A New Species of Glyphoglossus Günther,1869 (Anura:Microhylidae) from Western Yunnan,China
2021-12-30DongruZHANGShuoLIULixiaZHANGHongHUIHengXIAOandDingqiRAO
Dongru ZHANG ,Shuo LIU ,Lixia ZHANG ,Hong HUI ,Heng XIAO and Dingqi RAO*
1 State Key Laboratory of Genetic Resources and Evolution,Kunming Institute of Zoology,Chinese Academy of Sciences,Kunming 650223,Yunan,China
2 College of Life Sciences,Yunnan University,Kunming 650091,Yunan,China
3 Kunming Natural History Museum of Zoology,Kunming Institute of Zoology,Chinese Academy of Sciences,Kunming 650223,Yunan,China
4 Department of Ecology,College of Life Sciences,Henan Normal University,Xinxiang 453007,Henan,China
5 Research Center for ecological management and protection of the Yellow River Basin,Xinxiang 453007,Henan,China
Abstract A new species of microhylid frog of the genus Glyphoglossus Günther,1869 is described from Huadianba,Cangshan Mountain,Dali Bai Autonomous Prefecture,Yunnan Province,China.Glyphoglossus huadianensis,new species,is compared with congeners from China and other parts of Southeast Asia,and was distinguished from the others by the following combination of characters:adult male body size up to 37.8 mm;pupil rounded;supratympanic fold distinct;tympanum concealed;toe tips obtuse;subarticular tubercles prominent and rounded;foot webbing extensive;outer metatarsal tubercle present;dorsum tuberculate,yellowish-brown/earth-yellow;and a pair of conspicuous large round spots in groin.The new species is the tenth species of Glyphoglossus to be described,and the second known from China.
Keywords Glyphoglossus huadianensis sp.n.,taxonomy,morphology,China
1.Introduction
Currently,nine nominal species,which are known from Southeast Asia (Frost,2020),are recognized in the microhylid genusGlyphoglossus
Günther,1869.Only one species is known in China,Glyphoglossus yunnanensis
(Boulenger,1919).Owing to rare collection,little is known for most species exceptGlyphoglossus molossus
Günther,1869,Glyphoglossus guttulatus
(Blyth,1856) andGlyphoglossus yunnanensis
.The systematics ofGlyphoglossus
were once very confusing for above reason,and several junior synonyms are on record:Calluella
Stoliczka 1872,Colpoglossus
Boulenger1905,
Dyscophina
van Kampen 1905,andCalliglutus
Barbour and Noble 1916 (Frost,2020).The genusGlyphoglossus
was designated to placeGlyphoglossus molossus
,which was originally unassigned to any subfamily of Microhylidae (Frostet al
.,2006),and was subsequently transferred to Microhylinae by Van Bocxlaeret al
.(2006).Matsuiet al
.(2011) reportedCalluella
Stoliczka,1872,to be paraphyletic with respect toGlyphoglossus
based on mtDNA phylogenetic analysis.Meanwhile,a sister-relationship ofGlyphoglossus
withCalluella
was also suggested by Pyron and Wiens (2011).Based on denser samplings ofCalluella
,Daset al
.(2014) confirmed the placement ofGlyphoglossus
withinCalluella
.According to the law of priority,Calluella
Stoliczka,1872 was recognized as a junior synonym ofGlyphoglossus
by Pelosoet al
.(2016).Based on extensive sampling and in-depth investigation into the phylogeographic structure ofG.yunnanensis
,Zhanget al
.(2020) discovered there were four main maternal lineages in this nominal species on base of 2154 bp concatenated mitochondrial gene fragments,one of which (lineage LD,Figure 1) occurs in western Yunnan,China and is deeply separated from the remaining lineages with an average divergence of 9.3% (LD/E,9.6%;LD/W,9.1%;and LD/C,9.2%.) (Figure 1).Lineage LD was also shown to be differentiated from other known congeners(Figure 1).Here we further investigate the identity of lineage LD and describe it as a new species.Morphological comparisons demonstrate that the new species is distinctive fromG.yunnanensis
and other known congeners and therefore warrants taxonomic recognition.2.Materials and Methods
2.1.Sampling
Seven specimens were collected in Dali Bai Autonomous Prefecture,Yunnan,China (25°52’05’ N,100°01’05’E) in May,2010,and six in Lijiang,Yunnan,China (26°51’18’N,100°13’40’ E) in June,2010.Following Animal Use Protocols approved by the Kunming Institute of Zoology Animal Care and Ethics Committee,specimens were photographed in the field prior to euthanasia,and then fixed in 90% ethanol before being stored in 70% ethanol.A portion of liver or muscle tissues were preserved in 99% ethanol.Sexes of the type specimens were determined through checking vocal sac openings.Specimens and tissue samples were deposited at Kunming Institute of Zoology,Chinese Academy of Sciences(KIZA00252–KIZA00258,KIZ 2014005754,KIZ 2014005781–KIZ 2014005785).2.2.Genetic distance
Pairwise genetic distances (Kimura 2-parameter) among species ofGlyphoglossus
were calculated using MEGA 7 (Kumaret al
.,2016) based on a concatenated data of 16S rRNA,CO1,and CYTB of Zhanget al
.(2020).The GenBank accession numbers of specimen used in this study were compiled in Table S1.2.3.Morphology
The morphological measurements were taken to the nearest 0.1 mm with digital calipers.Comparative data on the morphology and taxonomy ofGlyphoglossus
were obtained from previous publications:Barbour and Noble (1916),Blyth (1856),Boulenger (1904;1919),Daset al
.(2004),Daset al
.(2014),Feiet al
.(1999),Günther (1869),Inger (1966),Kiew (1984),Liu (1950),Manthey and Grossmann (1997),Parker (1934),and Van Kampen (1905).Morphylogical terminology followed Daset al.
(2004) and Daset al
.(2014).Abbreviations used for measurements (in mm)and meristic characters are as follows:Snout-vent length (SVL,from tip of snout to vent);tibia length (TBL,distance between surface of knee and surface of heel,with both tibia and tarsus flexed);head length (HL,distance from tip of snout to angle of jaws);head width (HW,measured at angle of jaws);head depth(HD,greatest transverse depth of head,taken posterior of the orbital region);eye diameter (ED,horizontal diameter of the eyes);interorbital distance (IO,least distance between upper eyelids);internarial distance (IN,distance between nostrils);eye to snout distance (E-S,distance between anterior-most point of eyes and tip of snout);eye to nostril distance (E-N,distance between anterior-most point of eyes and nostrils);eye to tympanum distance (E-T,distance between posterior-most point of eye and anterior edge of tympanum);axilla to groin distance (A-G,distance between posterior edge of forelimb at its insertion to body to anterior edge of hindlimb at its insertion to body);body width (BW,greatest width of body);tympanum diameter (TD,vertical and horizontal);width of upper eyelid(UE,greatest width of upper eyelid).Webbing formula notation follows Savage and Heyer (1967) as modified by Myers and Duellman (1982).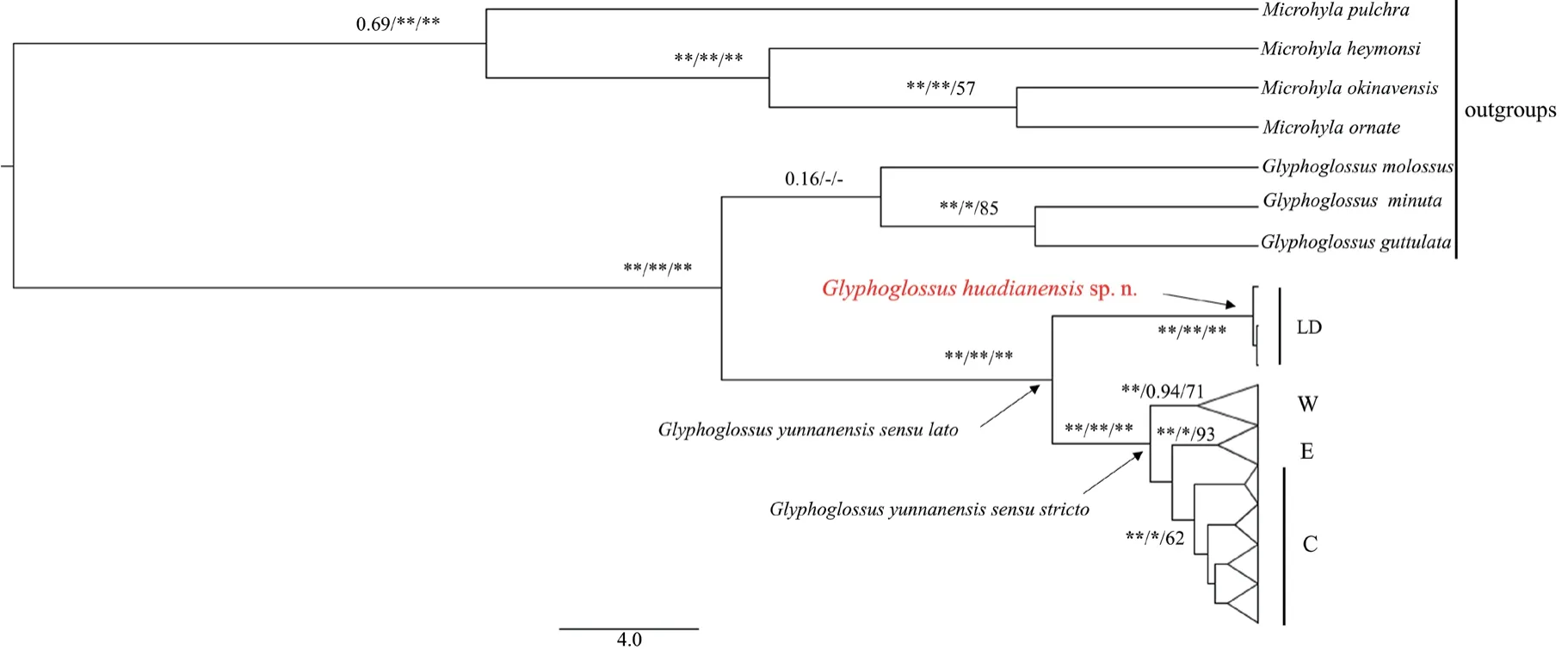
Figure 1 Simplified Bayesian inference (BI) tree of the species G.yunnanensis sensu lato reproduced from Zhang et al. (2020).Posterior probabilities (BEAST),Bayesian Posterior Probabilities (BPP),and maximum parsimony (MP) bootstrap values (* >95%,** >99%) are shown for main clades.Vertical bars show clade assignment.
