中国沙棘和云南沙棘的遗传分化及遗传多样性
2021-12-27王天翼王罗云张建国曾艳飞
王天翼,徐 悦,王罗云,张建国,曾艳飞
(中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091)
沙棘(Hippophae rhamnoidesLinn.),又名醋柳、酸刺、黑刺等,属胡颓子科(Eleagnaceae)沙棘属(Hippophae),是二倍体(2n=24),落叶灌木或小乔木,雌雄异株,风媒传粉;主要分布于欧亚大陆,集中在温带和寒温带地区,生态幅极广[1-4]。沙棘具有很高的营养价值和经济价值,在开发保健食品、药品和化妆品上有着独特的作用,还是很多偏远地区脱贫致富的经济来源。更重要的是,沙棘具有很高的生态价值,被广泛应用在治理荒漠化、退耕还林、植被绿化等方面[4-5]。
沙棘共包括了9个亚种[2-3],起源于青藏高原,后多次扩散至亚欧大陆其他地区[6],其中,中国沙棘(H.rhamnoidessubsp.sinensis)和云南沙棘(H.rhamnoidessubsp.yunnanensis)是我国特有的沙棘亚种。中国沙棘叶常近对生,成狭披针形或矩圆披针形,叶背面银白色、被鳞片,从西南到东北都有自然种群分布[7],同时它也是我国栽培和种植历史最悠久的一个亚种[8]。云南沙棘叶互生,基部最宽,常为圆形或有时楔形,叶背面灰褐色、被锈色鳞片,其分布范围相对狭窄,多分布于西藏、四川和云南的交界地带[7]。依据国家植物标本资源库(http://www.cvh.ac.cn/)、Global Biodiversity Information Facility(GBIF,https://www.gbif.org/)、国家资源标本平台(http://nsii.org.cn/2017/)和相关沙棘研究文献等资料记载的沙棘分布点,中国沙棘和云南沙棘在地理上有分类存在疑问的区域,主要集中在四川省北部和中西部。中国沙棘和云南沙棘形态相似,并且具有不完全谱系分选,亲缘关系较近,区分难度较大[8]。判定该区域种群基因流的变化、种群间的分界线,有利于明确遗传扩散适应机制[9]。目前,有关中国沙棘和云南沙棘谱系地理的研究文献以及标本资源库对于四川省北部和中西部沙棘的种类归属问题存在一定的分歧[8,10-11]。
本研究中,笔者利用18个SSR 标记对来自32个自然种群的352个中国沙棘和云南沙棘个体和沙棘属其他种和亚种的75个个体进行遗传变异分析,以期探讨以下问题:(1)中国沙棘和云南沙棘是否存在明显的遗传分化?二者的物理分界线在哪?(2)中国沙棘和云南沙棘的遗传多样性分布如何?
1 材料与方法
1.1 样本采集
