高地钩叶藤材薄壁细胞壁主要化学成分的微区分布1)
2022-01-06张菲菲何伟汪佑宏马建锋
张菲菲 何伟 汪佑宏 马建锋
(安徽农业大学,合肥,230036) (国际竹藤中心)
棕榈藤为棕榈科(Palmae)省藤亚科(Calamoideae)类植物,天然分布于东半球的热带地区及邻近区域。世界棕榈藤总共有13个属600~700种,其中我国自然分布有4属(黄藤属、省藤属、钩叶藤属和多鳞藤属)37种5变种,但有较高经济价值的不到20种[1]。藤茎俗称藤条,成熟后呈奶黄、乳白等颜色,除基部外被带刺叶鞘所覆盖;长度从几米至百余米、直径从约3~80 mm甚至100 mm不等。藤条为仅次于木竹材的、具有多种用途的重要林产品,被广泛用于制造桌、椅、沙发、床等藤制家具,以及各种藤编小饰品等,具有很高的经济价值和开发前景。
棕榈藤藤茎主要有维管束及环绕四周的薄壁组织构成,而薄壁细胞的大量存在,赋予了棕榈藤藤茎优良的柔韧性[2-3],有利于棕榈藤的弯曲加工和编织利用。而目前,关于棕榈藤的研究报道多见解剖构造特征[4-5],宏观物理力学性质[6-8]及化学性质方面[7,9]。当然,对棕榈藤材细胞壁木质素的微区分布也略有研究,如运用可见分光光度计对黄藤材导管分子[10]、纤维[11]及薄壁细胞[12]等细胞壁的木质素微区分布进行了探索。此外,刘杏娥等[13]结合共聚焦显微荧光和拉曼光谱成像技术系统,对黄藤材藤茎中不同类型细胞以及同一细胞不同形态区域的木质素化学特点进行了分析。
就棕榈藤材而言,关于薄壁组织细胞壁主要成分的微区分布尚未见报道。为构建棕榈藤材薄壁细胞壁结构模型、加大棕榈藤材的用途和使用率,对棕榈藤材薄壁细胞壁主要成分的微区分布进行研究,还有待进一步加强。
1 材料与方法
高地钩叶藤(PlectocomiahimalayanaGriff.)属钩叶藤属,主要分布于我国云南西部、南部地区海拔1 450~1 800 m的竹林、山地常绿阔叶林中,大径、攀援、丛生,一般仅用于编织较粗糙的藤器或扎栏[14]。
取生长适中、健康的高地钩叶藤1根,在距地面2 m处截取高约1 cm的试样,软化包埋处理后,用旋转式切片机切取厚10 μm的完好的横切面切片5片左右,室温下用0.2 mol·L-1NaBH4溶液浸泡5~6 h,然后用蒸馏水洗净并浸泡在蒸馏水中备用。LabRam XploRA显微共聚焦拉曼光谱仪,KD1508A型轮转式切片机等。
