不同耐盐性燕麦对盐胁迫的生理响应
2021-12-29王苗苗赵桂琴梁国玲柴继宽李娟宁周向睿
王苗苗,赵桂琴,梁国玲,柴继宽,李娟宁,周向睿
(1. 甘肃农业大学草业学院 / 草业生态系统教育部重点实验室 / 中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070;2. 青海大学畜牧兽医科学院 / 青海省畜牧兽医科学院 / 青海省青藏高原优良牧草种质资源利用重点实验室,青海 西宁 810016)
近年来,全球气候的变化、化肥的不合理施用和不科学的灌溉方法,导致全球土壤盐渍化程度越来越严重,我国的盐渍土面积也在逐年增加[1-2]。而盐渍化土壤对牧草及饲料作物的生产和种植产生了较大影响,因此,培育优良的耐盐牧草及作物是当今改良盐渍土的有效手段之一。高浓度盐胁迫会抑制植物的生长发育,植物体内吸收过量的Na+会打破细胞离子平衡,从而引起活性氧(reactive oxygen species, ROS)升高并对植物造成次级毒害[3-4]。同时盐胁迫会间接抑制叶片的光合作用,破坏细胞膜结构和抗氧化防御体系,并导致植物体内脯氨酸、可溶性糖等有机渗透调节物质大量积累,从而缓解盐胁迫对其造成的伤害[5]。
植物处于幼苗期时对于外界环境的变化较敏感,能够产生一系列反应来抵抗盐害[6]。在水稻(Oryza sativa)的研究中发现随着盐胁迫时间的延长,超氧化物歧化酶(superoxide dismutase, SOD)活性、丙二醛(malondialdehyde, MDA)、脯氨酸及可溶性糖含量上升,且耐盐水稻的过氧化物清除能力和渗透调节能力较强[7];孟祥浩等[8]研究显示,随着盐胁迫时间的延长,小麦(Triticum aestium)叶片中SOD 活性、可溶性蛋白含量降低,MDA 含量、游离脯氨酸含量与之相反,而过氧化物酶(peroxidase,POD)活性呈先增加后减少的趋势,且在胁迫1 d 时出现峰值;高彩婷等[9]对裸燕麦(Avena nuda)进行短期盐胁迫后发现,随胁迫时间的延长,裸燕麦丙二醛含量、可溶性糖含量、SOD、POD 活性总体呈先上升后下降随后又上升的趋势,而过氧化氢酶(catalase, CAT)活性先升高后降低,24 h 时出现最大值。综上所述,随着盐胁迫时间的延长,不同物种对盐胁迫的生理响应有一定差异。
燕麦(Avena sativa)是禾本科燕麦属的一年生草本植物,在生态条件脆弱地区是不可替代的特色粮饲作物[10-11],广泛分布于世界各地,种植面积在全球范围内位居第6 位。燕麦作为禾本科植物中耐盐性较好的作物之一,被广泛认为是改良盐渍土的替代作物,且不同燕麦品种之间的耐盐性差别较大[12]。目前,对于燕麦耐盐生理响应方面的研究多集中在不同盐浓度下抗氧化酶活性[13]、渗透调节物质变化规律[14]以及外源物质增强耐盐效应机制[15]等方面。对于比较不同耐盐性燕麦随盐胁迫时间延长的生理响应研究较少。研究不同耐盐性燕麦品种响应盐胁迫的生理差异,对燕麦耐盐育种和改良盐渍土具有重要意义。青永久195 和709 生育期100~120 d,株高140~150 cm,周散型穗,有效分蘖数为2~3 个。本研究采用150 mmol·L−1NaCl 进行盐胁迫,分析在同一浓度处理下燕麦随胁迫时间延长的光合参数、抗氧化酶活性、渗透调节物质及K+、Na+吸收的变化差异,并探讨燕麦响应盐胁迫的生理机制,以期为后续盐胁迫下对燕麦进行转录组测序、筛选燕麦耐盐基因和探究其耐盐的分子机理提供参考依据。
1 材料与方法
1.1 试验材料
根据前期燕麦苗期耐盐性综合评价筛选出耐盐材料青永久195[16],由燕麦种质资源耐盐性筛选得到盐敏感材料709,两种材料均由甘肃农业大学草业学院提供。试验于2019 年在甘肃农业大学草业学院实验室进行。
1.2 试验设计