从中国沙棘和云南沙棘的典型分布地区和分类存疑地区采集了32个自然种群,共352个个体。因沙棘存在克隆繁殖,为了避免取到同一基因型个体,采集时不同样品间至少间隔200 m。对所采集的个体进行形态学观察以便初步定种,并记录定位(表1)。为了明确中国沙棘和云南沙棘的系统发育关系,还收集了沙棘属其他种及亚种分别5~10个共75个个体用于分析(表1),包括蒙古沙棘(H.rhamnoidessubsp.mongolica)、溪生沙棘(H.rhamnoidessubsp.fluviatilis)、高加索沙棘(H.rhamnoidessubsp.caucasica)、中亚沙棘(H.rhamnoidessubsp.turkestanica)、海滨沙棘(H.rhamnoidessubsp.rhamnoides)、西藏沙棘(H.tibetana)、肋果沙棘(H.neurocarpa)、棱果沙棘(H.goniocarpa)、江孜沙棘(H.gyantsensis)、柳叶沙棘(H.salicofolia)。所有收集到的叶片均用变色硅胶干燥保存。

表1 沙棘种群采样信息Table 1 Sampling information of sea buckthorn populations
1.2 DNA 提取和SSR 基因型检测
叶片总DNA 的提取采用CTAB 植物基因组DNA 快速提取试剂盒(北京艾德莱生物公司)。从沙棘基因组和转录组开发的SSR 分子标记[12]中选取18 对对所有个体进行PCR 扩增,反应总体系为20 μL,反应体系和反应程序参照王罗云等[13]的工作进行。
用ABI 3730 测序仪(美国生命技术公司)对PCR 产物分析,内标选择Liz500,结果经由软件Genemapper 4.0(美国生命技术公司)分析,对于SSR 基因型完全一致的潜在克隆株个体仅保留一份样品,所得到数据均以大果沙棘品种“向阳”为阳性对照。
1.3 系统发育分析
两两样品间的遗传距离由软件MSA 4.05[14]计算,再用Phylip 3.695[15]软件中的Neighbor 程序对所有沙棘个体以及中国沙棘和云南沙棘分别构建系统发育树,自展值设置成1 000 次。
1.4 遗传结构分析
利用Structure 2.3[16]软件确定所采集中国沙棘和云南沙棘个体的遗传分组情况。采用混合模型和等位基因频率关联模型,分组值K设置为1~10,每个K值重复计算10 次,每次运行100 000 次Burnin和1000000次迭代[13]。将结果上传到Structure Harvester(http://taylor0.biology.ucla.edu/structureHa rvester/)中,根据lnP(D)和ΔK最大原则[17]选择合适的K值,再通过Distruct 1.1[18]将结果可视化。
1.5 遗传多样性和遗传分化分析
利用软件GenAlEx 6.503[19],计算中国沙棘和云南沙棘种群的遗传多样性参数,包括等位基因个数NA、有效等位基因个数NE、观测杂合度HO、期望杂合度HE和近交系数FIS。利用MSA 4.05[14]计算遗传分化系数FST,并进行分子方差分级分析(Analysis of Molecular Variance,AMOVA)。
2 结果
2.1 中国沙棘和云南沙棘的系统发育关系及遗传结构