将存放在蒸馏水中的切片取出1片,固定在载玻片上,置于拉曼显微镜下,分别在藤皮、藤中及藤芯处各选择1个完整的薄壁细胞、每个细胞壁上选择至少3个点,利用LabRam XploRA显微共聚焦拉曼光谱仪,采用逐点扫描显微探针成像方法获取光谱数据集,再利用LabSpec5软件进行处理。其中扫描步长为0.8 μm,激发波长为532 nm,功率8 mW,单点采集时间1 s,狭缝宽度100 μm,孔隙大小为300 μm,拉曼光谱测试范围为250~3 200 cm-1。为获得较高的空间分辨率,采用了100倍油镜(MPlan 100×,oil,NA=1.35),计算机控制平台的同步移动、对3个薄壁细胞至少9个点数据进行采集和数据处理。
高地钩叶藤薄壁细胞次生壁一般3~7层,壁层厚度285~900 nm,层与层之间有空隙。试验中薄壁细胞次生壁为7层,依据壁层厚度、拉曼光斑大小及扫描步长等因素,由外向里将薄壁细胞次生壁依次分成SⅠ(1~2层)、SⅡ(3~5层)和SⅢ(6~7层)3部分进行试验。
2 结果与分析
2.1 薄壁细胞纤维素与木质素的微区分布线谱
图1为薄壁细胞SⅡ(次生壁中部)的平均拉曼光谱,其中纤维素的主要特征峰为2 890、1 324、1 114、1 090和378 cm-1;木质素的主要特征峰为1 680和1 601 cm-1。拉曼位移2 890 cm-1归属于纤维素C—H和CH2的对称伸缩振动,1 601 cm-1是苯环的对称伸缩振动,378 cm-1归属于纤维素β—D葡萄糖甘键。拉曼位移1 680 cm-1归属于松柏醇或紫丁香醇和松柏醛或紫丁香醛结构,1 601 cm-1归属于苯环的对称伸缩振动。拉曼位移1 090 cm-1是重原子(CC和CO)的伸缩振动(见表1)。
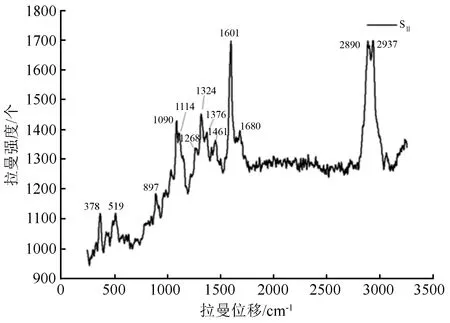
图1 薄壁细胞壁SⅡ层平均拉曼光谱
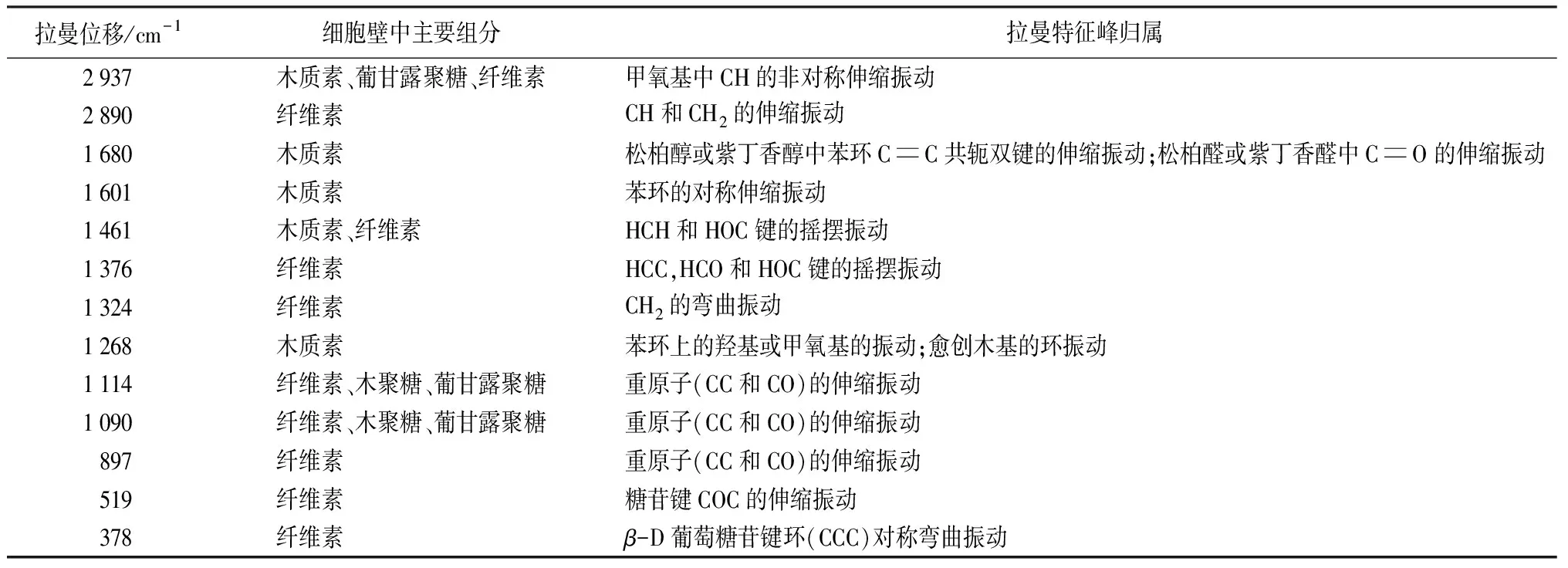
表1 薄壁细胞壁SⅡ层主要组分的平均拉曼光谱分布[18-19]
