二化螟CsCncC和CsKeap1基因的克隆与mRNA表达分析
2022-01-05季彩宏孟祥坤王建军
季彩宏,张 楠,孟祥坤,王建军*
(1. 扬州大学园艺与植物保护学院,江苏扬州 225009;2. 扬州市职业大学园林园艺学院,江苏扬州 225009)
在哺乳动物中,Nrf2-Keap1通路主要调控细胞氧化还原平衡,参与细胞抗氧化和Ⅱ相解毒反应(Kensleretal.,2007;Lobodaetal.,2016)。在该信号通路中,属于Cnc-bZIP (cap ‘n’ collar/basic region leucine zipper)家族的Nrf2(核因子E2相关因子2,nuclear factor erythroid 2-related factor 2),含有6个环氧氯丙烷(EHC,epichlorohydrin)相关蛋白同源结构域(Neh,Nrf2-EHC homology)(Neh1-Neh6),其中,氨基端Neh2功能结构域是Nrf2与Kelch样环氧氯丙烷相关蛋白-1(Keap1,Kelch-like ECH-associated protein 1)的结合区,后者在正常的生理条件下与Nrf2结合并将其隔离、锚定在胞浆,促进Nrf2的泛素化和蛋白酶体降解(Kobayashietal.,2006; Tianetal.,2018)。
害虫抗药性问题是害虫综合治理中最重要的问题之一,明确抗药性机制是害虫抗药性治理的基础。昆虫中第一个鉴定的Nrf2同源基因是黑腹果蝇Drosophilamelanogaster的CncC基因(Mohleretal.,1991;Sykiotis and Bohmann,2008),可调控36种细胞色素P450、17种谷胱甘肽S-转移酶(GST)、6种尿苷二磷酸葡萄糖醛酸基转移酶(UGT)和55种ABC转运蛋白基因的表达(Misraetal.,2011)。进一步对DDT抗性品系91R和RDDTR的研究发现,敲减CncC或者过表达Keap1都可导致与DDT抗性相关的细胞色素P450基因Cyp6a8和Cyp6a2的表达下调(Misraetal.,2013)。对赤拟谷盗Triboliumcastaneum的研究发现,CncC可调控与溴氰菊酯抗性相关的8个CYP6BQ基因的表达(Kalsi and Palli, 2015)。对棉蚜Aphisgossypii的研究发现,CncC可调控与棉酚耐性相关的CYP6DA2基因表达(Pengetal., 2016)。