With a correlation matrix of size-standardized measurements (all measurements divided by SVL),a multivariate principal component analysis (PCA) was performed.Scatter plots of the first two principal components scores were used to investigate the differences between the new species andG.yunnanensis
.The significance of differences between the two species for sizecorrected values of measurements was tested by means of one-way ANOVA (P
<0.01 as significant).Data were firstly tested for normality (using Shapiro-Wilk Test) and homogeneity (using Levene’s test).All analysis above mentioned were conducted in IBM SPSS Statistics 24 (IBM Corp).3.Results
3.1.Genetic divergence
Lineage LD was deeply separated fromG.yunnanensis sensu stricto
(lineage W+C+E) with a divergence of 8.9%,and from other known congeners 7.8%–20.0% (Table 1).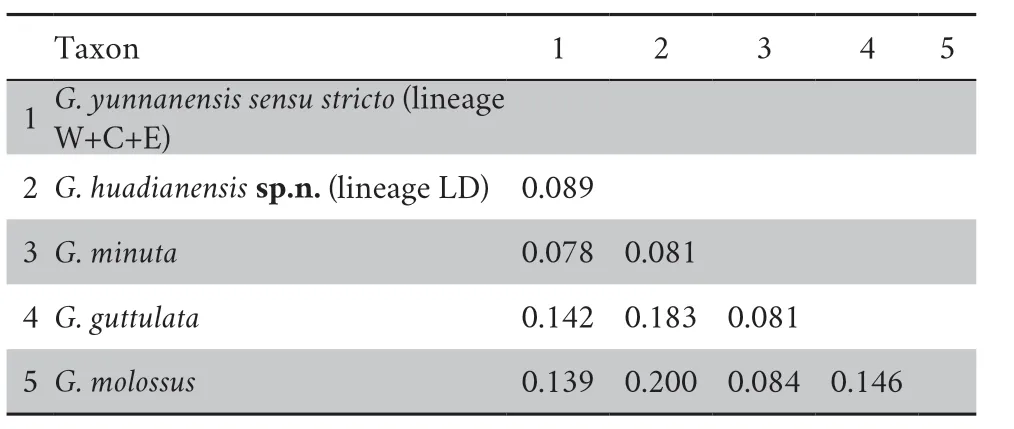
Table 1 Estimates of Net Evolutionary Divergence between Glyphoglossus huadianensis sp.n.and other known congeners.
3.2.Morphometric analysis
Morphometric data of the new species andG.yunnanensis
are summarized in Table 2.The first two principal components were retained,which had eigenvalues above 1.0 and cumulative contribution of variance accounted to 58.14% (Table 3).Loadings for PC1,which accounted for 46.95% of the total variance,were most heavily loaded (load factor >0.70) on HL,HD,TD,E-S,E-N,IO,and HW (Table 3).Differentiation was found along the PC 1 axis betweenG.yunnanensis
and the new species (Figure 2).The second principal component (PC 2) accounted for 11.19% of the total variance (Table 3),but no clear separation was observedalong this axis between the new species andG.yunnanensis
(Figure 2).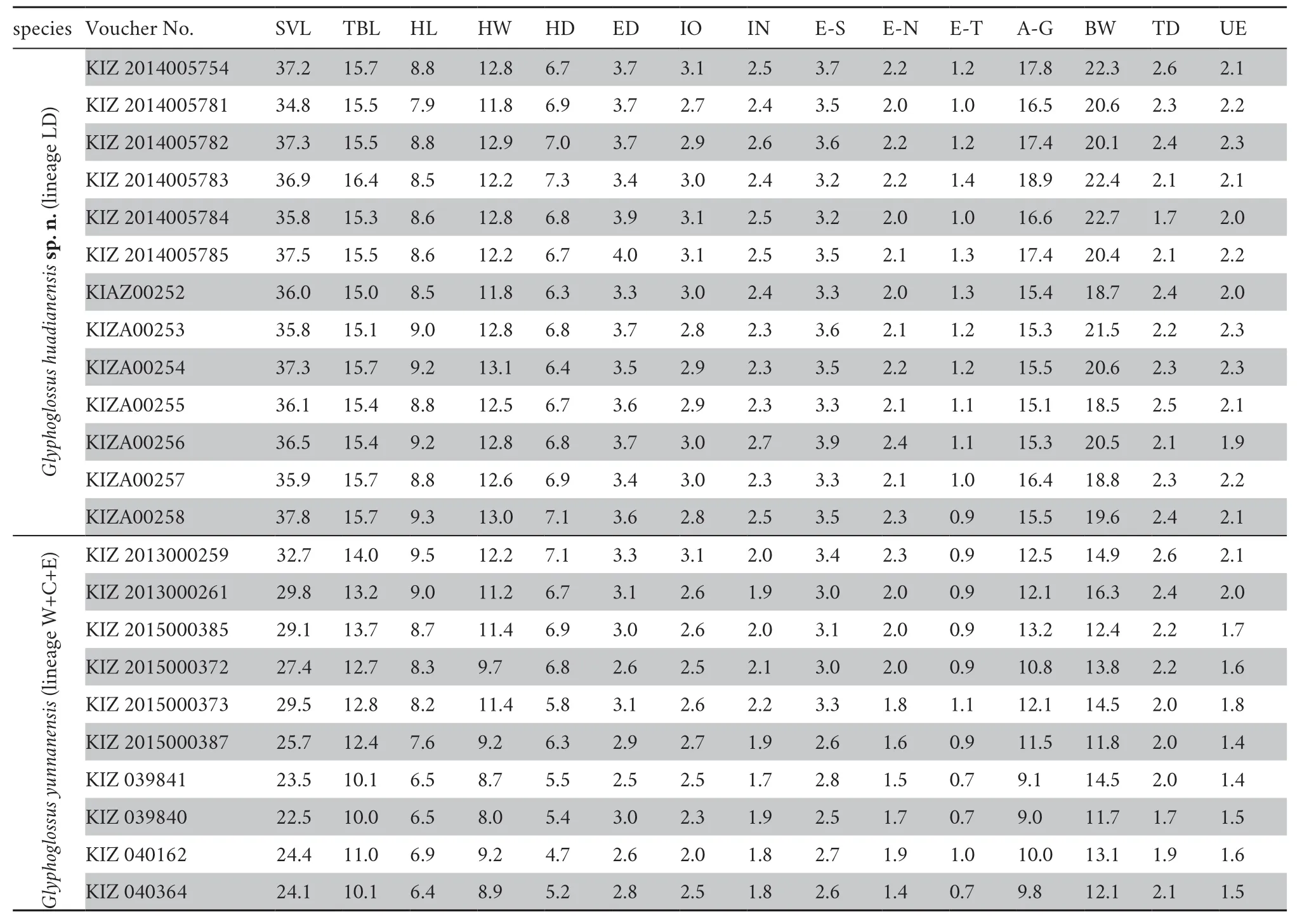
Table 2 Measurements (in mm) of G.huadianensis sp.n. and G.yunnanensis. The specimens are all males.Abbreviations defined in text.
