襀翅目化石稚虫口器及其食性分析
2022-01-05刘玉双张永娇
刘玉双,张永娇
(1. 沈阳师范大学古生物学院,沈阳 110034;2. 辽宁省文物考古研究院,沈阳 110003)
襀翅目昆虫,俗称石蝇、襀翅虫,是昆虫纲的一类水生昆虫。成虫陆生,多生活于水边的石头、枯枝、草丛、灌木或较高大树木的树枝上,多数不取食,少数种类危害农作物及果树;卵和稚虫水生,形态多样,食性因种类不同而具有很大差异。此外,襀翅目昆虫生境适应范围狭窄,不同种类对水质条件及周边植被状况要求不同,具有其独特的小生境,特别是稚虫对水质污染反映敏感,常用于水质监测,是国际流行的EPT(Ephemeroptera-蜉蝣目、Plecoptera-襀翅目、Trichoptera-毛翅目)水质生物监测的三大水生昆虫之一(Du, 1999)。
化石记录表明襀翅目昆虫在地球上至少存活了约3.2亿年之久,在漫长的地质历史中,留下了珍贵的化石资源(Béthoux, 2011)。特别是近些年,辽西早白垩世义县组地层中发现许多近原地埋藏的襀翅目稚虫化石,形态多样,保存完好,为古生态学研究提供了很好的材料。本文着重襀翅目稚虫化石形态、口器结构,及其食性的分析,结合现生襀翅目稚虫形态、生物学等特征,以及前人对襀翅目稚虫化石研究的成果,对辽西白垩世义县组地层中的襀翅目稚虫化石食性做了综合分析,为该地区的古生态学研究提供有价值的信息。
1 襀翅目现生稚虫的食性
现生襀翅目昆虫(特别是稚虫)食性多样,是水生生态系统中一个重要成员,在水生生态系统平衡和水体净化上具有一定的作用。植食性稚虫取食水中植物碎片、藻类和苔藓等,扮演着水中清道夫的角色;同时也被其它肉食性水生昆虫所取食,或者成为鱼类、小龙虾等水生动物的食料,是渔业养殖的重要食料来源。肉食性稚虫则捕食蜉蝣稚虫、蚊蝇幼虫、鱼卵和其它水生小型动物,对渔业养殖同样具有一定的影响。腐食-植食性(Detritivorous-herbivorous)、肉食性(Carnivorous)以及食性复杂难以确定或称之为杂食性(Omnivores),是目前襀翅目现生稚虫食性的大致情况。此外,有些种类的稚虫还会因发育阶段不同而食性不同,低龄稚虫不论肉食性、腐食-植食性的种类均以细小的碎屑物为食(Du, 1999; Tierno de Figueroa and López-Rodríguez, 2019)。
