NLRP3 炎性小体在抗新孢子虫感染中的作用及调控机制
2022-01-06王安琪孔琳延边大学农学院动物医学系133002
王安琪 孔琳/延边大学农学院动物医学系 133002
炎症小体(inflammasome)存在于细胞内,是一种由很多蛋白质组成的复合物,分子量很大,约为700 KDa,是天然免疫系统的重要组成部分。目前的许多相关研究都表明,炎症小体参与了宿主的防御反应,其中,核苷酸结合寡聚化结构域样受体3(nucleotide binding oligomerzation domain like receptorprotein3,NLRP3)炎症小体,参与多种抗寄生虫感染的作用及调控机制,已成为多个生物学领域的研究热点。
新孢子虫是一种细胞内寄生虫,牛、羊等一些中间宿主感染新孢子虫后会引起孕畜流产,对养牛业的繁殖具有巨大的冲击,同时也带来了极大的经济损失。为进一步探索NLRP3 介导的炎性小体反应,本研究利用NLRP3 基因缺失小鼠深入研究NLRP3 炎性小体在抗新孢子虫感染中的作用,为继续探索NLRP3 基因的作用及机制奠定基础。
1 材料
1.1 细胞与虫株新孢子虫速殖子和Vero 均为本实验室冻存。
1.2 实验动物C57BL/6 雌性小鼠,C57BL/6 雌性NLRP3,ASC 及caspase-1/11 基因缺失小鼠,均为6~8 周龄,体重20+4g,购自于长春亿斯实验动物技术有限公司。饲养环境:将小鼠置于25℃,湿度50%,独立通风的环境中喂养,定时定量给予鼠料并自由饮水。
1.3 试剂与药品RPIM-1640 培养基、胎血清牛、DMEM培养基均购自吉林省盟创生物科技有限公司;0.22μm PVDF膜购自北京世纪奥科生物技术有限公司;小鼠IL-1β,IL-18,IFN-γ,ELISA 试剂盒购自Sigma 公司。
2 方法
2.1 新孢子虫的培养和纯化参考杨慧珍的方法,从-80℃冰箱取出装有虫体的冻存管,立即放入37℃的水浴中,快速晃动,待虫体悬液完全融解后,移入已经长成单层的Vero 细胞的培养瓶中,通过显微镜,观察细胞和虫体的生长状态,间隔12h 换液一次,根据细胞状态和虫体数量确定传代时间。传代培养20d 后,获得足量的、生长状态良好的虫体备用。
2.2 分离Vero 细胞中的新孢子虫虫体参考玄学南的方法,在虫体生长达到旺盛时,倒掉旧的培养液,对细胞进行润洗后,用细胞刮刮下细胞,通过5.0μm 孔径的微孔滤膜,滤液收于50mL 离心管,将滤过的虫体悬液2000rpm 离心10min,弃去上清液,加入50mL 不含血清培养液仔细重悬虫体,在显微镜下,用细胞计数板计数虫体个数,保存备用。