挑选籽粒饱满的燕麦种子,用75%乙醇浸泡1 min 进行消毒,然后用蒸馏水冲洗干净,播种在装有沙子的育苗杯(直径9 cm、高13 cm)中,置于光照培养室[光照强度为1 200 μmol·(m2·s)−1],每天光照16 h,昼夜温度为(23 ± 1) ℃ / (20 ± 1) ℃,相对湿度为55%。萌发前每天浇灌适量蒸馏水,待长出两片真叶时,每隔2 d 浇灌1 L Hoagland 营养液,以保证幼苗生长。培养3 周后开始在营养液中加入150 mmol·L−1NaCl 溶液进行胁迫处理,分别在处理后的0 (CK)、6、24、72 h,取叶片(从上到下数第1~2 片叶)和根系,置于-80 ℃冰箱中保存,用于相关生理指标测定,每个处理3 次重复。
1.3 测定内容与方法
1.3.1 叶绿素含量及光合参数的测定
叶绿素含量:称取0.2 g 样品剪碎后置于具塞试管中,加入95%乙醇在黑暗中提取24 h,在波长665、649 和470 nm 下测定消光度[17]。
光合参数采用LI-6400 便携式光合仪(LI-COR,USA)测定,于晴天上午09:30 - 11:00,选取处理0、24、72 h 的不同燕麦材料叶片测其气孔导度(stomatal conductance, Gs)、 净光合速率(net photosynthetic rate, Pn)、胞间CO2浓度(intercellular carbon dioxide concentration, Ci)以及蒸腾速率(transpiration rate, Tr),每个处理3 次重复。光照强度为800 μmol·(m2·s)−1,CO2浓度为390 μmol·mol−1。
1.3.2 抗氧化酶活性及丙二醛含量的测定
参照张治安和陈展宇[18]的方法制备粗酶液,即称取0.5 g 燕麦幼苗待测叶片置于研钵中,加入50 mmol·L−1磷酸缓冲液(pH 7.8) 5 mL (研钵和磷酸缓冲液均提前预冷),在冰浴中研磨至匀浆。于4 ℃、12 000 r·min−1条件下离心10 min,取上清液进行相关抗氧化酶活性的测定。CAT 活性采用紫外吸收法测定[18];SOD 活性采用氮蓝四唑比色法测定[18];POD 活性采用愈创木酚比色法测定[18]。
MDA 含量采用硫代巴比妥酸法测定[18]。
1.3.3 渗透调节物质含量的测定
脯氨酸含量采用茚三酮比色法测定[18],可溶性糖含量采用蒽酮比色法测定[18],K+、Na+含量采用火焰光度计法测定[19-20]。
1.4 数据处理
通过计算平均值和标准误对供试燕麦材料的各处理性状进行单因素方差分析,分别利用SPSS 22.0 软件和Excel 2016 进行数据统计分析及制图。
2 结果与分析
2.1 盐胁迫对燕麦幼苗叶绿素含量的影响
盐胁迫使供试材料的叶绿素含量降低,且叶绿素含量在两种材料间差异不显著(P> 0.05) (图1)。与对照相比,胁迫24 h 后,青永久195 和709 的叶绿素含量分别下降了2.55%和4.38%,但差异未达显著水平(P> 0.05);胁迫72 h 后,两种材料的叶绿素含量较对照分别下降了13.88%和18.42%,且差异显著(P< 0.05)。

图1 不同盐胁迫时间下燕麦叶片的叶绿素含量Figure 1 Chlorophyll content of oat leaves under different salt stress times
2.2 盐胁迫对燕麦幼苗光合特性的影响
盐胁迫使两种材料的净光合速率均有所下降(图2)。胁迫24 和72 h 后,709 的净光合速率小于青永久195,随着处理时间的延长,709 的净光合速率下降幅度大于青永久195,且净光合速率两种材料间差异显著(P< 0.05);盐胁迫24 和72 h 后,青永久195 较CK 相比净光合速率下降了5.82%和26.35%,而709 则分别下降了39.67%和68.80%。盐胁迫后,两种材料的蒸腾速率和气孔导度与净光合速率的变化趋势一致,各胁迫时间处理后,青永久195 的蒸腾速率和气孔导度均大于709,且两种材料间差异显著(P< 0.05)。胁迫24 和72 h 后,青永久195 较CK相比蒸腾速率下降了5.05%和49.87%,气孔导度下降了16.91%和37.38%;而709 较CK 相比蒸腾速率下降了14.59%和56.10%,气孔导度下降了23.76%和44.67%。随着处理时间的延长,青永久195 和709的胞间CO2浓度均下降。胁迫24 h 后,青永久195和709 的胞间CO2浓度分别下降了8.01%和8.81%,但两种材料间差异不显著(P> 0.05);胁迫72 h 后,青永久195 和709 的胞间CO2浓度分别下降了24.99%和19.19%,且两种材料间差异显著(P< 0.05)。