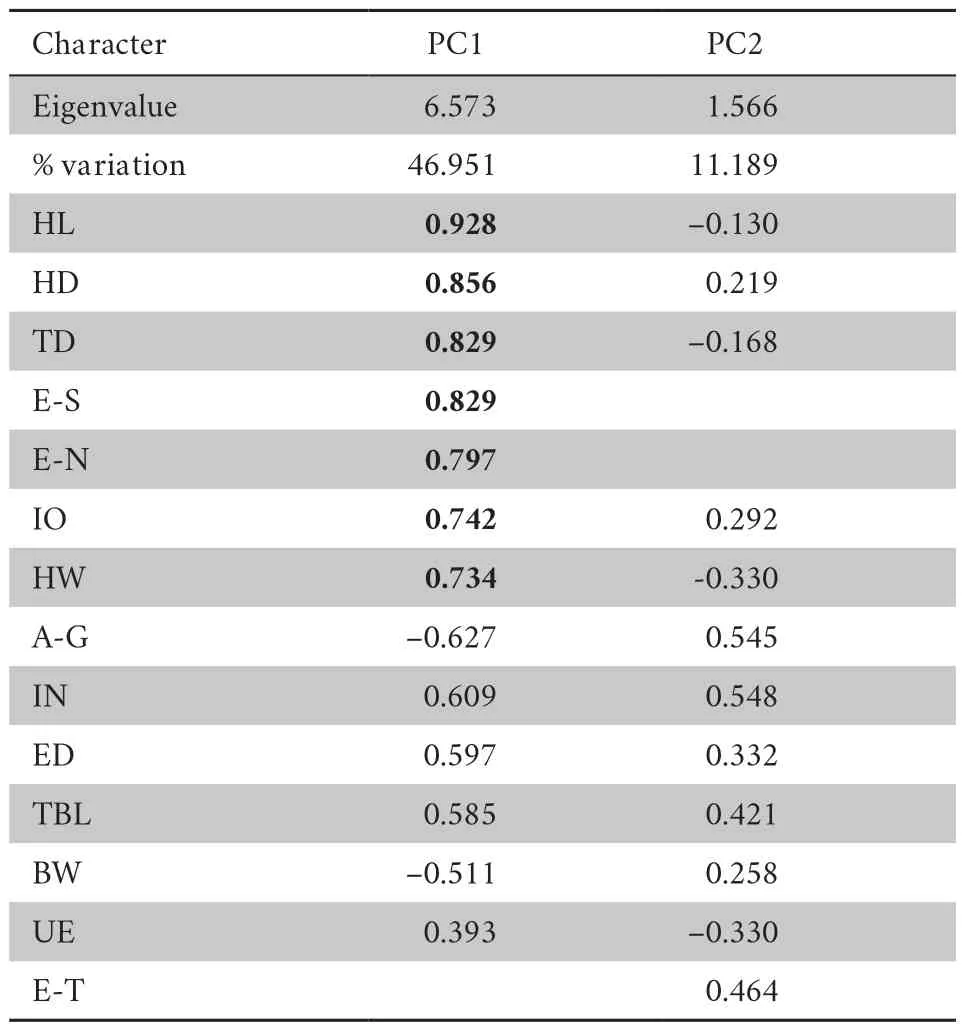
Table 3 Factor loadings of the first two principal components of 14 sizeadjusted morphometric characteristics of males of Glyphoglossus huadianensis sp.n. and G.yunnanensis.Absolute values of loading greater than 0.70 in boldface.Abbreviations defined in text.
The sizecorrected values of measurements follow the normal distribution (Table S2).The results of homogeneity of variables showed thatP
-value of HD,IO,and E-N were less than 0.01,rejecting null hypothesis (Table S3).One-way ANOVA test show significant difference for 9 characters at 99% significant level (Table S4),i.e.,TBL,HL,HW,HD,IO,E-S,E-N,A-G,and TD.Both Brown-Forsythe and Welch test further reveal significant difference between the new species andG.yunnanensis
in HD,IO,and E-N (Table S5).Genetically and morphologically,the newly identified matriline differed from the speciesG.yunnanensis
.Thus,we describe the new species of the genusGlyphoglossus
below.3.3.Taxonomic account
sp.n.
(Figures 3,4,5;Table 2)Holotype:
KIZA00254,an adult male,collected on 2 May 2010 from Huadianba (25°52’05’ N,100°01’05’ E,2920 m elevation,Cangshan Mountain,Dali Bai Autonomous Prefecture,Yunnan,China.Paratypes:
Twelve adult males,all from Yunnan,China:from the type locality,KIZA00253,KIZA00255–KIZA00258 collected on 2 May 2010,and KIZA00252 on 27 May 2010;from Lijiang City (26°51’18’ N,100°13’40’ E),KIZ 2014005754,KIZ 2014005781–KIZ 2014005785 collected in June 2010.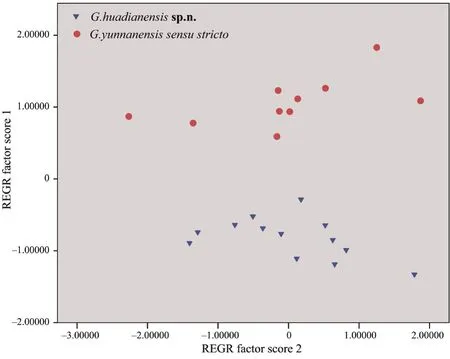
Figure 2 Scatterplot of principal components 1 and 2 of size-adjusted morphometric data for males of G.huadianensis sp.n. and G.yunnanensis sensu stricto.

Figure 3 Photographs of holotype of G.huadianensis sp.n.(KIZA00254)in life and its habitat,showing (A) dorsal view,(B) lateral view,(C) be in the wild,and (D) habitat.Photographs by Hong HUI.
Etymology:
The namehuadianensis
refers to Huadianba,the locality where the new species was first found.Its Chinese common name is“huā diàn xiǎo xiá kǒu wā”(花甸小狭口蛙).Diagnosis:
The new species ofGlyphoglossus
is medium-sized(average SVL 36.5 mm in adult males;body size of taxa ofGlyphoglossus
ranges from 26.9 ± 3.3 mm to 61.3 ± 0.7 mm;Table 4),and shares the following combination of characters with its congeners:flattened body;wide head;short snout;small eyes;maxillary and vomerine teeth present;paired dermal folds across palate;a crescentic,inner metatarsal tubercle present(Inger,1966;Manthey and Grossmann,1997;Parker 1934).The new species can be diagnosed from other congeners by the combination of the following characters:adult male body size larger than 34 mm (SVL range of 34.8–37.8 mm);pupil rounded;supratympanic fold distinct;tympanum concealed;toe tips obtuse;subarticular tubercles prominent and rounded;webbing between toes extensive,extending up to the second subarticular tubercle on toe IV;outer metatarsal tubercle present;dorsum tuberculate;and a pair of conspicuous large round spots in groin.
Description of holotype
(adult male) (measurements in mm):A medium-sized species ofGlyphoglossus
,SVL 37.3 mm;body stout ;head much broader than long (HW/HL ratio 1.42);snout rounded,projecting slightly beyond mandible in ventral view;canthus rostralis indistinct;loreal region oblique;nostrils rounded,upward positioned,slightly closer to tip of snout than to eye (E–N/E–S ratio 0.63);internarial distance slightly greater than eye to nostril distance (IN/E–N ratio 1.03);eye small (ED/HL ratio 0.38),its diameter greater than eye to nostril distance(ED/E–N ratio 1.59);interorbital distance broader than upper eyelid width (IO/UE ratio 1.26);maxillary teeth present;a feeble‘W’-shaped notch (=symphysial knob) on the anterior margin of mandible;mouth extends to posterior corner of eye;choanae visible;vomerine teeth present,positioning on two slightly sloping straight transverse ridges behind choanae;tongue oval,smooth,not nicked apically,free for approximately one third of its length;pupil rounded;tympanum concealed;single subgular vocal sac.Forelimbs more than half of body size (20.1 mm vs 37.3 mm);fingers free of web;inner three fingers (I,II,III) with skin fringes;relative length of fingers (measurements in parentheses,in mm):III (7.6) >II (3.8) >I (3.1) >IV (2.6);finger tips blunt;nuptial pads absent on fingers;subarticular tubercles prominent and rounded,subarticular tubercle formula:1:1:2:2;three metacarpal tubercles,distinct blade-like inner metacarpal tubercle longer than the outer,median smallest.
Hind limbs short (TBL/SVL ratio 0.42),and tibiotarsal articulation reaches up to angle of the jaw when extended forward along the body;heels untouched each other when legs at right angle to body;dorsal surfaces of thigh and tibia tuberculate;relative length of toes:IV >III >V >II >I;toes obtuse;webbing between toes extensive,webbing formula:I 0–1 II 0–1 III 0–1 IV 1–0 V,and the absent-web sides of phalanges have dermal fringe;subarticular tubercles distinct and rounded,subarticular tubercle formula:1:1:2:2:2;a large (~3.2 mm),crescentic inner metatarsal tubercle,that is longer than Toe I (2.0 mm),and an outer metatarsal tubercle small and compressed.