石蝇稚虫的食性与它们的口器外部形态特征密切相关,虽然同为咀嚼式口器,但食性不同,口器构造差异明显,尤其是上颚和下唇的结构。通常来说,肉食性种类上颚顶端具多个尖齿,磨区不明显,中唇舌退化,侧唇舌发达,如北襀亚目原颚组Systellognatha的大多数类群(图1-A, C)。腐食-植食性种类的口器上颚顶端具多个钝齿,磨区发达,中唇舌和侧唇舌近等长,指状,如北襀亚目Arctoperlaria真颚组Euholognatha和南襀亚目Antarctoperlaria的大多数种类(图1-D, F)(Du, 1999; Olifiers, 2004)。
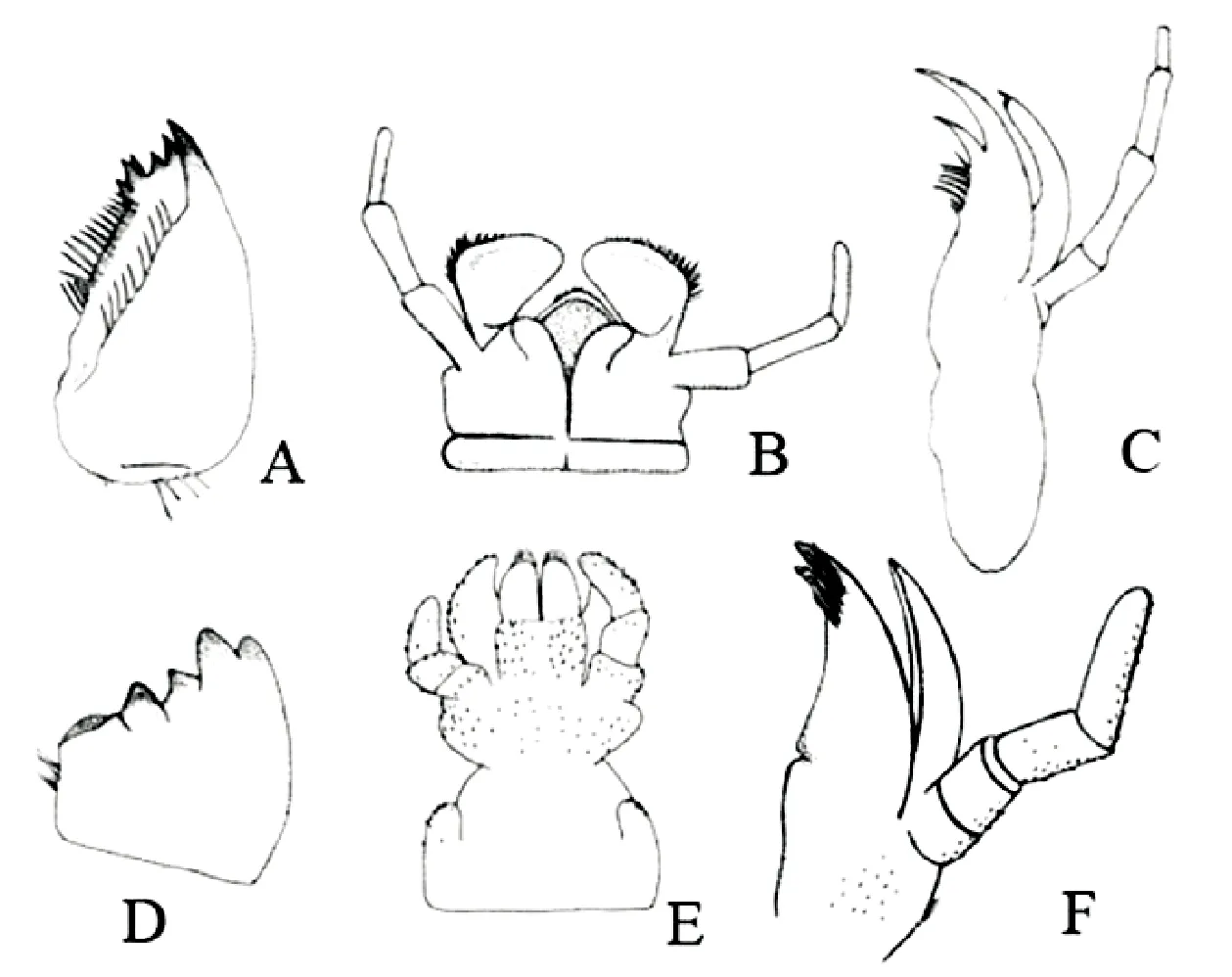
图1 现生襀翅目昆虫的口器结构Fig.1 Mouthparts aspect of the living Plecoptera注:A,右上颚(Kempnyia sp.);B,下唇(Kempnyia sp.);C,右下颚(Kempnyia sp.);D,右上颚(Gripopteryx sp.);E,下唇(Gripopteryx sp.);F,右下颚(Guaranyperla sp.)(引自Olifiers et al., 2004)。Note: A, Right mandible (Kempnyia sp.); B, Labium (Kempnyia sp.); C, Right maxilla (Kempnyia sp.); D, Right mandible (Gripopteryx sp.); E, Labium (Gripopteryx sp.); F, Right maxilla (Guaranyperla sp.) (Olifiers et al., 2004).