对朱砂叶螨Tetranychuscinnabarinus的研究发现,CncC可调控CYP389B1、CYP391A1和CYP392A28等与甲氰菊酯抗性相关基因的表达(Shietal.,2017)。最近对马铃薯甲虫Leptinotarsadecemlineata的研究发现,CncC可调控CYP9Z25、CYP9Z29、CYP6BJa/b和CYP6BJ1v1等4个与马铃薯次生物质解毒代谢和吡虫啉抗性相关P450基因的表达(Kalsi and Palli, 2017)。这些研究表明,昆虫Nrf2-Keap1信号通路可调控I相、II相和III解毒代谢基因的表达,在昆虫抗药性进化过程中具有重要作用。
二化螟Chilosuppressalis(Lepidoptera: Pyralidae)是水稻作物上一种主要的害虫,目前,对二化螟的防治主要依赖于化学农药,但随着化学农药的大量使用,二化螟先后对杀虫单、三唑磷、毒死蜱、氟虫腈和氯虫苯甲酰胺等杀虫剂产生了不同程度的抗药性(Suetal.,2014;常菊花,2016;Luetal.,2017;Yaoetal.,2017)。本文克隆了二化螟CsCncC和CsKeap1基因,测定了CsCncC和CsKeap1基因在二化螟不同发育阶段和不同组织的mRNA表达水平,并研究了CsCncC和CsKeap1基因对杀虫剂氯虫苯甲酰胺的诱导响应,为进一步研究二化螟对氯虫苯甲酰胺等杀虫剂的代谢抗性机制奠定基础。
1 材料与方法
1.1 供试昆虫与试剂
二化螟试虫采集于扬州市仪征市水稻田(32.39 N,119.42 E),置于恒温培养箱中,在温度28℃±1℃,相对湿度70%±5%,光周期为16 ∶8(L ∶D)条件下,用人工饲料进行饲养(Huangetal.,2016)。SMARTer RACE 5′/3′ Kit、Mighty Amp、PrimeScriptTM1stcDNA Synthesis Kit、Takara MiniBEST Universal RNA Extraction Kit、Takara MiniBEST Agarose Gel DNA Extraction Kit、PrimeScriptTMRT reagent Kit with gDNA Eraser、TB Green Premix Ex TaqTM等酶和试剂盒购买于宝生物工程(大连)有限公司(Takara);pEASY-T1克隆载体、Trans 5α感受态细胞购买于北京全式金生物技术有限公司。95%氯虫苯甲酰胺原药购自美国杜邦公司。