2.3 虫体感染小鼠模型的建立用500μl PBS 重悬2×107新孢子虫速殖子,然后进行小鼠的腹腔接种,感染10d 后,对组织进行采样检测;用500μl PBS 重悬1×107新孢子虫速殖子,感染15d 后,对组织进行采样检测。
2.4 虫量检测将小鼠处死之后,用1mL 冷PBS 反复充分的冲洗小鼠腹腔,将腹腔内的灌洗液全部吸出,离心后,-20℃保存待用;分别取小鼠的心、肺和脑组织,加入液氮充分研磨,取100~200mg,用DNA 提取试剂盒提取基因组DNA,进行浓度测定后,-20℃保存待用;最后用绝对荧光定量PCR 检测细胞组织中新孢子虫的数量。
2.5 ELISA 检测感染后的小鼠眼球采血,4℃静置12h,将析出的血清8000g 离心10min,收集上清液,-20℃保存待用;将获得的小鼠腹腔灌洗液离心取上清液,-20℃保存待用。稀释样品,用ELISA 试剂盒进行检测。
2.6 数据处理用 Graphpad 软件作图,Student’s t 检验用于统计学分析。
3 结果
3.1 宿主被新孢子虫感染后NLR P3 炎症小体的保护作用为进一步了解新孢子虫感染诱导的NLRP3 炎症小体反应对宿主产生致病作用还是保护作用,将采用2×107新孢子虫速殖子对小鼠进行持续30d 的腹腔感染。结果发现在感染第10d 时,对IL-1β 的分泌量进行检测,WT 及三种基因缺失小鼠的血清中均未见IL-1β(图1A),但发现WT 小鼠血清中有较高水平的IL-18,而在ASC-/-,Caspase-1/11-/-小鼠中IL-18 的含量显著减少,NLIP3-/-小鼠中IL-18 的分泌也存在降低趋势,但无明显差异(图1B)。对小鼠的脑组织、心脏、肺组织的新孢子虫数量进行检测发现,WT小鼠与NLIP 3-/-小鼠的虫数无明显差异,ASC-/-小鼠Caspase-1/11-/-小鼠脑组织中的数量与WT 小鼠相比明显增多(图1C);ASC-/-小鼠和Caspase-1/11-/-小鼠心脏和肺组织中的数量远远大于WT 小鼠,但ASC-/-小鼠和Caspase-1/11-/-的数量没有明显差异(图1D,E)。
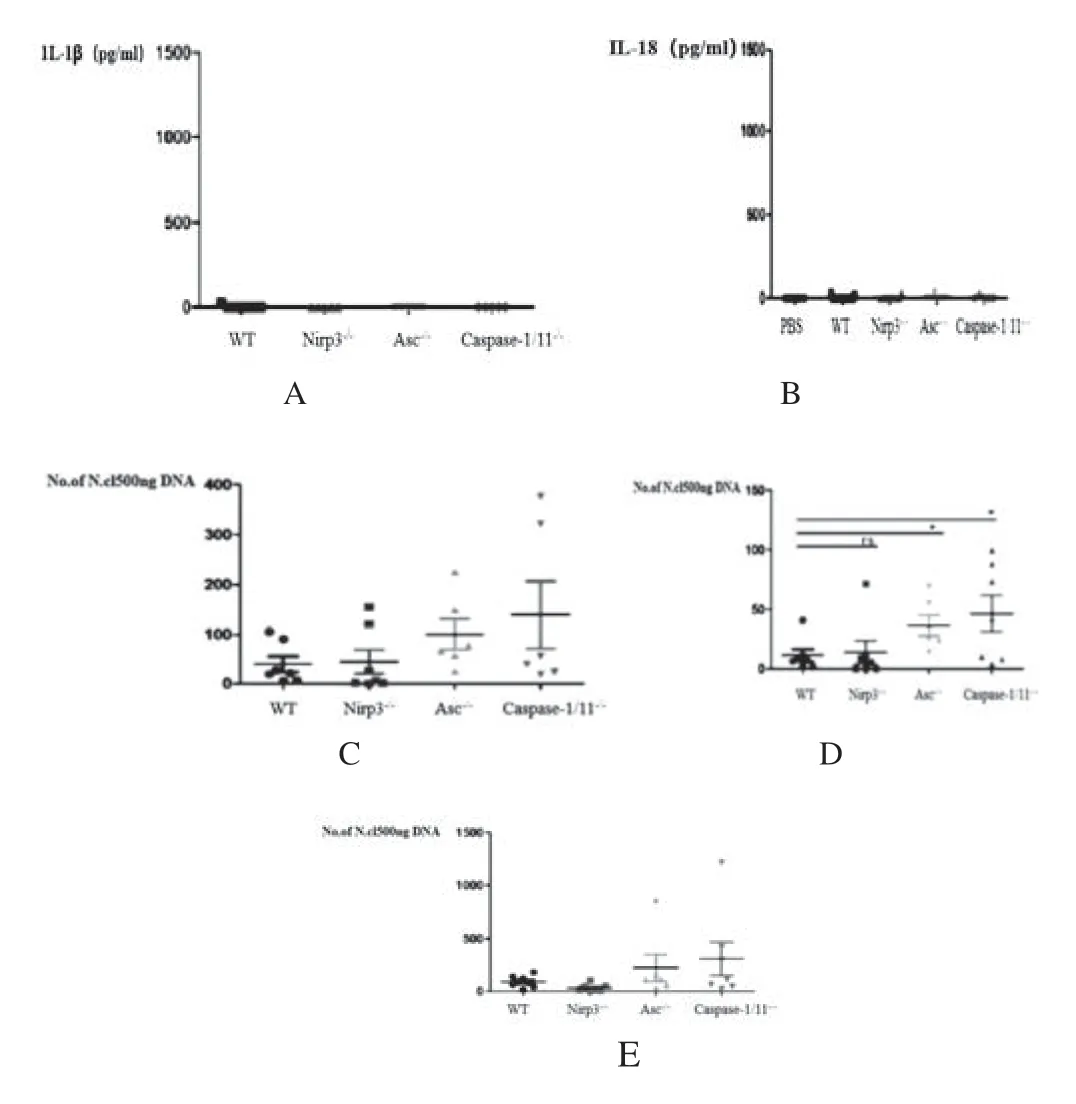
图1 NLRP3,ASC 和 caspase-1/11 在新孢子虫感染中的作用