2 襀翅目化石稚虫的食性
现生襀翅目昆虫通过饲养、观察以及肠道内存物分析可知其食性,但这些方法在化石种类的食性研究上不实用,由于化石保存的局限性,肠道内存物很少保存,无法准确分析其食性。目前,仅Sinitshenkova记载过蒙古地区早白垩世地层中发现带襀科的一种石蝇成虫Gurvanopteryxeffeta化石,其肠道内存物清晰可见,但不定形,推测其以液体食物为食(Sinitshenkova, 1986)。
昆虫口器的形态结构影响其取食模式及其消化的食物类型,能很好地反映其取食机制。作为昆虫的取食器官,口器在长期的进化过程中因食性和取食方式的不同,而形成多种多样的口器类型,并且口器结构在不同类群中往往是独特的,口器的形态学研究可为昆虫的食性分析提供依据(Krenn, 2007; 金丽, 2012)。目前,作为高度整合的取食功能单元,国内外学者已经开展了不同类群的口器形态及其功能的广泛研究(Geetal., 2004; Smith and Capinera, 2005)。
襀翅目稚虫的食性与其口器特征密切相关,并且口器部分结构骨化程度高,可以保存形成化石,因此,石蝇化石稚虫的食性可以根据口器结构及其分类位置间接分析获得。通过观察石蝇化石中保留的口器结构特征,及参照亲缘关系密切的现生石蝇稚虫的口器形态及食性,分析地史时期石蝇稚虫食性的研究方法是比较可行的。俄罗斯古生物学家Sinitshenkova曾根据这一方法分析襀翅目部分化石种类的食性,将襀翅目化石种类的食性分为腐食-植食性、刮食性、肉食性以及食性特征不确定4类,腐食-植食性又细分为粗糙型和细小型2种;她认为襀翅目Perlomorpha的大部分化石类群为肉食性,Gripopterygomorpha和Nemourina的大多数化石种类则是腐食-植食性,低龄稚虫无论是腐食-植食性还是捕食性的均取食于植物细小的碎屑(Sinitshenkova, 1987; 1997)。
2.1 腐食-植食性
腐食-植食性是石蝇稚虫化石的主要食性,取食死亡和不同程度腐烂的植物组织,较少吃新鲜植物,故称之为腐食-植食性,这种食性的稚虫化石通常保存了坚硬的上颚,且轮廓清晰,甚至有些稚虫的身体结构保存模糊,但上颚、上唇和下唇须清晰可见,这些结构均是腐食-植食性口器的典型特征。Sinitshenkova将这种石蝇口器比作碎屑机,体大型的稚虫,其口器似大型碎屑机,撕碎全叶或大片植物叶片,如灭绝的Siberioperlidae的一些种类,Siberioperlalacunose的上颚结构与现生食较硬的、粗糙植物碎片的Austroperlidae和Pteronarcyidae相似,但其上颚具更发达的研磨构造,上颚顶端内弯且坚硬,具短而钝的牙齿(图2-A),推测其可能取食一些特别坚硬的植物,如轮藻的钙质组织;体小型的稚虫,其口器似小型碎屑机,磨食小的薄的叶片,如灭绝的Palaeonemouridae、Baleyopterygidae、Mesoleuctridae和Perlariopseidae的一些种类,Mesoleuctragracilis上颚结构较传统(图2-B),推测其可能取食蕨类植物或裸子植物叶片。
2.2 刮食性
这一食性实际上可归属于上一种食性,刮食固着于植物或石头上的藻类等,如现生Taeniopterygidae的Brachyptera。Sinitshenkova将这一食性单独提出,并推测德国上新世地层中发现的B.schmidti的食性为这一类型。此外,灭绝种类Spinoperlaspinose的食性也可能是这种食性。

图2 不同襀翅目化石昆虫的部分口器结构Fig.2 Part mouthparts aspect of the different fossil Plecoptera注:A,上颚(Siberioperla lacunose),比例尺=1 mm;B,上颚(Mesoleuctra gracilis),比例尺=1 mm;C,上颚(Derancheperla collaris),比例尺=0.5 mm;D,下颚(Trianguliperla quassa);比例尺=1 mm(引自Sinitshenkova, 1982; 1983, 1990)。Note: A, Mandible (Siberioperla lacunose), scales=1 mm; B, Mandible (Mesoleuctra gracilis), scales=1 mm; C, Mandible (Derancheperla collaris), scales=0.5 mm; D, Maxilla (Trianguliperla quassa), scales=1 mm (Sinitshenkova, 1982; 1983; 1990).