1.2 二化螟处理
根据前期获得的氯虫苯甲酰胺对二化螟的毒力回归线,分别配制含有LC30(0.092 mg/L)和LC70(0.47 mg/L)氯虫苯甲酰胺的人工饲料(Huangetal.,2016)。使用含有氯虫苯甲酰胺的人工饲料和对照饲料处理二化螟3龄幼虫,每个处理接入60头幼虫,重复3次,分别在处理12 h、24 h和36 h后收集样品试虫,每5头二化螟幼虫为一个样品。
1.3 二化螟CsCncC和CsKeap1的基因克隆与序列分析
根据试剂盒说明,使用Takara MiniBEST Universal RNA Extraction Kit提取二化螟总RNA,置于-80℃条件下保存备用。以提取的总RNA为模板,使用PrimeScriptTM1stcDNA Synthesis Kit试剂盒进行反转录,合成cDNA第一链模板。根据从二化螟转录组(Mengetal.,2019a)中鉴定到的CsCncC和CsKeap1序列片段,设计特异性引物(表1),用于RT-PCR扩增并测序验证。引物由上海生工生物工程技术服务有限公司合成。25 μL PCR反应体系中包含:9 μL ddH2O、12.5 μL Amp Buffer、1 μL上游/下游引物(10 μM)、0.5 μL Amp聚合酶、1 μL cDNA模板。PCR扩增条件为:98℃预变性2 min;98℃变性10 s,55℃退火15 s,72℃延伸3 min,循环37次;72℃延伸10 min。使用Takara MiniBEST Agarose Gel DNA Extraction Kit对PCR产物进行回收,并连接到pEASY-T1克隆载体上,进一步转化到Trans 5α感受态细胞中,挑取阳性克隆送至上海生工生物工程技术服务有限公司进行测序。
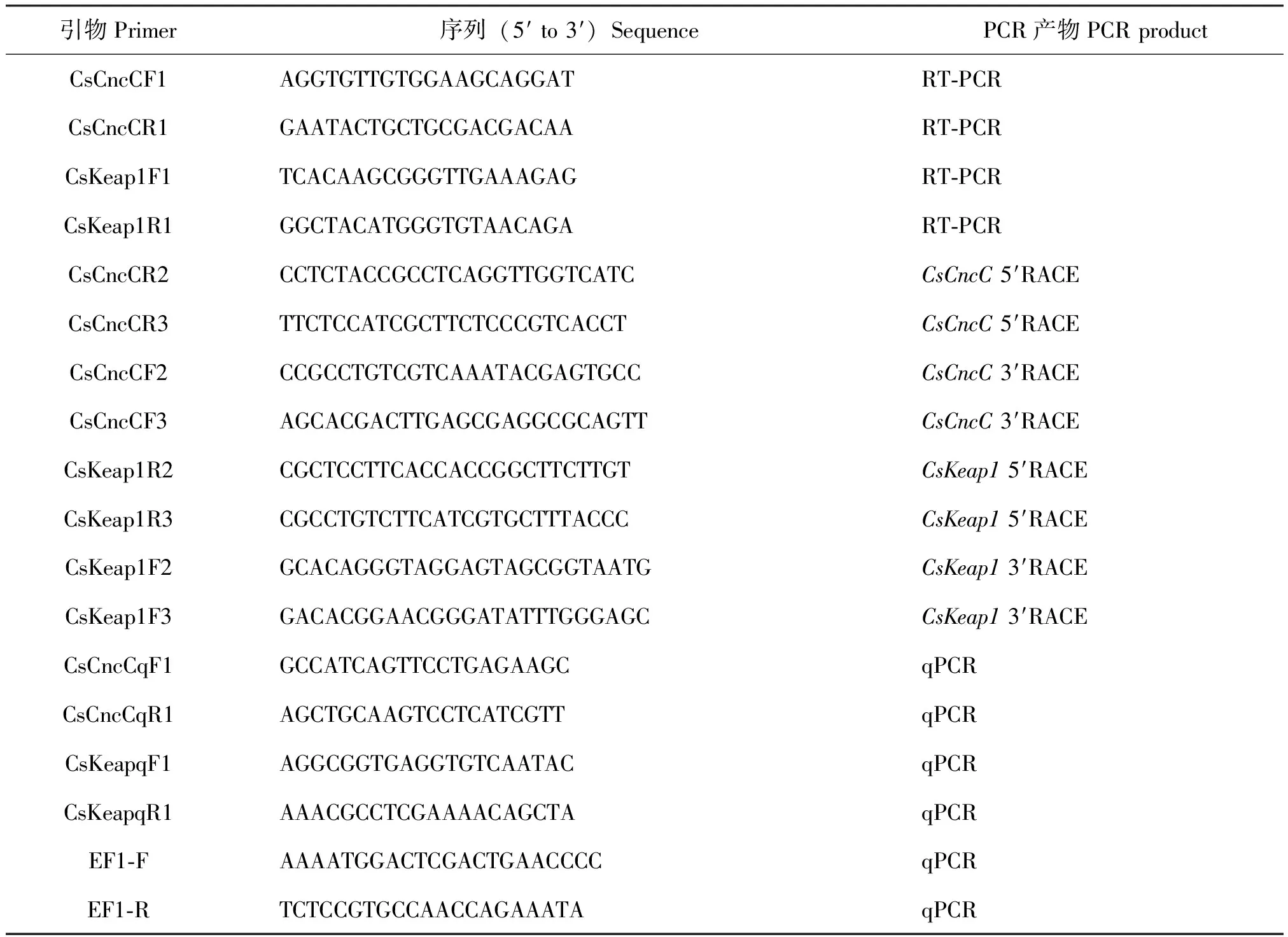
表1 本文中使用的引物
根据RT-PCR测序结果,设计特异性引物,使用SMARTer RACE 5′/3′ Kit对二化螟CsCncC和CsKeap1基因的5′和3′端进行扩增。分别使用CLUSTALW和MEGA 5.0软件对二化螟CsCncC和CsKeap1进行多重序列比对和系统发生分析。利用ExPASy在线软件(https: //web.expasy.org/protparam/)对翻译蛋白质的蛋白分子质量和等电点进行预测。通过与已发表的其它昆虫同源基因的比对,分析CsCncC和CsKeap1的保守结构域和特征性基序。