3.2 在新孢子虫早期感染中NLR P3 炎症小体介导细胞因子分泌宿主在感染新孢子虫之后,NLRP3,ASC 和Caspase-1/11 可以诱导分泌IL-18,需要继续探讨这三种分子是否对其它细胞因子的分泌造成影响。采取1×107新孢子虫速殖子对小鼠进行持续15d 的腹腔感染,再利用ELISA 方法检测血清中的IL-1β,IL-18,IFN-γ 和TNF-α,结果发现新孢子虫的感染无法介导NLIP3-/-,ASC-/-,Caspase-1/11-/-小鼠及WT 小鼠分泌IL-1β(图2A);新孢子虫感染可以介导WT 小鼠分泌较高的IL-18,但其它三种基因缺失小鼠分泌的IL-18 数量显著降低(图2B)。并且三种基因缺失小鼠分泌IFN-γ 的数量也较WT 小鼠明显降低(图2C)。四种小鼠所分泌的TNF-α 数量无显著差异(图2D)。
4 讨论
NLRP3 炎症小体需要双重信号的激活,第一种信号是由NF-KB 信号通路激活的,激活后,pro-IL-1β 和 pro-IL-18表达上调;第二种信号是由各种 DAMPs 或 PAMPs 激活的,与此同时,可以介导 IL-1β/IL-18 成熟分泌和细胞焦亡。为了进一步探讨小鼠感染新孢子虫后,NLRP3 炎症小体在机体中的作用,本研究利用NLRP3,ASC 及caspase-1/11 缺失小鼠开展了一系列的试验。

图3.2 NLRP3,ASC 和 caspase-1/11在诱导促炎细胞因子中的作用
NLRP3,ASC 及caspase-1/11 在对宿主的保护过程及抵抗新孢子虫的传播中发挥着各自的作用。其中,NLRP3和ASC 主要的作用是控制宿主对新孢子虫的易感性,而caspase-11 则会影响caspase-1 对新孢子虫感染的易感性。缺失NLRP3,Asc 和 caspase-1/11 的小鼠感染新孢子虫后,IL-1β 的成熟分泌及caspase-1 活化明显减少甚至消失,说明新孢子虫激活的炎症小体反应依赖NLRP3,Asc 和 caspase-1/11。
机体感染新孢子虫之后,可以激发机体的Th1 免疫应答,可以介导IFN-γ 和IL-12 的大量分泌,而且这些细胞因子在清除新孢子虫和抗新孢子虫的过程中发挥非常重要的作用。将IFN-γ 缺失的小鼠感染新孢子虫后,小鼠不能使T 细胞分化,且NO 分泌减少,同时小鼠的存活率显著下降。在本研究中,NLRP3,Asc 和 caspase-1/11 缺失小鼠感染新孢子虫后,测量血清中的IFN-γ,结果显示,IFN-γ 分泌明显减少,推测这或许和炎症小体介导分泌的IL-18 有关。
5 结论
通过以上的数据结果可以发现,由新孢子虫感染所分泌的IL-18 部分依赖NLRP3,主要依赖ASC 和Caspase-1/11;在清除新孢子虫与保护宿主时,这三种分子发挥不同的作用;NLRP3,ASC,Caspase-1/11 在新孢子虫早期感染中介导IFN-γ 的分泌,对宿主抗新孢子虫的Th1 应答有促进作用。