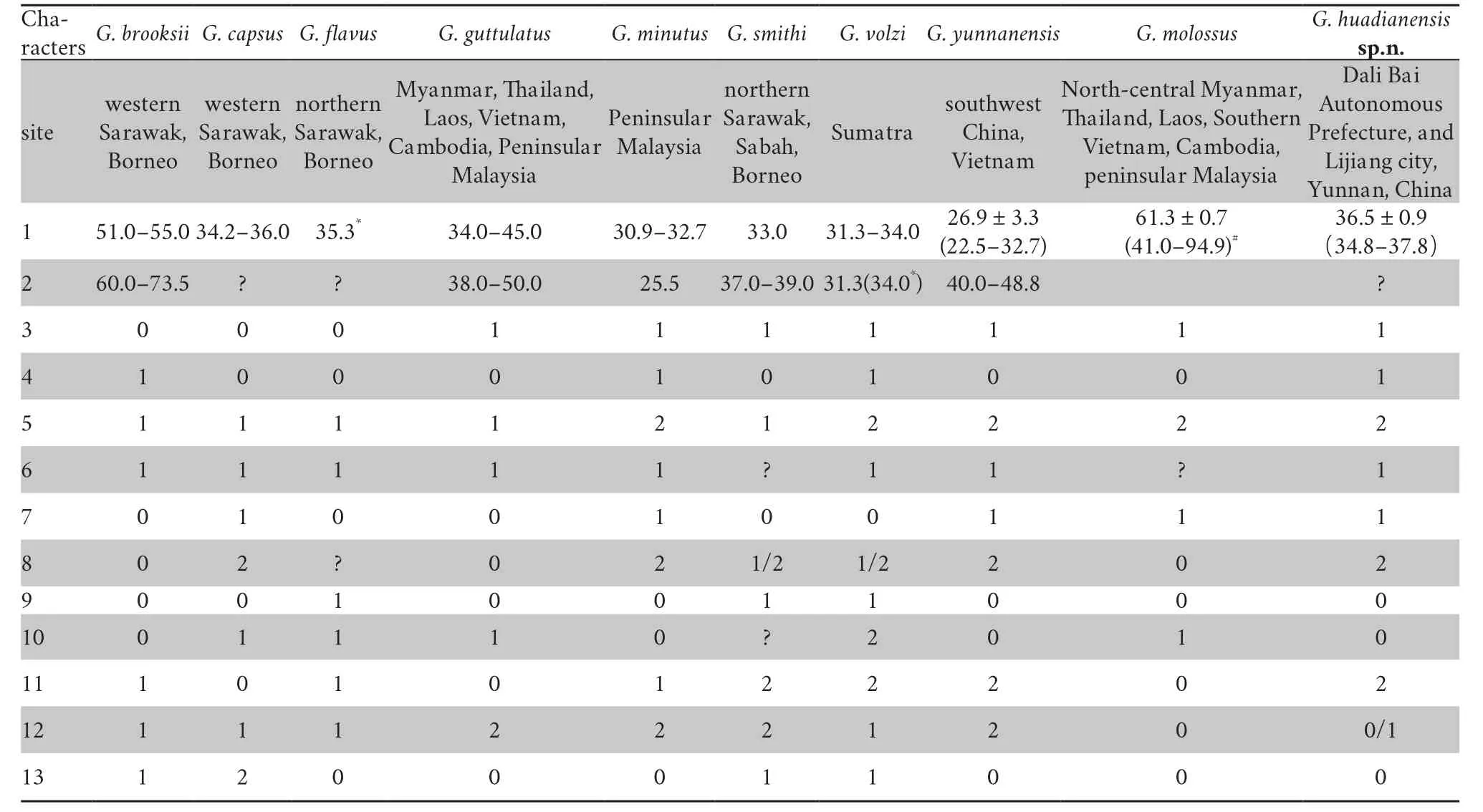
Table 4 Mensural and meristic data for Glyphoglossus huadianensis sp.n.,compared with congeneric species (adapted from Das et al.(2014) based on published descriptions of currently recognized species of Glyphoglossus).Characters:1.Male SVL (maximum,unless range given,in mm);2.Female SVL (maximum,unless range given,in mm);3.Supratympanic fold indistinct (0) or distinct (1);4.Dorsum smooth or granular (0) or tuberculate (1);5.Foot webbing absent(0),basal (1) or extensive (2);6.Lateral fringes on toes absent (0) or present (1);7.Outer metatarsal tubercle absent (0) or present (1);8.Pupil shape vertical (0),horizontal (1) or rounded (2);9.Tips of toes obtusely rounded (0) or expanded (1);10.Interorbital fold absent (0),indistinct (1) or distinct (2);11.Dorsum ground colour olive (0),yellow (1) or brown (2);12.Dorsum unpatterned (0),with small spots (1) or large dark central area (2);13.Flanks unpatterned (0),with dark blotches (1) or with red bars (2).In addition,'?' denotes an unknown character state;'/' denotes a multistate character;* denotes unknown sex;# denotes there is no sexual dimorphism in terms of size and shape for G.molossus.
Dorsum covered with slender tubercles or granules edged with black border,also small granules scattered over the head,and limbs;an indistinct and interrupted dark lateral band from the posterior of eye to middle of torso,which extend forward to canthus rostralis;supratympanic fold extending from posteriormost point of eye to angle of jaws;abdomen and inner side of limbs smooth.
Color of holotype in life:Dorsal surface yellowish-brown;supratympanic fold with an interrupted pale-edged dark line;loreal and tympanic region pale yellowish-brown,mottled with dark brown spots or patches on edge of upper jaw;flanks light yellow,mottled with brown patches;reticular markings on posterior of thighs and region around vent with a yellow background and brown border;one or two indistinct dark stripes on fore limb and tibia dorsally;a pair of conspicuous large round black spots in groin (4.6 mm × 3.7 mm),with yellow border.

Figure 4 Views of holotype of G.huadianensis sp.n. (KIZA00254) in preservative:dorsal (A),ventral view (B),lateral (C),posterior of thighs and region around vent (D),tympanum (E),mandible (F),vomerine teeth (G),features of dorsal skin (H),ventral view of hand (I) and foot (J).
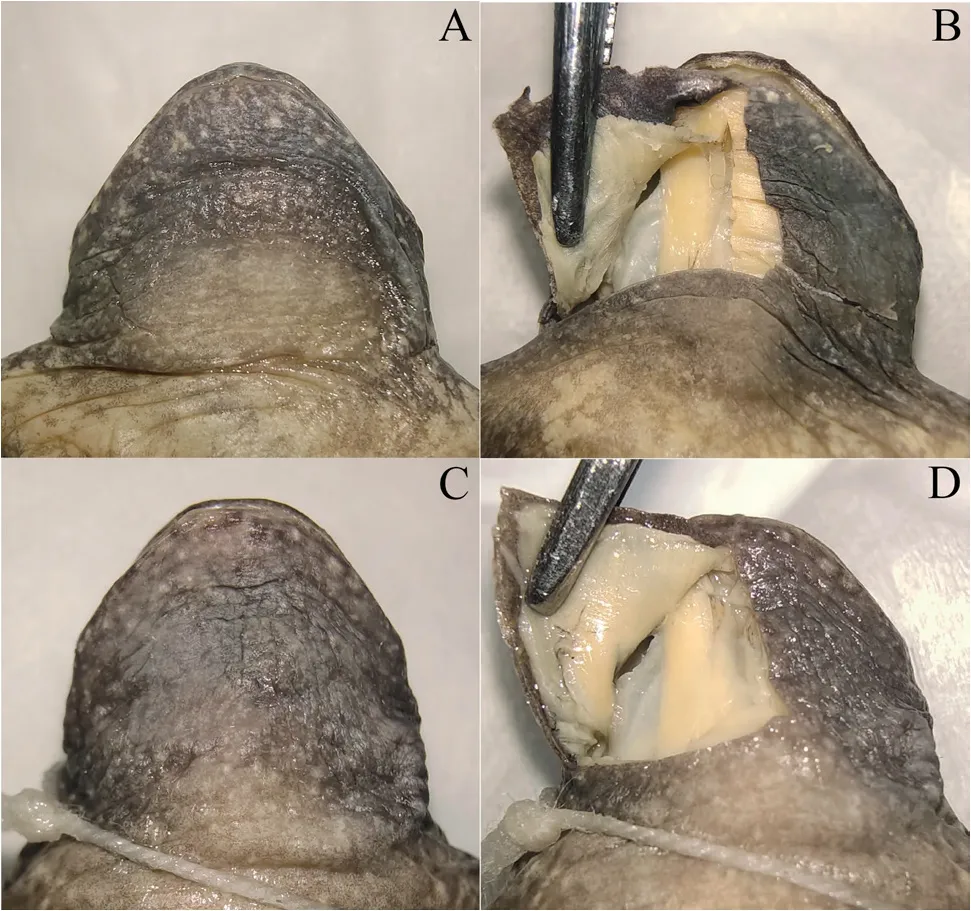
Figure 5 Vocal sac view of G.huadianensis sp.n. and G.yunnanensis:ventral view of gular skin (A) and anatomical structure of vocal sac (B)of G.yunnanensis; ventral view of gular skin (C) and anatomical structure of vocal sac (D) of G.huadianensis sp.n.