1.4 定量PCR(qPCR)检测
根据克隆获得的二化螟CsCncC和CsKeap1序列信息,设计特异性引物,使用荧光定量PCR对两个基因在二化螟不同龄期、不同组织部位中的表达量进行分析。分别收集二化螟幼虫、蛹和成虫的不同龄期试虫,并从3龄幼虫中解剖出血淋巴、神经索、脑、脂肪体、表皮、前肠、中肠、后肠和马氏管组织,从24 h雌成虫中解剖出卵巢组织。每个样品收集3个生物重复。以在二化螟中稳定表达的EF-1a为内参基因(Huietal.,2011;Mengetal.,2019b)。荧光定量PCR反应体系中含有10 μL 2×TB Green Premix Ex Taq,10 μM的上下游引物各1 μL,cDNA模板2 μL,6 μL灭菌ddH2O。PCR反应条件为95℃预变性2 min;95℃变性30 s,60℃退火30 s,40个循环;在扩增结束后进行熔解曲线分析检测扩增反应的特异性。利用2ΔΔCt方法计算计算目标基因的相对表达量,利用one-way ANOVA 进行差异显著性分析与多重比较。实验数据用平均数±标准误表示。
2 结果与分析
2.1 二化螟CsCncC基因克隆及序列分析
通过克隆获得二化螟CsCncC基因2 845 bp cDNA全长序列(GenBank登录号:MW147281),其中包含一个长度为2 160 bp的开放阅读框,一个262 bp的5′非翻译区为和一个423 bp的3′非翻译区(图1)。二化螟CsCncC基因编码719个氨基酸,蛋白质分子质量为80.59 kDa,等电点为5.87。

图1 二化螟CsCncC基因核苷酸序列及其编码的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of CsCncC in Chilo suppressalis
氨基酸多重序列比对发现,二化螟CsCncC蛋白具有Nrf2家族的典型结构特征,如含有高度保守的CNC类型的碱性亮氨酸拉链结构(basic leucine zipper,bZIP)和ETGE基序,但DLG基序变为DMG(图2)。对不同昆虫CncC的系统发生分析显示,二化螟CsCncC与鳞翅目昆虫CncC聚于同一支(图3)。

图2 二化螟CsCncC与其它昆虫CncC的氨基酸序列比对Fig.2 Multiple alignments of CncC from Chilo suppressalis and other insects
