游站点,优势叉长组内计算叉长平均值按照由小到大顺序连接定义为鱼的洄游方向。智利竹筴鱼的产卵场依据本研究及相关文献进行判断[22]。有研究表明,智利竹筴鱼属于一个种群且经济鱼类具有相对固定的洄游时间和路线[2,13],因此,本研究中假设智利竹筴鱼在固定海区出现固定大小的鱼群。频率分布用R语言进行编程。洄游站点计算公式为
(5)
(6)
其中:Lonj为j月洄游站点经度位置;Latj为j月洄游站点经度位置;Mij为i年j月优势叉长组对应经度;Nkj为k年j月优势叉长组对应经度;mij为i年j月优势叉长组中对应纬度;nkj为k年j月优势叉长组中对应纬度;I为优势叉长组中叉长个数;n为月份个数。
1.2.4 智利竹筴鱼洄游路线的预测及验证 使用统计分析软件DPS进行BP神经网络模型的计算,以2000—2003、2007—2009年各月的叉长数据作为拟合样本,2016年各月的样本作为预测样本,以海洋环境因子SST、SSS、Chl-a结合拟合样本经纬度及叉长数据作为输入因子,隐含层为1层[23],隐含层神经元选取9[24]。以2016年预测叉长数据作为输出因子进行训练。每个月预测得到的叉长统计其优势叉长组范围,并计算在该范围内实际叉长量占当月总叉长量的比值,验证预报的准确性[24]。
神经网络模型结构如图1所示。
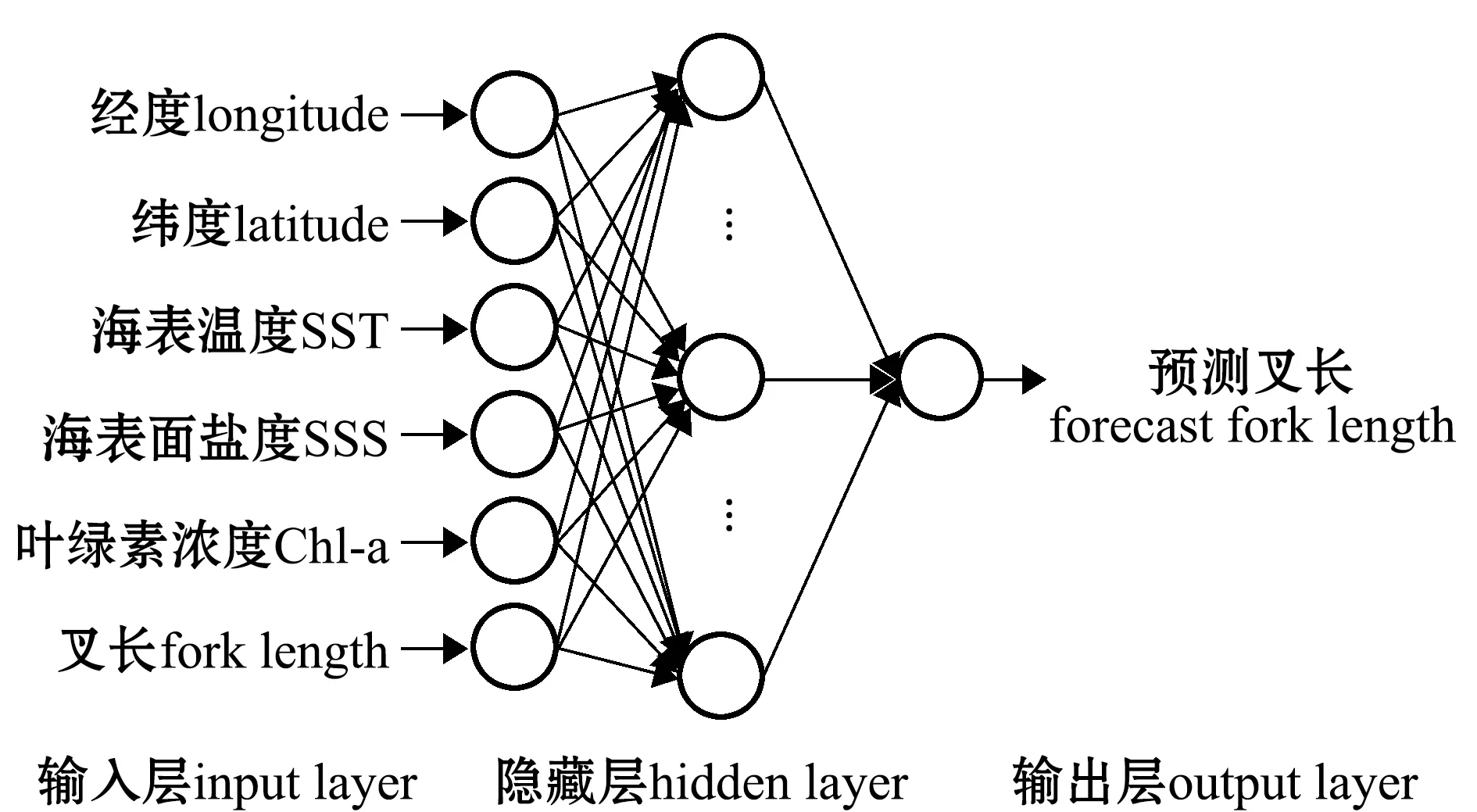
图1 BP模型结构Fig.1 Structure of BP model
2 结果与分析
2.1 智利竹筴鱼群体年龄及叉长生长
对采集的耳石进行年龄鉴定,鉴定结果如表1所示,其中,2 龄鱼叉长为 193~268 mm,3龄鱼叉长为269~317 mm,4 龄鱼叉长为318~349 mm,5 龄鱼叉长为350~388 mm,6 龄鱼叉长为389~437 mm,7 龄鱼叉长为438~463 mm,8龄鱼叉长为464~483 mm,9龄鱼叉长大于 483 mm。

表1 智利竹筴鱼不同年龄组的叉长Tab.1 Fork length of Chilean jack mackerel at different age classes
拟合得到智利竹筴鱼年龄与叉长的Von Bertalanffy生长方程、生长速度方程分别为
Lt=759.2947[1-e-1(t+1.856)],
从图2可见,智利竹筴鱼的叉长生长速度随年龄的增大呈现递减趋势,在9龄时为最小生长速度。取9龄时的叉长平均值与生长月份的比值即502.4/(12×9)=4.65作为组距。向下取整数,故取4为组距。

图2 叉长及叉长生长速度的变化趋势Fig.2 Change curve of fork length and growth rate of fork length
2.2 智利竹筴鱼的时空分布
图3(a)、图4(a)为智利竹筴鱼在纬度上的时空分布,图3(b)、图4(b)为智利竹筴鱼在经度上的时空分布,从中可以看出,智利竹筴鱼不同渔区优势叉长组中叉长平均值在纬度方向上呈由北向南增大趋势,经度方向上呈由东向西增大趋势,表明智利竹筴鱼在洄游过程中同一经度上较大个体鱼位于鱼群的南部,同一纬度上较大个体鱼位于鱼群的西部。