Variations:
Morphometric variation of the type series is shown in Table 2.Because type series are all males,sexual dimorphism of the new species could not be determined.Paratypes are generally similar to the holotype in body proportions,with only slight variation observed in three aspects.Firstly and most obviously,specimens from the type locality are differentiated from those from Lijiang City in the following respects (Figure S1):dorsal colouration (yellowish-brownvs.
earth yellow);shape of the dorsal pattern (irregular markingvs.
regular dotted stripes);pattern of posterior side of thighs and region around vent (with reticular markingsvs.
no reticular markings);and webbing formula:(I 0–1 II 0–1 III 0–1 IV 1–0 Vvs.
I 0–1 II 0–1 III 0–1IV 1–0 V).Also,there is 0.1% genetic divergence (p-distance,based on COI gene,Genebank accession number in Table S1) between these two populations.Secondly,there are only two metacarpal tubercles in four paratypes (KIZ 2014005781,KIZ 2014005784,KIZ 2014005754,KIZA00255).Thirdly,the subarticular tubercle formula of three paratypes (KIZA00252,KIZA00253,KIZA00256) is:1:1:2:3:2.Distribution and ecology:
So far,the new species is known from two localities:the type locality,Huadianba,Cangshan Mountain,Dali Bai Autonomous Prefecture,Yunnan Province,and Lijiang City,Yunnan Province.At the type locality,tracked their call,the new species was found in puddles in meadows(Figure 3).No females,tadpoles or eggs were found.Rana shuchinae
Liu,1950 was also encountered at the type locality.Comparisons:
The new species,Glyphoglossus huadianensis
sp.n.,has geographically and genetically closer relationship withG.yunnanensis
than with other known congeners according to recent work (Zhanget al
.,2020),but morphologically it can be significantly differentiated fromG.yunnanensis
(P
<0.01) based on one-way ANOVA test (Tables S4,S5):larger body size,SVL of 36.5 ± 0.9 mm (34.8–37.8 mm) in adult males (n
=13) (vs.
26.9 ±3.3 mm (22.5–32.7 mm) inG.yunnanensis
,n
=10);smaller ratios of head length (HL) (0.24vs.
0.29),head width (HW) (0.34vs.
0.37),head depth (HD) (0.19vs.
0.22),interorbital distance (IO)(0.08vs.
0.09),E-S (0.09vs.
0.11),E-N (0.06vs.
0.07),tibia length(TBL) (0.43vs.
0.45),tympanum diameter (TD) (0.06vs.
0.08) to body size (SVL);larger ratio of A-G (0.45vs.
0.41) to body size(SVL).Also,G.huadianensis
sp.n.
differs fromG.yunnanensis
by having tuberculate dorsum (vs.
dorsum smooth,except for some feebly raised glandular ridges which correspond with the outlines of the markings),having irregular markings/regular dotted stripe on dorsum (vs.
dorsum with a symmetrical vaseshaped brown marking,edged with darker and lighter,from between the eyes to the sacral region),having an indistinct and interrupted dark lateral band from the posterior of eye to middle of torso (vs.
a very distinct dark lateral band from the eye to the groin,bordered above by a series of more or less confluent black spots),having indistinct canthus rostralis(vs.
canthus rostralis distinct),having prominent subarticular tubercles (vs.
subarticular tubercles feeble prominent),having heels untouched each other when legs at right angle to body (vs.
heels feebly overlapping).The new species from Yunnan Province,western China differs from all other congeners in Southeast Asia in morphology (Table 4).Glyphoglossus brooksii
(Boulenger,1904),a Bornean endemic,distributed in western Sarawak (Malaysia),north-central and West Kalimantan (Indonesia) (IUCN SSC Amphibian Specialist Group 2019,and references therein).The new speciesG.huadianensis
sp.n.can be distinguished fromG.brooksii
by the following characters:smaller SVL of adult males 36.5 ± 0.9 mm (34.8–37.8 mm) (vs.
51–55 mm inG.brooksii
:Inger,1966;Parker,1934);supratympanic fold distinct (vs.
indistinct);foot webbing extensive (vs.
basal);outer metatarsal tubercle present (vs.
absent);pupil round (vs.
vertically elliptical);dorsum yellow-brown/earth yellow and no spots on the sides of the body (vs.
dorsum yellow,with two dark stripes or rows of spots,and flanks with small black blotches).Glyphoglossus flavus
(Kiew,1984) is distributed in Borneo,Gunung Mulu National Park (Sarawak),Danum Valley Conservation Area (Sabah),and Ulu Temburon National Park(Brunei) (IUCN SSC Amphibian Specialist Group,2018,and references therein);G.huadianensis
sp.n.differs fromG.flavus
by dorsum tuberculate (vs.
smooth);foot webbing extensive (vs.
basal);outer metatarsal tubercle present (vs.
absent);tips of toes obtusely rounded (vs.
expanded);dorsum yellow-brown/earth yellow and flanks without stripe (vs.
orange-yellow,and flanks with black stripes).Glyphoglossus guttulatus
(Blyth,1856) is distributed in Southern Myanmar,most of mainland Thailand,the Central Highlands of Vietnam,Laos and south-western Cambodia(Frost,2020);G.huadianensis
sp.n.differs fromG.guttulatus
in the following characters:smaller SVL of adult males 36.5 ± 0.9 mm(34.8–37.8 mm) (vs.
34.0–45.0 mm inG.guttulatus
:Manthey and Grossmann,1997);foot webbing extensive,extending up to the second subarticular tubercle on toe IV (vs.
basally webbed);outer metatarsal tubercle present (vs.
absent);pupil round (vs.
vertical);dorsum tuberculate (vs.
dorsum smooth with distinct blackedged brown irregular markings).Glyphoglossus minutus
(Daset al
.,2004) is distributed in Peninsular Malaysia;G.huadianensis
sp.n.differs fromG.minutus
in the following characters:larger SVL of adult males 36.5 ± 0.9 mm (34.8–37.8 mm) (vs.
30.9–32.7 mm inG.minutus
);irregular markings/regular dotted stripe on dorsum (vs.
a triangular mark,wide anteriorly in interorbital region,and a darker variegation and a large,dark,central area on dorsum);tympanum concealed (vs.
present);loreal region oblique (vs.
vertical);finger tips blunt (vs.
pointed);relative length of fingers:III >II >I >IV (vs.
III >II >IV >I);relative length of toes:IV >III >V >II >I (vs.
IV >V >III >II >I).Glyphoglossus smithi
(Barbour and Noble,1916) occurs in northern Sarawak and Sabah,Malaysia,and Sumatra,Indonesia;G.huadianensis
sp.n.differs fromG.smithi
in the following characters:larger SVL of adult males 36.5 ± 0.9 mm (34.8–37.8 mm) (vs.
33.0 mm inG.smithi
:Manthey and Grossmann,1997);foot webbing extensive (vs.
basally webbed);outer metatarsal tubercle present (vs.
absent);toe tips obtusely rounded (vs.
expanded);dorsum yellow-brown/earth yellow and tuberculate (vs.
dorsum smooth and dark brownish grey);flanks without spots (vs.
flanks with symmetrical black blotches,edged with pinkish);reticular markings around vent (vs.
a red X-shaped spot above the vent).Glyphoglossus volzi
(Van Kampen,1905) occurs in northwestern and south-eastern Sumatra,Indonesia;G.huadianensis
sp.n.differs fromG.volzi
in the following characters:larger SVL of adult males 36.5 ± 0.9 mm (34.8–37.8 mm) (vs.
31.3–34.0 mm inG.volzi
);outer metatarsal tubercle present (vs.
absent);toe tips obtusely rounded (vs.
expanded);interorbital dermal fold absent(vs.
distinct);dorsum yellow-brown/earth yellow and flanks without spots (vs.
dorsum reddish-brown with black spots,and flanks with dark blotches).Glyphoglossus capsus
(Daset al.
,2014) is distributed in Matang Range in Sarawak,Malaysia,and the upper reaches of Gunung Penrissen (the boundary between Sarawak (Malaysia) and Kalimantan (Indonesia)).G.huadianensis
sp.n.differs fromG.capsus
in the following characters:supratympanic fold distinct(vs.
indistinct);foot webbing extensive (vs.
basally webbed);dorsum yellow-brown/earth yellow and flanks without spots(vs.
dorsum olive with red spots and venter with a large area of red).Glyphoglossus molossus
Günther,1869,a large,globose frog,is found in north-central Myanmar,most of mainland Thailand,Laos,and southern Vietnam;G.huadianensis
sp.n.differs fromG.molossus
in the following characters:smaller SVL of adult males 36.5 ± 0.9 mm (34.8–37.8 mm) (vs.
SVL 61.3 ± 0.7 mm (41.0–94.9 mm) inG.molossus
,Laojumponet al.