图2(a)为薄壁细胞壁5个不同形态区,即细胞角隅处(CC),复合胞间层(CML),次生壁外部(SⅠ),次生壁中部(SⅡ)及次生壁内部(SⅢ)的平均光谱图,线谱范围为900~3 200 cm-1。从图中可以看出CC中2 937 cm-1特征峰的信号很强,而2 890 cm-1处的信号几乎被其覆盖,CC形态区拉曼信号谱图整个基线远高于其他形态区。CML、SⅠ、SⅡ和SⅢ处2 937 cm-1特征峰的信号均比2 890 cm-1处信号强。
图2(a)经谱线分析处理得到图2(b)和图2(c)。图2(b)比较了CC、CML和SⅡ3个形态区的拉曼光谱,从中可以看出2 890 cm-1特征峰处的拉曼强度为CC处最小,说明CC处纤维素的集团数量比最低;CML与SⅡ处在2 890 cm-1特征峰处的拉曼强度没有差别。另一特征峰1 601 cm-1处的拉曼强度由大到小为CC、CML、SⅡ,说明木质素集团数量比又大到小为CC、CML、SⅡ,这与Cybulska J et al.[15]对苹果组织研究的结果完全一致,与刘杏娥等[13]、汪佑宏等[11]研究的黄藤材纤维细胞壁及薄壁细胞壁[12]上木质素的分布也相似,单就细胞角隅处木质素集团数量比最高的观点而言,与Richter S et al.[16]及Sun L et al.[17]的观点也是相同的。
图2(c)比较了SⅠ、SⅡ和SⅢ3个形态区的拉曼光谱,从中可以看出3个形态区在2 890 cm-1特征峰和1 601 cm-1特征峰两处的拉曼强度均差别不大,说明这3个形态区纤维素的集团数量比与木质素集团数量比区别较小。而黄藤材导管分子细胞次生壁上木质素由外向内呈“S型”分布,但差异不大[10];原因可能是由于薄壁细胞次生壁太薄,拉曼光斑直径大于次生壁各壁层厚度,导致细胞壁层间信号重合,以及壁层间存在空隙共同所致。
2.2 薄壁细胞拉曼成像
在显微镜下的试材横切面上,选择适合的薄壁细胞进行拉曼扫描并成像。由于纤维素的拉曼光谱信噪比在2 890 cm-1处最大,能够得到较清晰的分布图像,而木质素的主要特征峰在1 601 cm-1处具有较好的信噪比,因此本研究选择在2 780~3 060 cm-1(用C—H以及C—H2伸缩振动的空间分布规律来代表薄壁细胞中纤维素的分布),1 550~1 640 cm-1(用苯环的对称伸缩振动的空间分布规律来代表薄壁细胞中木质素的分布)以及1 026~1 195 cm-1(C—C和C—O—C伸缩振动的空间分布规律来代表薄壁细胞中半纤维素的分布)3个振动区分别进行拉曼成像,结果如图3所示。