2.3 肉食性
肉食性石蝇稚虫的口器与前两种食性的稚虫口器不同,其上颚较狭长,骨化程度低,端部具锋利的齿,下颚具锋利的长齿,及细长的下颚须。目前发现的捕食性石蝇稚虫化石多集中在灭绝科Platyperlidae、Tshekardoperlidae和灭绝属Trianguliperla,以及现生科Perlodidae的灭绝属Derancheperla。Sinitshenkova根据化石保存的口器特征,推测灭绝属Platyperla、Trianguliperla和Derancheperla的稚虫食性为肉食性,上颚狭长,顶端具锋利的尖齿(图2-C);下颚也保存有锋利的长齿(图2-D)。此外,Platyperla稚虫上颚保存多不完整,轮廓不清晰,有压痕,表明其骨化程度弱;并且在稚虫化石P.platypoda肠道内容物中未发现植物表皮碎片,其内容物为暗色致密物质和非结构化的碎片,极有可能是与Platyperla稚虫一起生活的蜉蝣稚虫,推测Platyperla主要食物是弱小的蜉蝣稚虫和其它水生稚虫。Trianguliperla稚虫则有可能在水中捕食双翅目水生昆虫的幼虫。此外,Sroka等描述了缅甸北部克钦邦琥珀中Perlidae的一个稚虫种类Electroneuriaronwoodi,其生存时代为白垩纪中期(Srokaetal., 2018),这一琥珀石蝇稚虫保存了明显的肉食性口器特征:右上颚端部保留了4个圆形的齿,左下颚的外颚叶狭窄,微短于内颚叶,内鄂叶具2个长的尖齿。
2.4 未知食性
由于保存条件的限制,许多石蝇稚虫化石存在保存不完整、信息残缺等问题。那些未保存口器的石蝇稚虫化石的食性是难以推测的,故食性未知。此外,有些保存口器的稚虫化石,其口器特征处于中间过渡状态,不明显接近于腐食-植食性口器或肉食性口器,如灭绝科Palaeoperlidae和Perlopseidae中的一些化石稚虫,这类化石种类的食性同样难以确定。
3 早白垩世义县组襀翅目化石稚虫的口器及其食性分析
近年来,辽西北票早白垩世义县组地层中发现许多保存较好的石蝇稚虫化石和及少量的成虫化石。成虫化石,口器退化,很难保存,其它外部形态特征鉴定隶属于现生科级阶元Perlidae、Taeniopterygida和Nemouridae(Liuetal., 2007; Liuetal., 2008);稚虫化石则分类特征保存较少,初步鉴定隶属于灭绝科Mesoleuctridae和Platyperlidae,以及现生科Perlidae、Taeniopterygidae和Nemouridae,其中Perlidae稚虫化石发现众多,其身体粗壮,硬体部分骨化程度相对发达,少数标本保存下来部分口器结构。
本文研究的化石材料为Perlidae一稚虫样本(图3-A),正负版保存,PMOL-AI00910AB,体长(不包括触角和尾须)约23 mm,其头部、前胸和腹部具Perlidae典型的斑纹特征,以及单眼2个。此外,此标本清晰地保存了口器的上颚印痕,可清楚的看到上颚端部具有4个明显尖锐的小齿(图3-B, E),用来撕裂食物,磨区不明显,结合其系统分类位置,现生Perlidae稚虫通常都是捕食者,上颚具有类似结构;故而推测这个化石稚虫的食性为肉食性。

图3 辽西早白垩世义县组Perlidae化石稚虫Fig.3 Fossil Perlidae nymph from Yixian Formation in the west of Liaoning注:A,稚虫,PMOL-AI00910A,比例尺=4 mm;B,头部线条图,PMOL-AI00910B,比例尺=2 mm;C,左上颚,PMOL-AI00910A,比例尺=1 mm;D,右上颚,PMOL-AI00910B,比例尺=1 mm;E,头部,PMOL-AI00910B,比例尺=2 mm。Note: A, Nymph, PMOL-AI00910A, scales=4 mm; B, Line drawing of head, PMOL-AI00910B, scales=2 mm; C, Left mandible, PMOL-AI00910A, scales=1 mm; D, Right mandible, PMOL-AI00910B, scales=1 mm; E, Head, PMOL-AI00910B, scales=2 mm.
4 结语
通过昆虫口器形态特征可以分析食性,其食性特征又是区分物种的重要特征,王书永等(2005)提出在外部形态特征差异不明显的情况下,食性特征往往是区分物种的关键特征或区分标志。襀翅目稚虫体柔软,且由于化石保存的局限性,一些鉴别特征不能完全清晰的保存下来,骨化程度较高的口器结构(如上颚)如若能保存下来,其形态特征,食性分析则有利于石蝇稚虫化石的鉴定。
此外,环境可促进口器形态的演变,口器特征也反映一定的环境信息(刘明等, 2005);襀翅目昆虫生境适应范围狭窄,作为水生环境指示昆虫之一,其在水生生态系统中占有一席之地,口器特征及食性分析也可为推测其生存的水生生态环境提供一些信息。当然,作为古生物研究的热点,无论是类群古生态分析还是区域古生态环境构建,我们还需要综合利用多方面证据,如系统发育、伴生生物、沉积矿物、岩相、孢粉分析,及地化分析等信息,利用将今论古、形态比较、功能分析、以及埋藏学分析等方法,交叉综合分析、印证,才可能较真实地重建当时的古生态环境。