图2 不同盐胁迫时间下燕麦叶片的光合特性Figure 2 Photosynthetic characteristics of oat leaves under different salt stress times
2.3 盐胁迫对燕麦幼苗抗氧化酶活性及丙二醛含量的影响
随着盐胁迫时间的延长,青永久195 可以保持相对较高的POD、SOD 活性(图3)。其中,供试材料叶片的POD 活性随胁迫时间的延长呈先升高后下降的趋势,青永久195 在24 h 时达到最大,而709在6 h 时达到峰值;盐胁迫6、24、72 h 后,青永久195的POD 活性分别升高了21.59%、27.11%和17.34%,709 的POD 活性分别升高了28.85%、24.51%和11.65%,但POD 活性两种材料间差异均不显著(P> 0.05)。SOD 活性的变化趋势与POD 活性类似,在24 h 时达到峰值,后又下降,且耐盐材料青永久195 的SOD 活性均大于盐敏感材料709。随着处理时间的延长,二者叶片CAT 活性均升高,且盐胁迫6、24、72 h 后青永久195 的CAT 活性均大于709;盐胁迫24、72 h 后,CAT 活性在两种材料间差异显著(P<0.05);盐胁迫6、24、72 h 后,青永久195 的CAT 活性分别升高了39.17%、216.28%和309.89%,709 的CAT活性升高了18.88%、132.51%和169.82%。随胁迫时间的延长,MDA 含量均有所升高,且耐盐材料青永久195 的MDA 含量低于盐敏感材料709。其中,盐胁迫6、24、72 h 后青永久195 的MDA 含量分别升高了21.39%、42.75%和66.29%,709 的MDA 含量分别升高了20.50%、44.99%和75.99%。
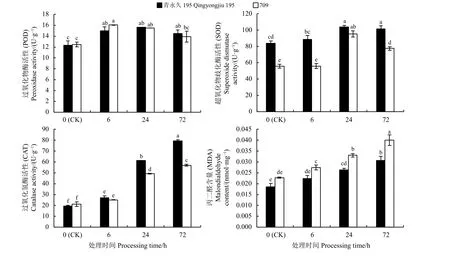
图3 不同盐胁迫时间下燕麦叶片的抗氧化酶活性及丙二醛含量Figure 3 Antioxidant enzyme activity and malondialdehyde content of oat leaves under different salt stress times
2.4 盐胁迫对燕麦幼苗渗透调节物质含量的影响
2.4.1 盐胁迫对燕麦幼苗可溶性糖和脯氨酸含量的影响
盐胁迫下,燕麦幼苗的可溶性糖含量高于CK,且盐胁迫24、72 h 后,可溶性糖含量在两种材料间差异显著(P< 0.05) (图4)。青永久195 的脯氨酸和可溶性糖含量随胁迫时间的延长先上升后下降,均在24 h 时达到峰值。709 的脯氨酸含量在6 h 时与CK 相比无显著差异(P> 0.05),24 h 时略有增加,而72 h 时明显下降(P< 0.05);709 的可溶性糖含量随时间呈先上升后下降的趋势,在6 h 时达到最大。在不同时间处理下,耐盐材料青永久195 的脯氨酸和可溶性糖含量均大于盐敏感材料709。
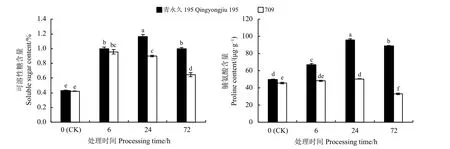
图4 不同盐胁迫时间下燕麦叶片的脯氨酸和可溶性糖含量Figure 4 Proline content and soluble sugar content of oat leaves under different salt stress times
2.4.2 盐胁迫对燕麦根和叶中K+、Na+含量的影响
正常条件下,燕麦幼苗叶片中K+含量大于根系,但Na+含量在根和叶中均较少,且在两个品种上表现一致(图5)。随处理时间的延长,两个材料根和叶中K+含量均呈下降趋势。其中,盐胁迫6 和24 h时,青永久195 根中K+含量均低于709,且差异显著(P< 0.05),而叶中K+含量高于709,差异不显著(P> 0.05);盐胁迫72 h 时,青永久195 根和叶中K+含量均与709 基本持平,无显著差异(P> 0.05)。