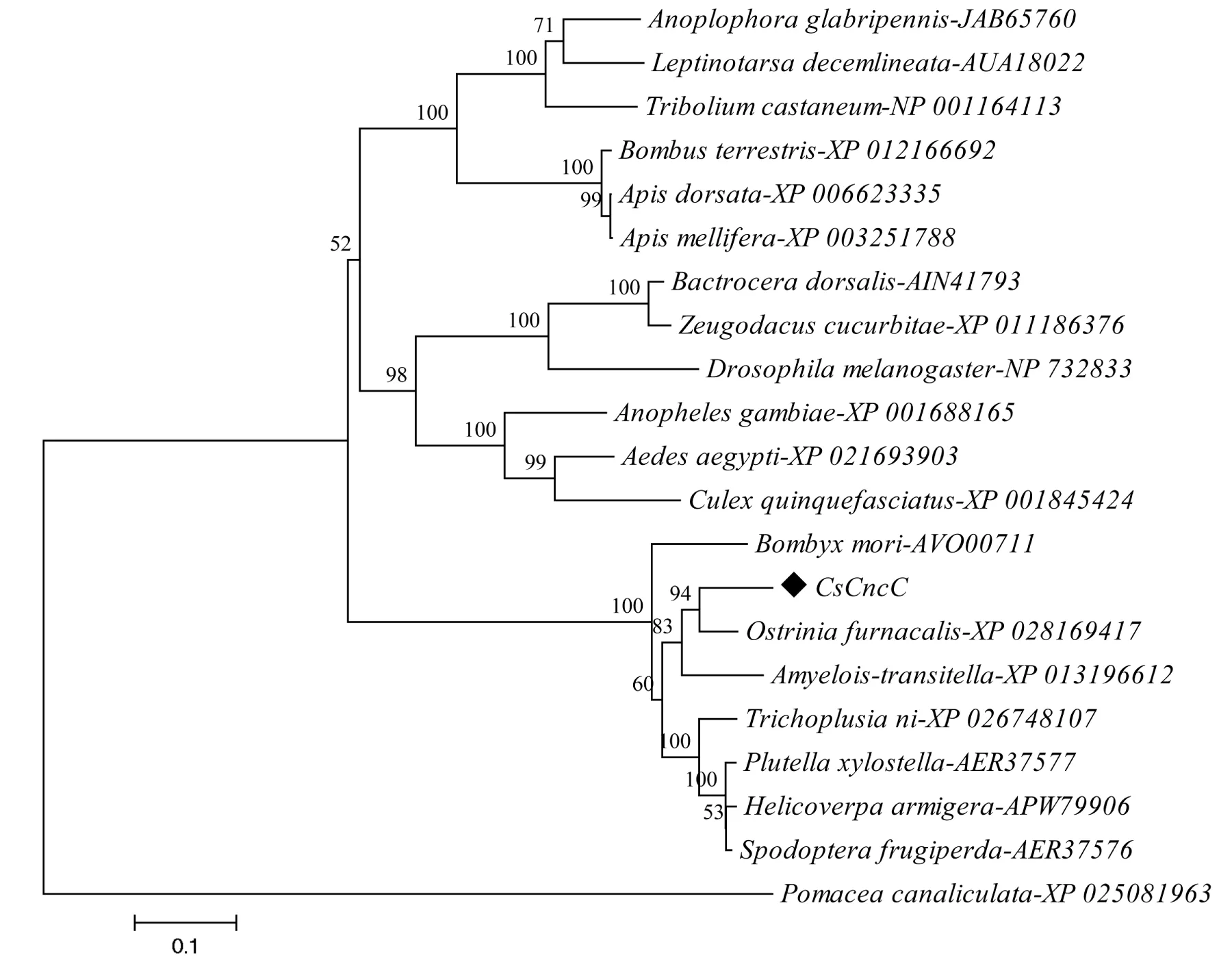
图3 二化螟CsCncC的系统发生分析Fig.3 Phylogenetic analysis of CsCncC
2.2 二化螟CsKeap1基因克隆及序列分析
通过克隆获得二化螟CsKeap1基因2 544 bp cDNA全长序列(GenBank登录号:MW147282),其中包含一个长度为1 923 bp的开放阅读框,一个321 bp的5′非翻译区为和一个300 bp的3′非翻译区(图4)。二化螟CsKeap1基因编码640个氨基酸,蛋白质分子质量为71.15 kDa,等电点为5.87。

图4 二化螟CsKeap1基因核苷酸序列及其编码的氨基酸序列Fig.4 Nucleotide and deduced amino acid sequences of CsKeap1 in Chilo suppressalis
氨基酸多重序列比对发现,二化螟CsKeap1蛋白具有昆虫Keap1家族的典型结构特征,如高度保守的BTB(bric-abrac/tramtrack/broad complex)区域、干预区(IVR,intervening region)/linker domain和双甘氨酸或Kelch重复区(DGR,double glycine or Kelch repeat)(图5)。对不同昆虫Keap1的系统发育树分析显示,二化螟CsKeap1与鳞翅目昆虫Keap1共聚一枝,具有较高的同源性(图6)。
2.3 CsCncC和CsKeap1的时空表达模式分析
2.3.1CsCncC和CsKeap1在二化螟不同发育阶段的表达模式分析
对CsCncC和CsKeap1在二化螟不同发育龄期(幼虫、蛹和成虫)中的表达模式分析发现,CsCncC在蛹末期的表达量最高,在其它龄期的表达水平变化不大(图7)。CsKeap1在二化螟预蛹期表达量最低,在成虫初期的表达水平显著高于其它龄期,羽化24 h后CsKeap1的表达水平显著降低并维持在较低水平(图8)。
2.3.2CsCncC和CsKeap1在二化螟不同组织中的表达模式分析
分别解剖二化螟神经索、脑、前肠、中肠、后肠、马氏管、血淋巴、脂肪体、表皮和卵巢组织,对CsCncC和CsKeap1在二化螟不同组织部位的表达量进行测定分析。结果显示,CsCncC在神经组织脑和神经索中的表达量较低,在中肠和马氏管中高表达,此外在前肠和卵巢中也具有较高的表达量(图9)。CsKeap1在二化螟脂肪体中的表达量最高,在中肠、马氏管、神经索和卵巢中也具有较高的表达水平(图10)。