从图3可见,2000年5—6月、2001年7—10月、2002年8—9月、2003年1—2月随着时间的变化,智利竹筴鱼作业海区在纬度上按照南-北-南方向进行移动,经度上按照东-西方向进行移动,其优势叉长组中叉长平均值也随时间变化呈现递增的趋势,表明智利竹筴鱼在30°~42°S、78°~87°W海域内按照东南-西北-西南方向进行洄游。从图4可见,2007年5—8月、2008年5—7月、2009年8—10月随着时间的变化,智利竹筴鱼作业海区在纬度上按照由南向北方向移动,经度上按照由东向西移动,优势叉长组中叉长平均值同样随时间的变化呈现递增趋势。由此认为,智利竹筴鱼在32°~48°S、80°~110°W海域内按照东南-西北方向进行洄游。从年份来看,智利竹筴鱼优势叉长组中叉长平均值随着年份的增加一直呈现递增趋势。说明在2000—2003、2007—2009年间东南太平洋智利竹筴鱼的补充群体较少,智利竹筴鱼资源存在过度捕捞现象。

图3 2000—2003年智利竹筴鱼时空分布Fig.3 Spatio-temporal distribution and range of Chilean jack mackerel from 2000 to 2003

图4 2007—2009年智利竹筴鱼时空分布Fig.4 Spatio-temporal distribution and range of Chilean jack mackerel from 2007 to 2009
2.3 智利竹筴鱼的洄游路线
张敏等[22]认为,1—2月的作业海区中37°~39°S、85°~88°W海域存在一个产卵场。本研究中产卵场及以东海域智利竹筴鱼优势叉长组主要以2~4龄为主(图5(a)),产卵场以西海域优势叉长组主要以3~5龄为主(图5(b))。根据本研究结果绘制的智利竹筴鱼洄游路线如图6所示。
图5 优势叉长范围Fig.5 Range of dominant fork length

图6 智利竹筴鱼的洄游路线图Fig.6 Migratory routine of Chilean jack mackerel
产卵场及以东海域:优势叉长组为2~4龄的智利竹筴鱼于5月份到达39.5°S、78°W附近海域后向西北方向进行越冬前的索饵洄游;6—8月鱼群在35°~38.5°S、80°~83°W海域继续向西北方向进行越冬洄游至35°S、83°W附近海域;春季鱼群继续向西北方向移动,形成越冬后的索饵洄游,于10月到达33°S、85°W附近海域。
产卵场以西海域:优势叉长组为3~5龄的智利竹筴鱼于5月到达41°S、87°W附近海域后向西北方向进行越冬前的索饵洄游;6—8月鱼群在36.5°~40°S、90~95°W海域继续向西北方向进行越冬洄游至36.5°S、95°W附近海域;春季鱼群继续向西北方向移动,形成越冬后的索饵洄游,于10月到达35°S、103°W附近海域。
2.4 洄游路线的验证
从图7可见,用神经网络模型预测智利竹筴鱼叉长,计算2016年3—10月实际叉长在预测优势叉长范围内的比值分别为31%、37%、47%、46%、42%、34%、35%、33%,5—7月的比值明显高于其他月份。

图7 实际叉长在预测优势叉长内的占比(2016年)Fig.7 Proportion of actual fork length in the predicted dominant fork length(2016)
3 讨论