图5 不同盐胁迫时间下燕麦根和叶中K+、Na+含量Figure 5 K+ and Na+ content in root and leaf of oats under different salt stress times
随盐胁迫时间的延长,青永久195 和709 的根和叶中Na+含量呈增加趋势。盐胁迫6 h 时,青永久195和709 根中Na+含量基本相同,而青永久195 叶中Na+含量低于709,差异不显著(P> 0.05);盐胁迫24 h 时,青永久195 根和叶中Na+含量均低于709,根中差异显著(P< 0.05),而叶中差异不显著(P> 0.05);盐胁迫72 h 时,青永久195 根中Na+含量显著高于709 (P<0.05),而叶中Na+含量显著低于709 (P< 0.05) (图5)。
3 讨论
3.1 盐胁迫对燕麦光合作用的影响
叶绿素是植物进行光合作用的重要物质,叶绿素含量通常是衡量植物对盐胁迫耐受能力的指标之一,且大量研究表明,盐胁迫下耐盐品种叶绿素含量高于盐敏感品种[21]。王旭明等[22]研究发现,水稻(Oryza sativa)叶绿素含量随土壤盐浓度的增加呈降低的趋势,耐盐水稻品种叶绿素含量显著高于盐敏感水稻品种。本研究中,供试燕麦材料随盐胁迫时间的延长,叶绿素含量降低,且耐盐材料青永久195 的降幅小于盐敏感材料709,表明盐胁迫对燕麦叶绿体结构造成了一定伤害,耐盐材料对盐胁迫的耐受能力大于盐敏感材料。盐胁迫24 h 时叶绿素含量降低速率较慢,与无胁迫对照相比差异不显著,但在胁迫72 h 时叶绿素含量降低速率较快。这可能是因为盐胁迫初期燕麦叶片的叶绿体结构损伤较轻,叶绿素合成速率未受影响。盐胁迫对植物的光合作用有抑制作用,会降低植物的光合速率,光合电子传递也会受到抑制,进而产生光抑制,最后使植物产量下降甚至死亡[23]。本研究中,盐胁迫24 h时Pn、Tr、Gs、Ci降幅较小,72 h 时两种材料的光合参数显著降低,且耐盐材料青永久195 的Pn、Tr、Gs均高于盐敏感材料709,与叶绿素含量的变化趋势一致。可能是因为盐胁迫诱导燕麦体内光合相关基因表达,使其光合参数降低、速率减缓,从而减轻了盐胁迫对燕麦光合器官的损伤。
3.2 盐胁迫对燕麦抗氧化酶活性和丙二醛含量的影响
SOD 在植物体内主要起清除自由基的作用,受到盐胁迫时SOD 活性有所上升;POD 主要参与催化化合物的氧化反应;CAT 能有效清除对植物体内细胞有氧化作用的过氧化氢[24-25]。董杰等[26]采用300 mmol·L−1NaCl 分别处理小麦(Triticum aestivum)24、72 h 后,结果表明,SOD 活性在处理24 h 后明显增加,72 h 后呈现下降的趋势,而POD、CAT 活性随盐胁迫时间的延长持续增加。孙璐[27]在对高粱(Sorghum bicolor)的研究中发现,盐胁迫下SOD、POD、CAT 等抗氧化酶活性均高于正常水平,从而抵抗盐胁迫对其造成的氧化伤害。本研究中,随着盐胁迫时间的延长,两种材料SOD 和POD 活性呈先升高后下降的趋势,青永久195 在24 h 时达到最大,在72 h 时略微下降;CAT 活性升高,且在胁迫6、24、72 h时耐盐材料青永久195 酶活性高于盐敏感材料709。说明燕麦在盐胁迫初期通过维持较高的SOD 和POD 酶活性来抵御NaCl 对其细胞造成的伤害,随着胁迫时间延长,可能使燕麦体内活性氧水平升高,SOD、POD 合成受到抑制,这时通过CAT活性升高来缓解盐胁迫对植物的伤害,这与Wu 等[28]的研究结果相似。
MDA 含量可反映植物组织内脂质过氧化的程度,间接反映细胞膜损伤程度,其含量越高说明细胞膜受损越严重,抗逆境胁迫能力越弱[29]。刘文瑜等[30]研究发现,NaCl 胁迫后,藜麦(Chenopodium quinoa)叶片MDA 含量显著增加,加剧了膜脂过氧化程度并造成细胞膜系统损伤。本研究中,盐胁迫6 h 时两种材料的MDA 含量增幅较小,青永久195的MDA 含量与对照相比差异不显著,但709 的MDA 含量与对照相比差异显著,在胁迫24、72 h 时MDA 含量增幅较大,两种材料的MDA 含量均与对照差异显著,这可能是因为胁迫初期燕麦幼苗叶片受伤害程度低,能够开启自身的渗透调节保护机制以减轻盐胁迫的影响,导致MDA 含量升高幅度较小,这一点也体现在盐胁迫后叶绿素含量的变化上,与许晓萱[31]在玉米(Zea mays)上的研究结果一致。