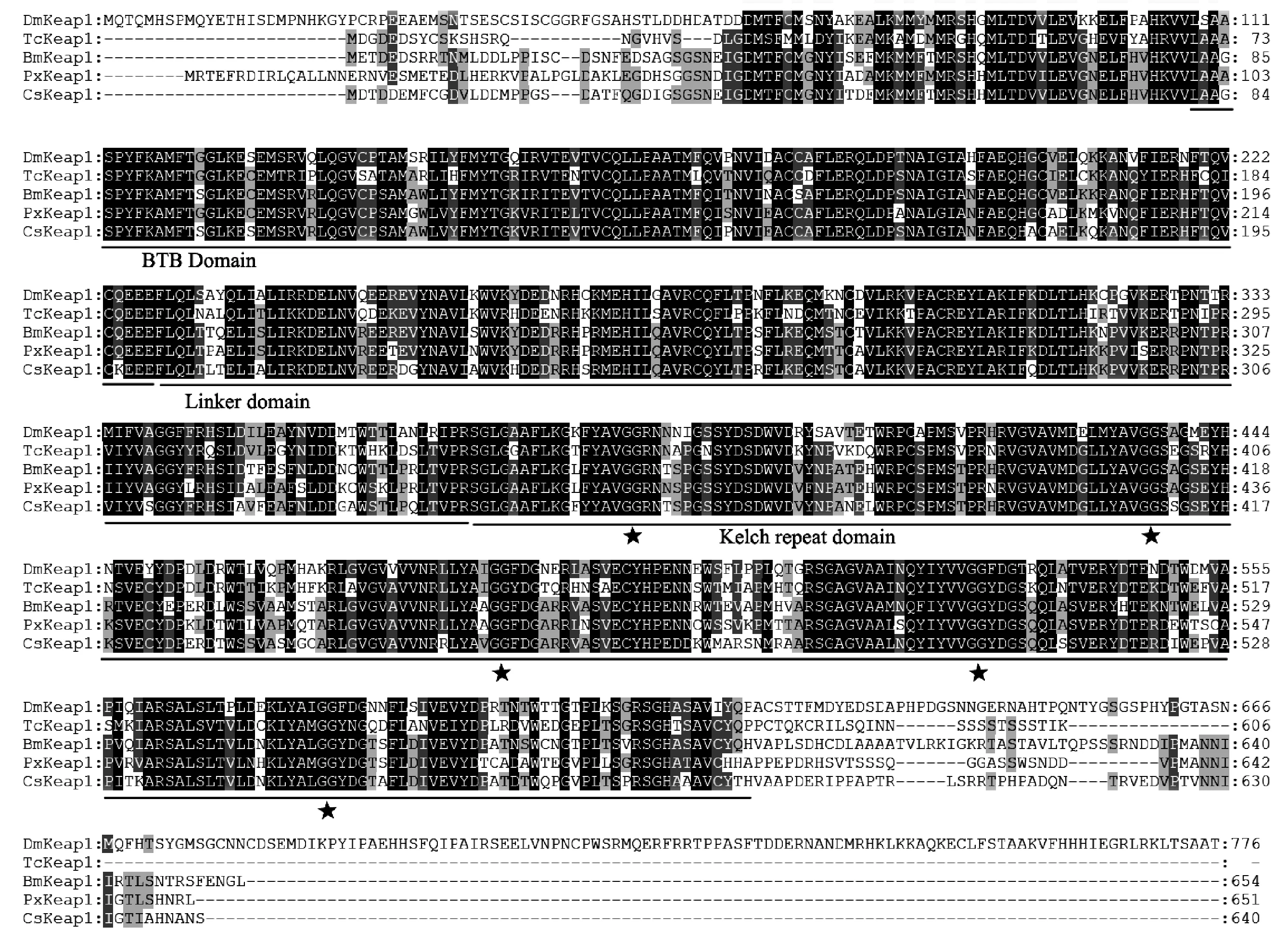
图5 二化螟与其它昆虫Keap1的氨基酸序列比对Fig.5 Multiple alignments of Keap1 from Chilo suppressalis and other insects
图6 二化螟Keap1的系统发育树分析Fig.6 Phylogenetic analysis of CsKeap1
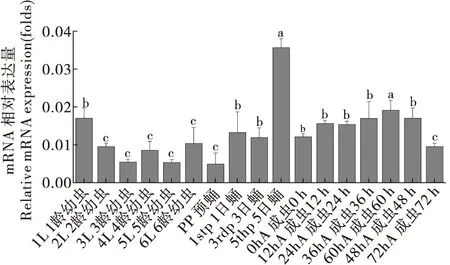
图7 CsCncC基因在二化螟不同发育龄期中的相对表达量Fig.7 Relative expression levels of CsCncC gene in different developmental stages of Chilo suppressalis
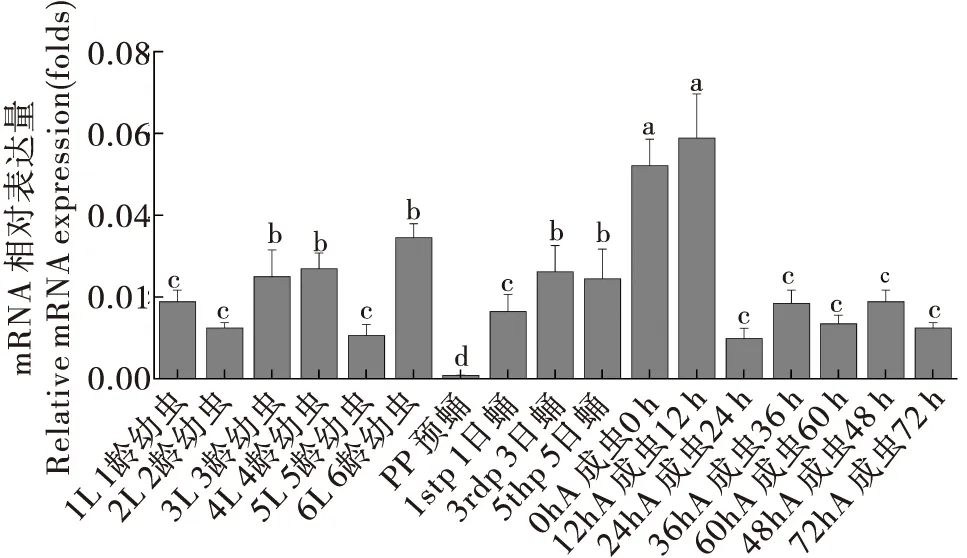
图8 CsKeap1基因在二化螟不同发育龄期中的相对表达量Fig.8 Relative expression levels of CsKeap1 gene in different developmental stages of Chilo suppressalis
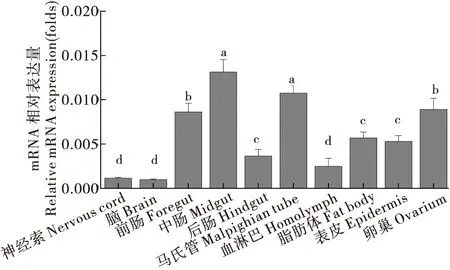
图9 CsCncC基因在二化螟不同组织中的相对表达量Fig.9 Relative expression levels of CsCncC gene in different tissues of Chilo suppressalis
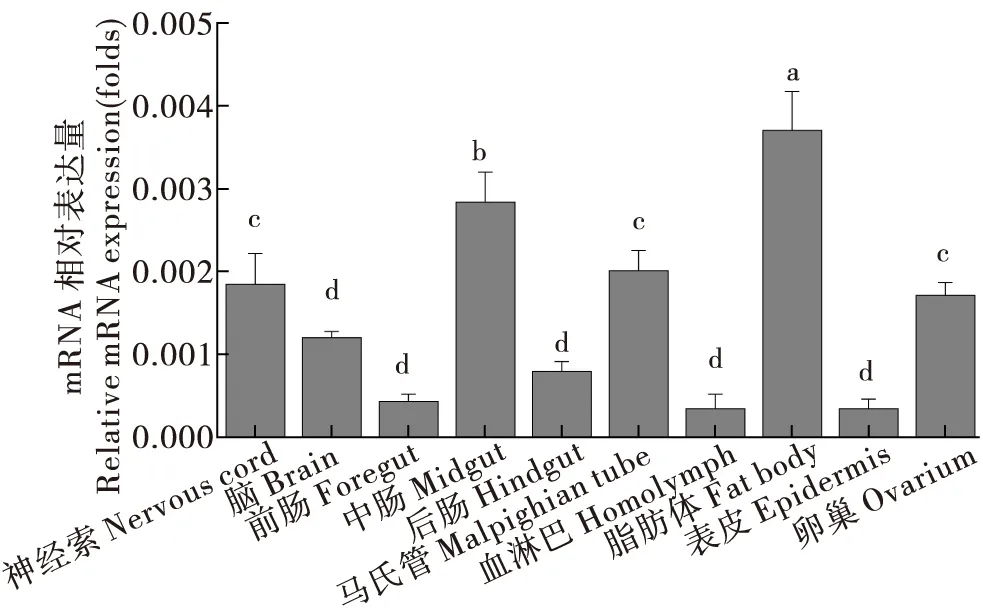
图10 CsKeap1基因在二化螟不同组织中的相对表达量Fig.10 Relative expression levels of CsKeap1 gene in different tissues of Chilo suppressalis
2.4 二化螟CsCncC和CsKeap1对氯虫苯甲酰胺的诱导响应