2012);dorsum yellowbrown/earth yellow and tuberculate (vs.
dorsum brownish olive and smooth,with large dark central area);pupil round (vs.
vertical);snout rounded (vs.
an unusually truncate snout:snout very short and blunt covered by a leathery finely granular skin;the fleshy part of the lower jaw swollen,truncated in front,forming a semicircular disk).An additional species has been described in the genus-Calluella ocellata
Liu (1950),from“Szekuaipa,Chaochiaohsien,Sikang,7800 feet”(currently,Sikuaiba,Zhaojue County,Sichuan Province,China).The study of Zhanget al
.(2020) included samples from Yanziluo village,Jiefang township,Zhaojue County,Sichuan Province,China,3129 m,and discovered the morphological characteristics of these specimens were consistent with the original description ofCalluella ocellata
,but they clustered together (member of lineage C),and were nested among specimens ofG.yunnanensis
.That is to say,the results supported Liu and Hu (1961):Calluella ocellata
is a synonymy ofG.yunnanensis
Boulenger 1919.Moreover,G.huadianensis
sp.n.(lineage LD) was deeply separated from lineages C with a divergence of 9.2% (Zhanget al
.,2020).So,no further morphological comparison was conducted.4.Discussion
Zhanget al
.(2020) investigated the geographical distribution and pattern of genetic variation ofG.yunnanensis
,and discovered one of maternal lineages (LD) is deeply divergent from the remaining ones (W,C,E) (Figure 1),suggested it may be an undescribed species.In the study,morphological comparisons and genetic divergence clearly support this possibility and demonstrate that the lineage LD is a new species,namedG.huadianensis
sp.n.,and is distinctive fromG.yunnanensis
and other known congeners.Firstly,based on 2154 bp concatenated mitochondrial gene fragments,the genetic distance betweenG.huadianensis
sp.n.and theG.yunnanensis sensu stricto
(lineages W+C+E) was 8.9%,and the difference betweenG.huadianensis
sp.n.and other known congeners was 7.8%–20.0% (Table 1).Secondly,the result of PCA and ANOVA indicates thatG.huadianensis
sp.n.differs fromG.yunnanensis
significantly.As well,G.huadianensis
sp.n.separated from other known congeners by a series of morphometric characters(Table 4).One main factor probably hindered the discovery of the new species described here:there are obvious intraspecific variation in the color pattern and body size among populations ofG.yunnanensis
(personal observation).
G.huadianensis
sp.n.was superficially morphologically similar with specimens from some populations ofG.yunnanensis