3.3 盐胁迫对燕麦渗透调节物质含量的影响
脯氨酸、可溶性糖等渗透调节物质在稳定质膜、保护质膜的完整性、参与多种生理生化代谢过程中发挥着重要作用。陈晓晶等[32]研究发现,燕麦三叶期经盐胁迫后,叶片脯氨酸含量升高。李珍等[33]研究发现盐胁迫使新麦草(Psathyrostachys juncea)脯氨酸和可溶性糖含量增加。本研究中,青永久195 和709 经盐胁迫后脯氨酸和可溶性糖含量均呈先升高后降低的趋势,说明在盐胁迫初期,燕麦能够通过增加有机渗透调节物质含量来维持细胞渗透势的平衡,当胁迫时间延长,细胞渗透调节能力逐渐下降,从而影响燕麦苗期的生长。青永久195 的脯氨酸和可溶性糖含量在24 h 时达到最大,但709 的可溶性糖含量在6 h 时达到最大,可能是因为盐敏感材料在盐胁迫开始,合成的可溶性糖作为其主要渗透调节物质来维持植物正常生长,这一结果与高彩婷等[9]的研究结果类似。
植物能否保持较高的K+/ Na+比是其耐盐能力强弱的体现[34]。姜瑛等[17]研究发现,随着胁迫浓度的升高和时间的延长,燕麦叶片中Na+含量增加而K+含量降低。刘晓龙等[35]研究发现,盐胁迫后水稻叶片中Na+积累量显著增加而K+积累量显著降低。本研究中,青永久195 和709 随盐胁迫时间的延长,根系和叶片中Na+含量显著增加而K+含量显著降低,说明盐胁迫抑制了燕麦对K+的吸收。海霞等[36]研究表明,盐胁迫下燕麦地上部Na+积累量明显高于根系,燕麦根系没有明显贮存Na+的作用。本研究则表明,盐胁迫6、24、72 h 时,青永久195 根系中Na+积累量高于叶片,而709 经盐胁迫6、24 h时根系中Na+积累量高于叶片,胁迫72 h 时,叶片中Na+积累量高于根系,说明盐胁迫后耐盐材料青永久195 可以将Na+贮存在根系中以缓解Na+对自身的伤害,同时也证明了耐盐材料对盐离子的转运能力较强。本研究还发现盐胁迫72 h 时,青永久195 根系中K+含量下降幅度低于709,说明了盐胁迫下耐盐材料阻止K+流失的能力优于盐敏感材料,这可能与K+转运蛋白有关,但具体的分子机理有待进一步研究。
盐胁迫后燕麦在不同处理时间下的生理响应,可以推测其相关基因表达调控的变化。在盐胁迫6、24 h 时供试材料MDA 含量的增幅比72 h 小,这可能是因为盐胁迫初期可诱导SOD、CAT 和POD酶相关基因及其他耐盐基因的表达,使抗氧化酶活性升高,从而减缓MDA 的合成,这也体现在光合参数、脯氨酸含量以及可溶糖含量的变化方面。盐胁迫后,青永久195 根系中Na+积累量高于叶片,且胁迫72 h 时青永久195 根系中K+含量下降幅度低于709,而在胁迫72 h 时,709 叶片中Na+积累量高于根系,说明青永久195 能够限制Na+向地上部运输,同时维持较高的K+含量,这可能是由于青永久195 经盐胁迫诱导后HKT、SOS1等离子转运相关基因的表达调控发生了变化,从而使青永久195 表现出较好的渗透调节能力、维持较高的生物膜稳定性和抗氧化酶活性,最终表现出较好的耐盐性。综上所述,供试材料对盐胁迫生理响应的不同,也可能导致耐盐相关基因的差异表达,为挖掘燕麦与盐胁迫相关差异表达基因提供依据。
4 结论
随盐胁迫时间的延长,青永久195 和709 的叶绿素含量、Pn、Tr、Gs、Ci、K+含量均有所下降;POD、SOD 活性、脯氨酸和可溶性糖含量呈先升高后降低的趋势,CAT 活性、MDA 含量、Na+含量均有所增加。但盐胁迫下青永久195 叶片的SOD 活性、CAT活性、脯氨酸含量和可溶性糖含量均高于709,MDA含量低于709,且叶片Na+积累量低于709。说明耐盐材料青永久195 在盐胁迫下渗透调节能力较强,可以维持较高的生物膜稳定性和抗氧化酶活性,从而抵御一定的盐胁迫,保证植物正常生长。