3.1 洄游路线的影响因素分析
智利竹筴鱼渔场主要受西风漂流和秘鲁寒流两股海流的影响[22]。其中,西风漂流主要来自南大洋的冷水海流[25],因此,水温这一环境因子是西风漂流强弱的最重要指标。在所有环境因子中,学者一致认为,温度是影响智利竹筴鱼洄游最重要的因素[10,26-27]。研究表明,东南太平洋南部为西风漂流带来的冷水区域,北部为逆赤道暖流控制的暖水区域,智利竹筴鱼主要生活在冷暖水交汇处的混合区内[26]。李显森等[7]认为,智利竹筴鱼鱼群冬春季节向北方向进行洄游,夏秋季节向南方向进行洄游,与本文中5月份向西北方向洄游,10月份后折向西南方向进行洄游的结论相符。分析其原因,可能是冬春季节西风漂流带来的南极冷水势力加强致使海表温度降低,混合区向北移动,鱼群向北洄游;夏秋季节冷水势力减弱,混合区向南移动,鱼群向南洄游。牛明香等[26]认为,随着年龄的增长,智利竹筴鱼鱼群逆向沿西风漂流由东向西进行洄游,苏联学者认为,智利竹筴鱼在智利外海沿着35°~45°S的条带状洄游通道洄游至新西兰[22],这与本研究中智利竹筴鱼随时间推移以S形向西洄游的结果一致。分析原因可能是秘鲁寒流是来自高纬度海域的涌升流,能将海底大量的营养盐输送到海洋中上层,并促使大量的浮游生物繁衍进而为中上层鱼类提供丰富的饵料,从而控制着智利竹筴鱼在东西方向上的分布[26]。
3.2 智利竹筴鱼洄游路线分析
本研究中,1—2月的作业海区37°~39°S、85°~88°W为智利竹筴鱼的产卵场(图6)。相关学者研究认为,智利竹筴鱼幼鱼在秘鲁和智利沿岸海域进行南北洄游,成年后向西洄游[25-26,28-29],故笔者认为,公海智利竹筴鱼鱼群来自智利和秘鲁沿岸海域。本研究显示,鱼群进入公海后,以2~4龄为优势叉长的智利竹筴鱼于5月在39.5°S、78°W附近海域,并开始集群[21]向西北方向索饵洄游,6月鱼群进入索饵洄游的中期[26]并到达39°S、80°W附近海域,7月鱼群继续向西北方向洄游到达38°S、80.5°W附近海域。5—7月智利竹筴鱼的洄游距离并不大,分析认为,可能是由于这3个月的渔场空间分布重叠形成了较为稳定的渔场[26],因而导致鱼群的洄游距离较短。8月鱼群到达35°S、83°W附近海域,9—10月鱼群继续向西北方向进行索饵洄游,索饵前锋到达33°S、85°W附近海域。牛明香等[26]认为,智利竹筴鱼于10月到达最北端并向西进行洄游,因而作者推测10月后鱼群折向西南方向进行产卵前的分散索饵洄游直至产卵场。李显森等[7]研究显示,冬春季节鱼群向北洄游,与本研究中从5月份开始鱼群向西北方向洄游的结果一致。此外,有研究表明,智利竹筴鱼的首次产卵群体主要以3龄鱼为主,产卵盛期为12月至翌年2月[14],这也印证了此次推测。
杨嘉樑等[25]研究认为,智利竹筴鱼鱼群3月产卵后的聚集度最低,开始分散洄游至翌年5月再次集群。据本研究中翌年5月洄游站点和产卵场的位置推测,翌年3—4月鱼群自产卵场向西南方向进行分散索饵洄游,于翌年5月到达41°S、87°W附近海区,并再次集群向西北方向进行索饵洄游,翌年6月鱼群到达40°S、90°W附近海域,翌年7月鱼群到达39°S、92°W附近海区,翌年8月到达36.5°S、95°W附近海区,翌年9—10月鱼群分散索饵,索饵前锋到达35°S、103°W附近海域,翌年10月鱼群则再次折向西南方向进行产卵前的索饵洄游,并于下一年1—2月到达105°~125°W、35°~48°S海域进行产卵洄游[22]。李显森等[14]研究表明,鱼群第2次性成熟后,4~5龄鱼在南太平洋中部产卵场35°~38°S、105°~125°W海区产卵,印证了本研究结论。