,easily misleading the identifications if made without detailed morphological and genetic examination.In future investigations,data on advertisement calls and larvae may be particularly helpful in resolving the taxonomic uncertainty.In addition toG.yunnanensis
,the new taxa described herein is the second species known from Yunnan,China in the genusGlyphoglossus.
The discovery of the new species indicates that the diversification ofGlyphoglossus
in China still needs further investigation.Moreover,the new species andG.yunnanensis
were found to co-distribute in Dali Bai Autonomous Prefecture and Lijiang City,which suggests that these regions may play an important role in the formation and evolution of species diversity ofGlyphoglossus
.Among specimens ofG.huadianensis
sp.n.,distinct morphological variation occurs between populations Dali and Lijiang in dorsal colouration,shape of the dorsal pattern,pattern of posterior side of thighs and region around vent,and webbing formula.But the genetic distance between them was 0.1%,which was much less than interspecific differentiation.These evidences indicated variation between them are only intraspecific differentiation.To date,the new species was only found in Dali Bai Autonomous Prefecture and Lijiang City,and more research is needed on its life history information and distribution to determine its conservation level.
Acknowledgements We would like to thank G.H.YU and J.S.WANG for their help in the initial study,and express sincerely appreciation to K.JIANG for his valuable help.We also thank the reviewers for their valuable comments on the manuscript,and J.Z.WANG and D.C.OUYANG for assistance in the field.This work was supported by grants from National Key Research and Development Program,Grant/Award Number:2017YFC0 505202;the National Natural Science Foundation of China (31301872 to D.R.ZHANG;31372165,31970404 and 31772424 to D.Q.RAO;31501870 and 31872216 to L.X.ZHANG);State Key Laboratory of Genetics Resources and Evolution (Grant No.GREKF17–08);and Biodiversity Survey,Monitoring and Assessment program of Ministry of Ecology and Environment of China (2019HB2096001006).
Appendix
Table S1 Information on Voucher No.,GenBank No.,and localities of specimens used in this study.
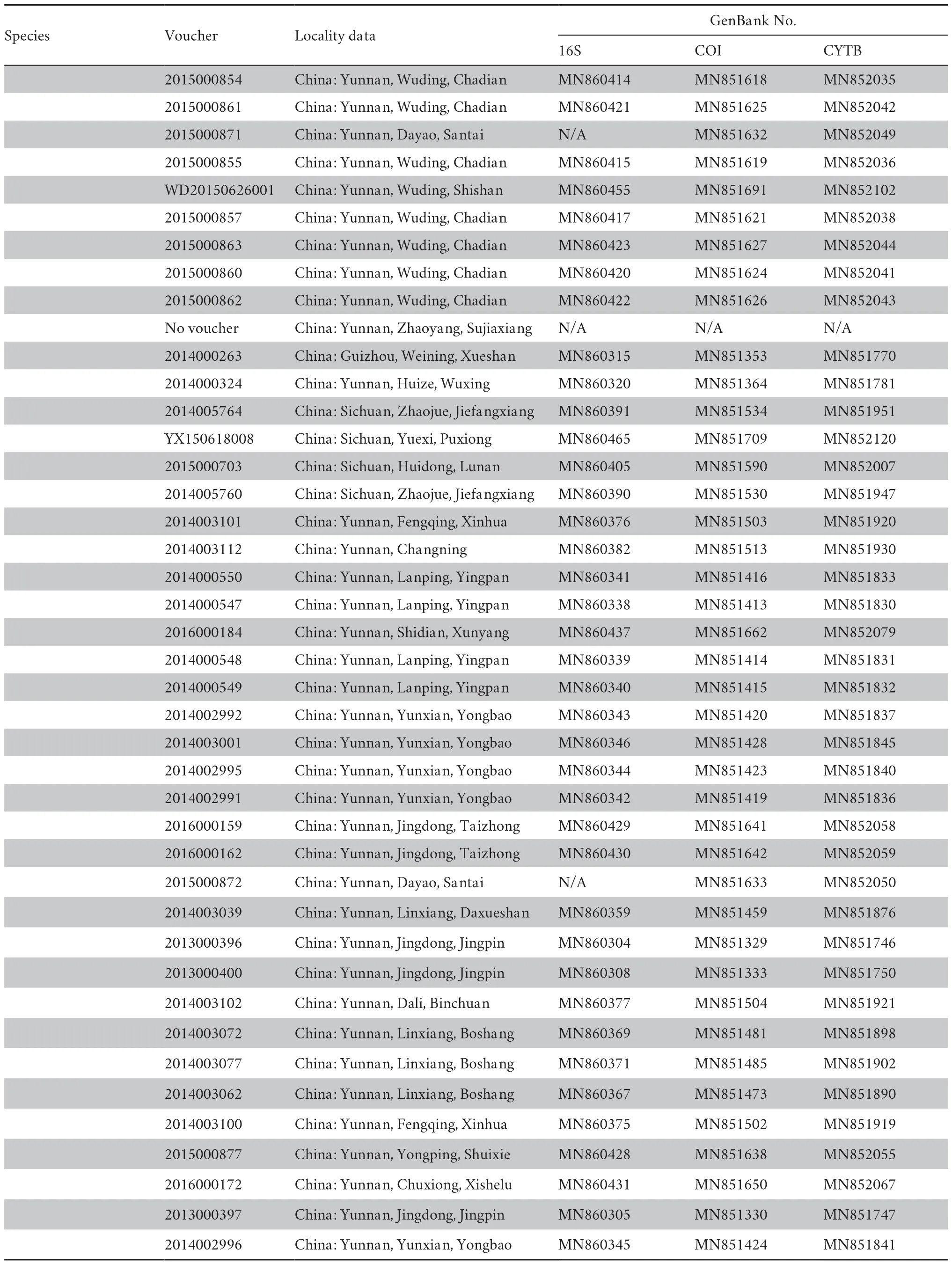
(Continued Table S1)
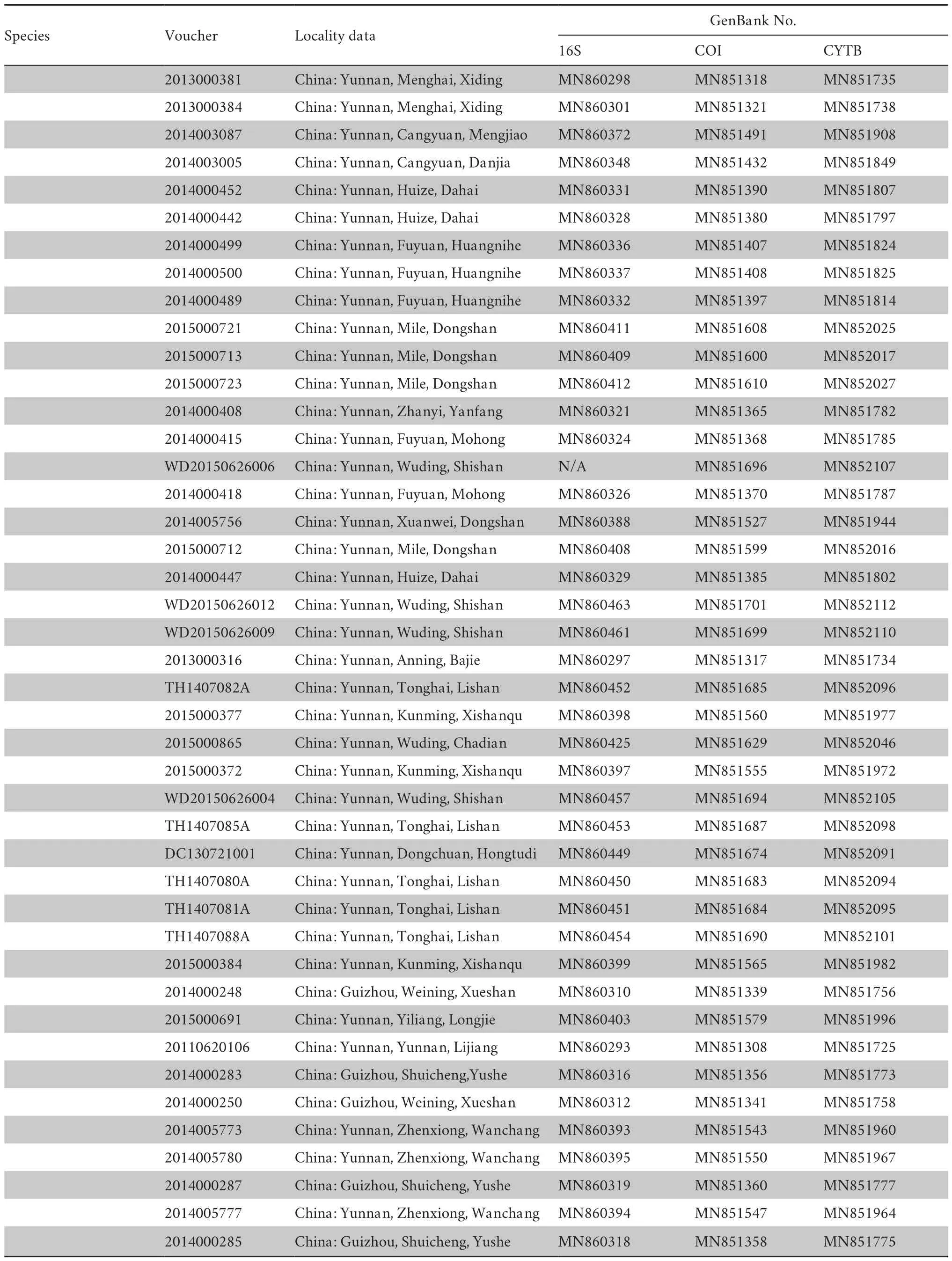
(Continued Table S1)

(Continued Table S1)
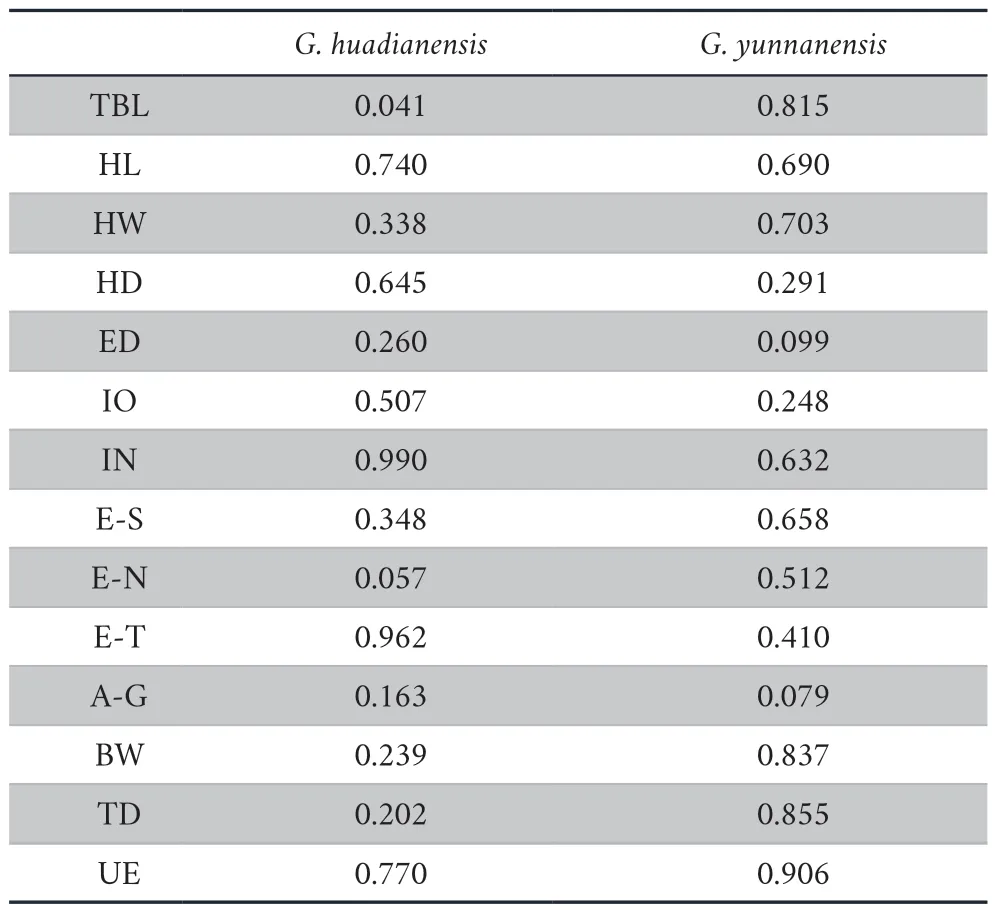
Table S2 Test of Normality based on Shapiro-Wilk Test (P <0.01).
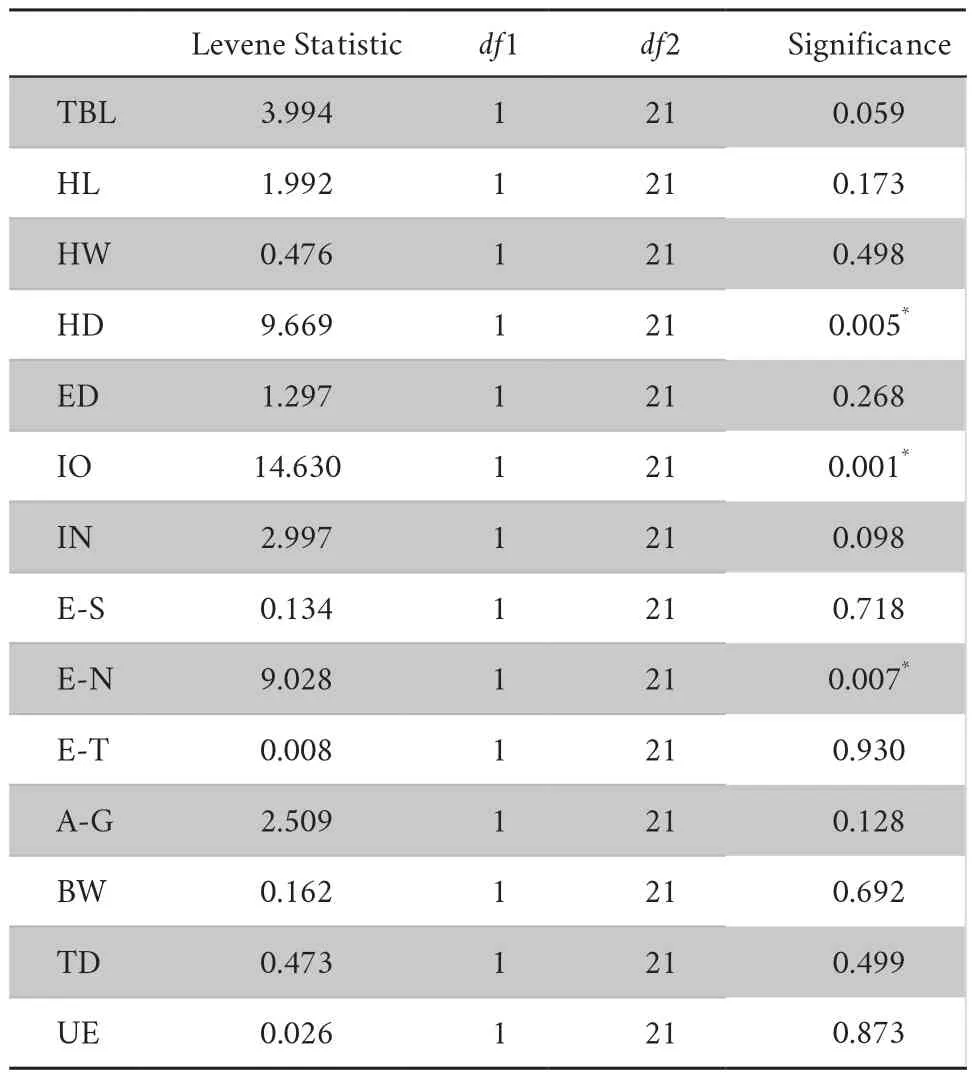
Table S3 Test of Homogeneity of Variances (P <0.01).