图3(a)为显微镜下选择的细胞区域,(b)、(c)、(d)分别为薄壁细胞中纤维素、木质素、半纤维素的空间分布图;其中(b)的成像振动区主要特征峰是2 890 cm-1,是C—H以及C—H2伸缩振动,(c)的成像振动区主要特征峰是1 601 cm-1,是苯环的对称伸缩振动,而(d)的成像振动区主要特征峰是1 090 cm-1,是重原子(CC和CO)的伸缩振动,对应的主要组分是纤维素、木聚糖、葡甘露聚糖。
如图3(b)所示,在薄壁细胞中可以明显的看出,纤维素在次生壁层(S)上的拉曼信号最强(对比度最亮),说明此处纤维素的集团数量比最高;复合胞间层(CML)的纤维素集团数量比次之;而细胞角隅(CC)处的纤维素的拉曼信号非常弱(对比度很暗),说明CC处纤维素的集团数量比很低。此外,还能明显的看出细胞壁的水平方向的拉曼信号对比度比垂直方向暗,说明该薄壁细胞壁上的纤维素集团数量比水平方向低于垂直方向。
图3(c)所示,CC处木质素拉曼信号最强(对比度最亮),CML处木质素拉曼信号强度仅次于CC处,而S上的木质素拉曼信号强度比较弱,说明CC处的木质素集团数量比最高,CML次之,S上的木质素集团数量比较低,这与前文中对薄壁细胞壁木质素的微区分布线谱分析结果是一致的。
图3(d)中可以看出S层上拉曼信号对比度较亮,CML次之,CC处的对比度很暗,说明S层上的半纤维素集团数量比最高,CML次之,CC处集团数量比最低。此外,成像图中薄壁细胞壁水平方向拉曼信号对比度明显比垂直方向亮,说明半纤维素集团数量比水平方向高于于垂直方向,与纤维素的情况相反。
3 结论与讨论
高地钩叶藤薄壁细胞壁上纤维素、半纤维素集团数量比由高至低依次是次生壁(S)、复合胞间层(CML)、细胞角隅(CC)处,而木质素集团数量比恰恰相反。薄壁细胞的次生壁外部(SⅠ)、中部(SⅡ)和内部(SⅢ)3个形态区纤维素集团数量比、木质素集团数量比差别均较小;CML与SⅡ处纤维素集团数量比没有差别,但SⅡ木质素集团数量比小于CML。此外,薄壁细胞壁的纤维素集团数量比水平方向低于垂直方向,而半维素集团数量比水平方向高于于垂直方向,与纤维素的情况相反。
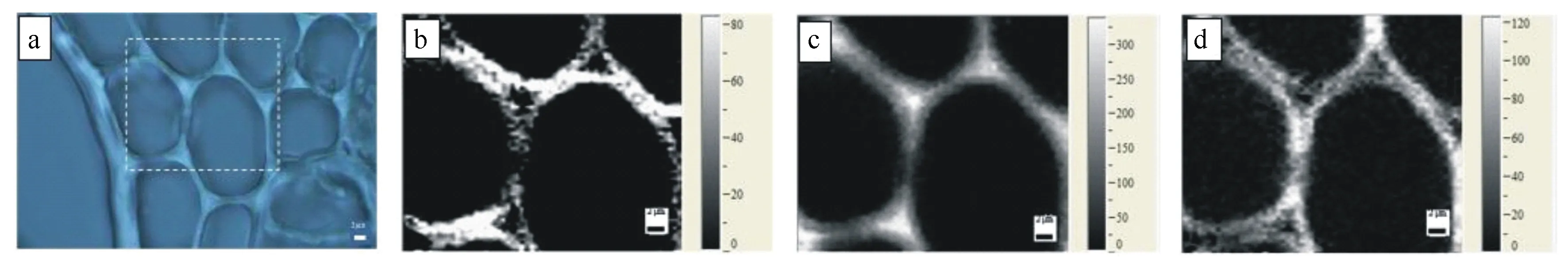
(a)与薄壁细胞壁中纤维素;(b)木质素;(c)半纤维素;(d)分布图像。