邹晓荣[30]认为,较大个体智利竹筴鱼分布于偏南、偏西水域,较小个体鱼分布于偏东、偏北水域,与本研究结论一致。这可能是由于大个体智利竹筴鱼鱼群最适觅食海区温度相较小个体鱼群温度低[25]所致,而南部为西风漂流带来的冷水,北部为赤道暖流控制的暖水团,因此,南部海区温度较北部海区温度偏低。经度方向上,主要是由于智利竹筴鱼会逆西风海流向西洄游[25],致使大个体鱼偏西。
本研究表明,智利竹筴鱼在向西洄游过程中,冬春季节鱼群向西北方向进行洄游,夏秋季节鱼群向西南方向进行洄游,洄游路线在月际间呈现大S状路线向西洄游,月内则呈小S状路线向西洄游,此结果与张敏等[22]的研究结果一致。由于智利竹筴鱼具有分批产卵、分批洄游的特性[9],且每年的环境、种群资源丰度等诸多因素不固定,因此,其具体某个月份的洄游海区可能并不固定。有文献表明,智利竹筴鱼很可能在成熟以后永不返回东部海区[22]。
3.3 未来研究展望
目前,国内外对智利竹筴鱼的研究多集中于渔场、渔情预报、栖息地指数等资源分布方面,而对智利竹筴鱼的洄游路线提及较少,因此,建议后期在以下方面进一步开展研究。
1)因数据限制,本研究中在时间上采用月份,空间采用1°的矩形海区统计智利竹筴鱼鱼群的优势叉长,并对洄游路线进行分析。如果获取全年份数据,以周为时间单位,以30′的矩形海区统计鱼群优势叉长,会得到更为详细的洄游路线。
2)智利竹筴鱼渔业与海洋环境要素研究多停留在海表温度、盐度、叶绿素a浓度等方面,而气候变化对智利竹筴鱼渔业的影响研究较少,以后有必要开展气候变化对智利竹筴鱼渔业影响研究,从而为智利竹筴鱼渔业的可持续发展提供理论指导。
3)渔业管理部门有必要加大对中国智利竹筴鱼不同时间、不同作业海区的渔业数据的统计,并结合耳石微化学与海洋环境的关系,对智利竹筴鱼的洄游路线进一步进行推测,从而掌握智利竹筴鱼在不同生活史阶段更为具体的洄游路线。
4)随着全球环境的不断变化,海洋环境的理化特性也产生了新的改变,需要开展智利竹筴鱼鱼卵仔鱼分布与环境因子关系的研究,探究在不同海洋环境下智利竹筴鱼产卵场空间分布的变迁,并为其产卵场提出具体保护措施和管理建议。
4 结论
1)利用中国2000—2003、2007—2009、2016年东南太平洋智利竹筴鱼大型拖网渔捞日志及生物学数据,绘制了鱼群在30°~50°S、75°~110°W海区的洄游路线。研究发现,智利竹筴鱼鱼群随季节的变化呈现南北洄游,其中,温度是影响智利竹筴鱼洄游最重要的因素。
2)智利竹筴鱼的洄游主要受西风漂流和秘鲁寒流两股海流的影响,其中,成年鱼类随着鱼龄的增大呈逆西风漂流的流向向西洄游,较大个体竹筴鱼偏向于西南方向,整体呈现S形向西洄游。
3)结合环境因子利用神经网络模型对2016年3—10月叉长进行预测,利用各月预测的叉长统计其优势叉长组范围,计算在该范围内实际叉长量占当月总叉长量的比值并进行验证,验证结果显示,2016年3—10月比值分别为31%、37%、47%、46%、42%、34%、35%、33%。