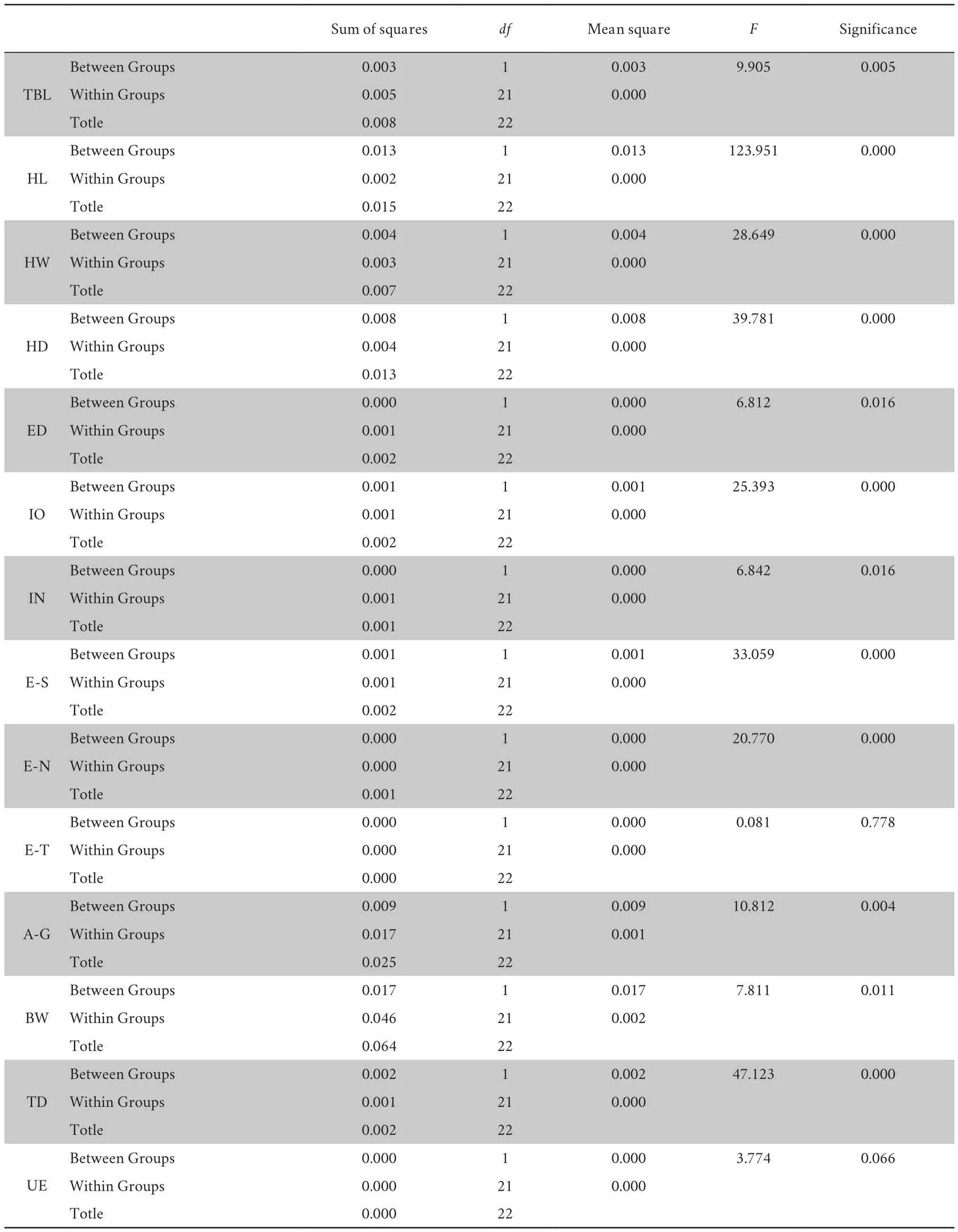
Table S4 Analysis of variance (ANOVA) at one way.
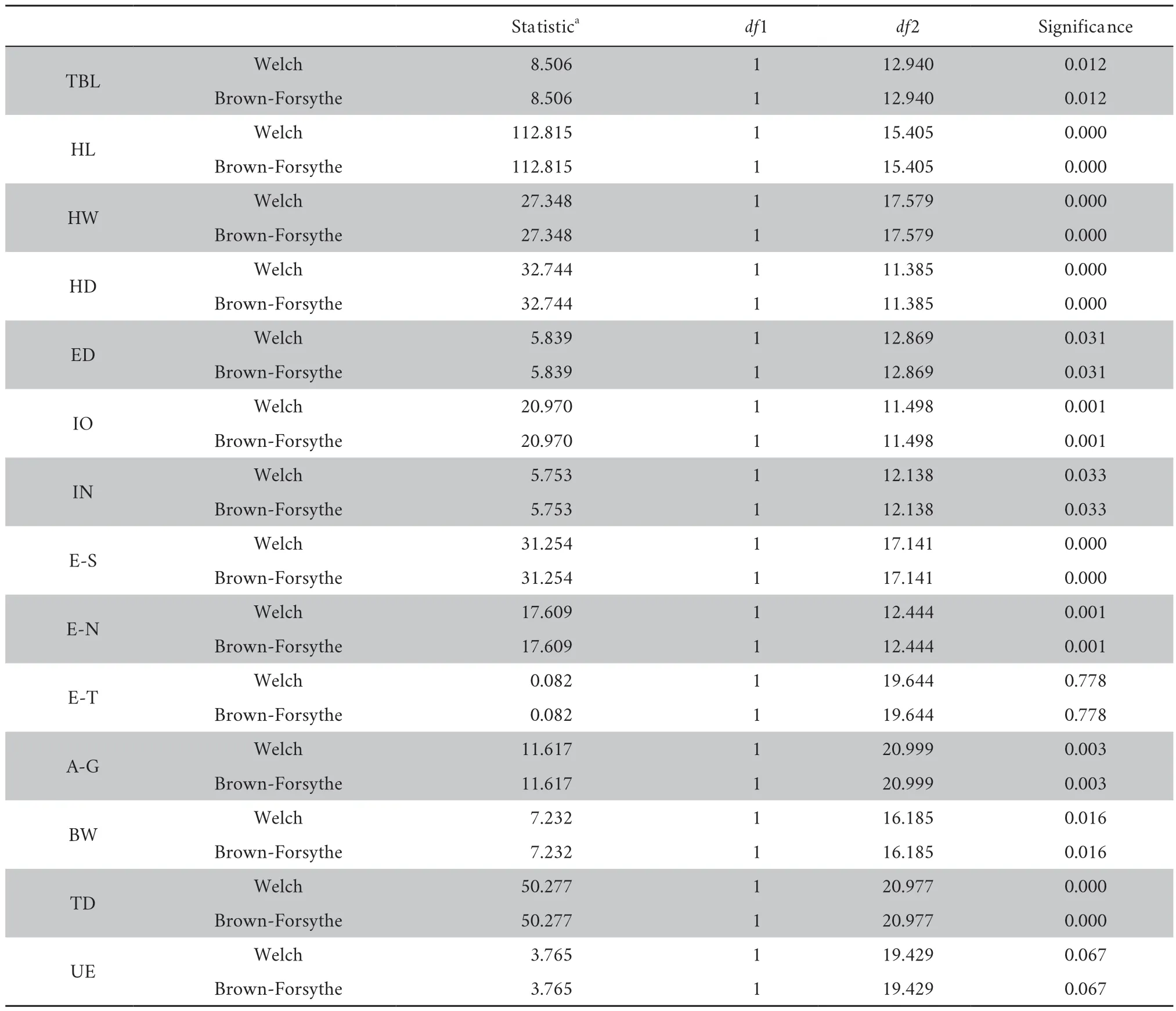
Table S5 Results of both Brown-Forsythe and Welch test.

Figure S1 Photographs of individuals of G.huadianensis sp.n.from two localities:(A) dorsal view,(B) ventral view:the upper from Dali Bai Automotous Prefecture,and the lower from Lijiang City.
杂志排行

Asian Herpetological Research的其它文章
- DNA Barcoding and Systematic Review of Minervaryan Frogs (Dicroglossidae:Minervarya) of Peninsular India:Resolution of a Taxonomic Conundrum with Description of a New Species
- A New Species of Odorous Frog Genus Odorrana (Anura,Ranidae) from Southern Guizhou Province,China
- Mitogenomic Conservation Genetics of the Critically Endangered Liaoning Clawed Salamander
- Chemical Composition and Crystal Structure of the Eggshell of the Green Crested Lizard Bronchocela cristatella (Agamidae)