使用LC30和LC70剂量氯虫苯甲酰胺处理二化螟3龄幼虫,测定药剂处理对二化螟CsCncC和CsKeap1的表达量的影响。结果显示,使用LC30剂量氯虫苯甲酰胺分别处理幼虫12 h、24 h和36 h后,相比于对照,CsCncC在处理12 h和24 h后下调表达,在处理36 h后的表达量无显著变化(图11-A);CsKeap1的表达量均显著降低(图11-B)。使用LC70剂量氯虫苯甲酰胺处理后,CsCncC在处理12 h和24 h后的表达水平无显著变化,在处理36 h后被显著诱导表达(图11-A);CsKeap1的表达量在处理12 h后显著下降,在处理24 h后无明显变化,在处理36 h后则显著上调表达(图11-B)。

图11 二化螟CsCncC(A)和CsKeap1(B)对氯虫苯甲酰胺的诱导响应Fig.11 Induction responses of CsCncC(A) and CsKeap1(B) to chlorantraniliprole treatment in Chilo suppressalis
3 结论与讨论
Nrf2-Keap1通路调控细胞氧化还原平衡以及保护性细胞抗氧化和Ⅱ相解毒反应,对于维持机体内稳态具有至关重要的作用。本文对二化螟CsCncC和CsKeap1进行了克隆,结构域分析发现,与哺乳动物Nrf2相似,CsCncC具有负责与小Maf蛋白(small Maf proteins)形成异源二聚体并与靶标基因的抗氧化反应元件(ARE,antioxidant response element)结合的bZIP结构(Nioietal.,2005)以及对结合keap1起关键作用的保守性ETGE基序(Chowdhryetal.,2013),但与其它鳞翅目昆虫一致,与Nrf2和Keap1识别结合相关的DLG基序变为DMG(Huetal.,2018)。Cskeap1蛋白分别含有BTB、linker和Kelch结构域。BTB结构域在介导Keap1形成同源二聚体和招募Cullin-3 (Cul3)过程中具有重要作用(Tkachevetal.,2011;Pitoniak and Bohmann, 2015)。IVR结构域富含半胱氨酸,对氧化应激信号敏感(Kimetal.,2014;Yangetal.,2017)。在正常条件下,Keap1会与Nrf2相互结合形成异二聚体,多余的Nrf2会被迅速降解掉,从而维持胞质中Nrf2的低水平。当机体受到氧化刺激时,Keap1的IVR区域会被迅速激活,这个过程会抑制Nrf2的降解,从而诱导Nrf2在胞质中的积累(Lietal.,2008)。DGR区也叫Kelch区,其中含有5个双甘氨酸重复序列,是Keap1与Nrf2的结合区。系统发生分析也表明,CsCncC和CsKeap1与其它鳞翅目昆虫同源蛋白聚为一支。
基因表达模式通常与其生理功能相关。对家蚕BombyxmoriLinnaeus的研究发现,BmCncC在5龄幼虫表皮、马氏管、头部和脂肪体表达水平较高,Bmkeap1在5龄幼虫脂肪体、头部、生殖腺和表皮表达水平较高(Wangetal.,2020)。对马铃薯甲虫的研究发现,LdCncC在幼虫中肠、前胸腺和雌成虫卵巢中高水平表达,LdKeap1在脑-心侧体-咽侧体复合体中表达水平最高,在幼虫前肠、前胸腺和雌成虫卵巢中表达水平也较高(Sunetal.,2017)。本文研究发现,CsCncC在二化螟中肠和马氏管中的表达水平最高,在前肠和卵巢中也具有较高的表达水平;CsKeap1在二化螟脂肪体中的表达水平最高,在中肠、马氏管、神经索和卵巢中也具有较高的表达水平。消化道是昆虫摄入外源化合物后的第一道防线,马氏管和脂肪体是昆虫对外源化合物进行降解和解毒代谢的重要组织,因此CsCncC和CsKeap1在二化螟不同组织中的表达模式表明这2个基因在二化螟的解毒代谢过程中具有重要作用。值得注意的是,与家蚕相似,CsCncC和CsKeap1在卵巢中也具有一定的表达水平,表明昆虫CncC和Keap1除了在抗化学和氧化胁迫中具有重要作用外,还可能具有其它生理功能。
氯虫苯甲酰胺是美国杜邦公司开发的高效、安全的二酰胺类杀虫剂。研究发现,二化螟对氯虫苯甲酰胺的抗性与细胞色素P450基因CYP6CV5,CYP9A68,CYP321F3和CYP324A12以及UDP-葡萄糖醛酸基转移酶CsUGT40AL1和CsUGT33AG3的过表达相关(Xuetal.,2019;Zhaoetal.,2019)。本文使用LC30和LC70浓度氯虫苯甲酰胺处理二化螟3龄幼虫,发现LC30浓度处理并不能诱导CsCncC和CsKeap1的表达,而用LC70浓度处理36 h后这两个基因表达水平都显著上调。对家蚕的研究也发现,0.01 mg/L氯虫苯甲酰胺处理家蚕5龄幼虫后,脂肪体中BmCncC和Bmkeap1的mRNA表达水平均显著上调(Maoetal.,2019),但4 mg/L辛硫磷处理家蚕5龄幼虫后24 h后,BmCncC的表达显著上调,而Bmkeap1的表达水平显著下调(Huetal.,2018),表明农药对昆虫CncC和Keap1的诱导效应可能具有剂量依赖性,并且这种诱导效应也因杀虫剂而异。今后有必要进一步研究氯虫苯甲酰胺处理对CsCncC亚细胞定位的影响以及CncC-keap1通路在二化螟对氯虫苯甲酰胺抗性中的作用。