基于SSR 基因型构建的沙棘属的5个种和7个亚种的系统发育树结果(图1A)显示:所有的中国沙棘都聚为一大支,而云南沙棘个体主要聚为4 支;除一支及少量个体与中国沙棘聚在一起外,其他3 支都和中国沙棘距离较远,其中,1 支由12个云南沙棘个体(图1A 虚线部分)组成的分支与沙棘属的其他4个种——西藏沙棘、肋果沙棘、江孜沙棘和柳叶沙棘聚在一起,这12个个体包括来自BS种群1个、XZD种群1个、XJ种群1个、BX种群7个、WL种群2个。中国沙棘和云南沙棘的个体单独构建的系统发育树(图1B)显示:除6个云南沙棘个体(LX种群2个、LTB种群2个、CY种群1个、BS种群1个)与中国沙棘种群交叉聚在一起外,其余个体明显地分为2 部分,各分支支持率大都不高(小于50%);云南沙棘个体可分为a、b 2 支,其中,a 支为种群的个体与中国沙棘聚在一起,b 支为西藏、云南和四川中南部与中国沙棘距离较远;中国沙棘各分支支长较短,差异不明显,云南沙棘各分支支长较长,遗传距离较远。
利用Structure 2.3[16]软件,基于中国沙棘和云南沙棘个体SSR 基因型的分组分析显示:lnP(D)在K= 2 时迅速增高,K>2 后趋于平缓(图2A);ΔK在K= 2 时有明显峰值(图2B)。因此,所有个体分2 组最合适,对应与按初步形态分为中国沙棘和云南沙棘2个亚种(图2C)。位于四川的MWS 和QY 2个种群的大部分个体表现为这2 组的中间基因型(图2C),因此,后续分析将把MWS、QY 作为中间型种群。
2.2 中国沙棘和云南沙棘的遗传多样性
基于18个SSR 位点的中国沙棘遗传多样性参数(表2)表明:NA为1.7~4.9(平均3.5),NE为1.4~2.7(平均2.3),HO为0.213~0.528(平均0.402),HE为0.266~0.539(平均0.462)。基于18个SSR 位点的云南沙棘遗传多样性参数如下(表2)表明:NA为1.4~4.2(平均2.7)、NE为1.3~2.4(平均1.8)、HO为0.056~0.331(平均0.202)、HE为0.127~0.490(平均0.334)。中国沙棘和云南沙棘中HO最高的种群分别是YY 和KD,最低的种群分别为TW和PM。中国沙棘的NA、NE、HO、HE均值均高于云南沙棘。
2.3 中国沙棘和云南沙棘的遗传分化
基于18 对SSR 标记,中国沙棘两两种群间的FST为0.026~0.286,分化程度最低的为种群CFA和WC,分化程度最高的为种群TW 和HY;云南沙棘两两种群间的FST为0.069~0.539,分化程度最低的为种群LX 和SJS,分化程度最高的为种群LZ 和SJS;云南沙棘的FST值、FIS值普遍高于中国沙棘(表2、3)。AMOVA 分析显示(表4)显示:中国沙棘和云南沙棘亚种间遗传变异占比为23.7%,亚种内不同种群间占23.3%,种群内个体间占53.0%。中国沙棘25.5% 的遗传变异来自种群间,74.5%的遗传变异来自种群内个体间;云南沙棘种群36.7%的遗传变异来自种群间,63.3%的遗传变异来自种群内个体间。
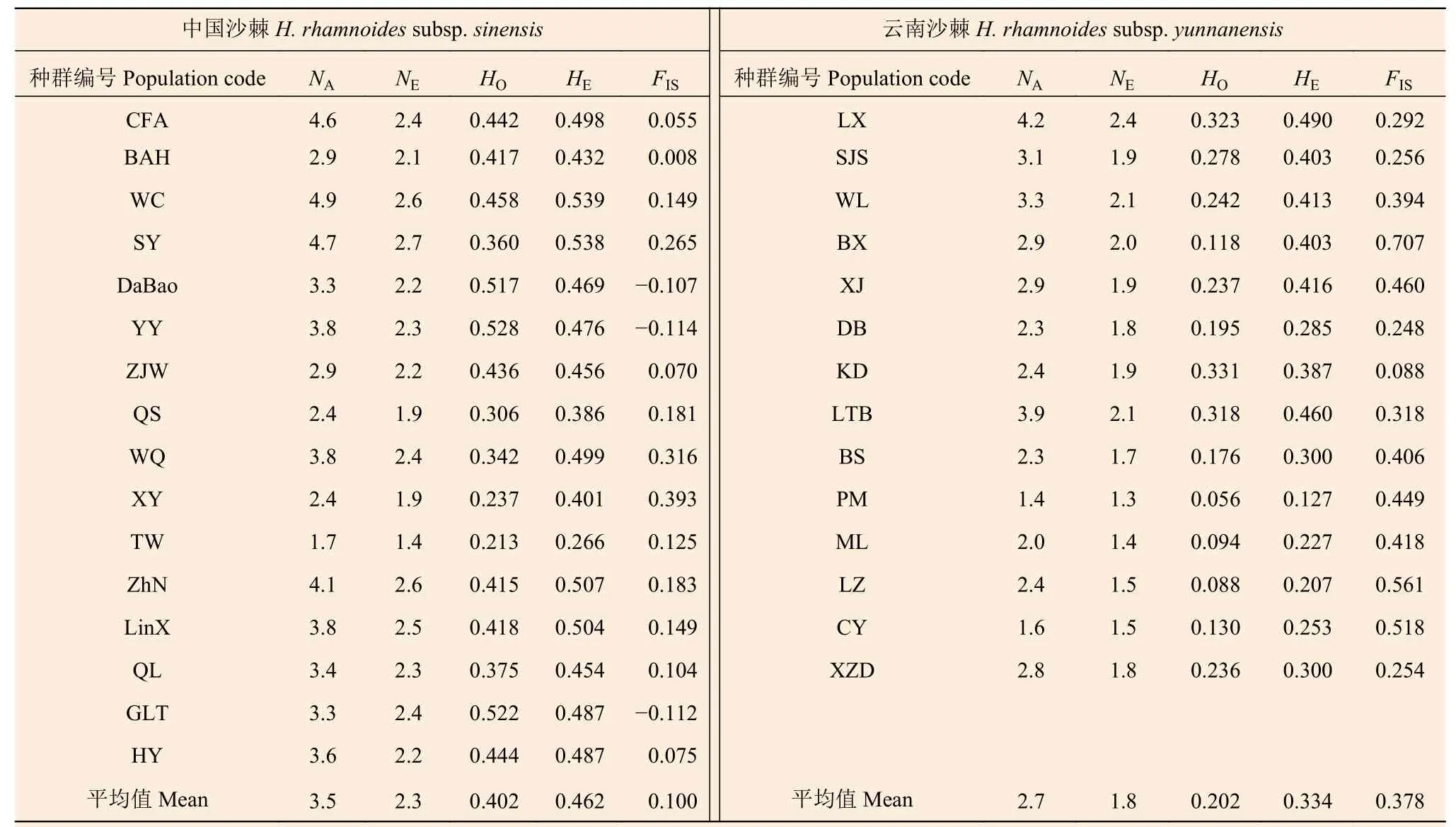
表2 本研究分析的30个沙棘种群的遗传多样性参数Table 2 The genetic diversity of 30 sea buckthorn populations
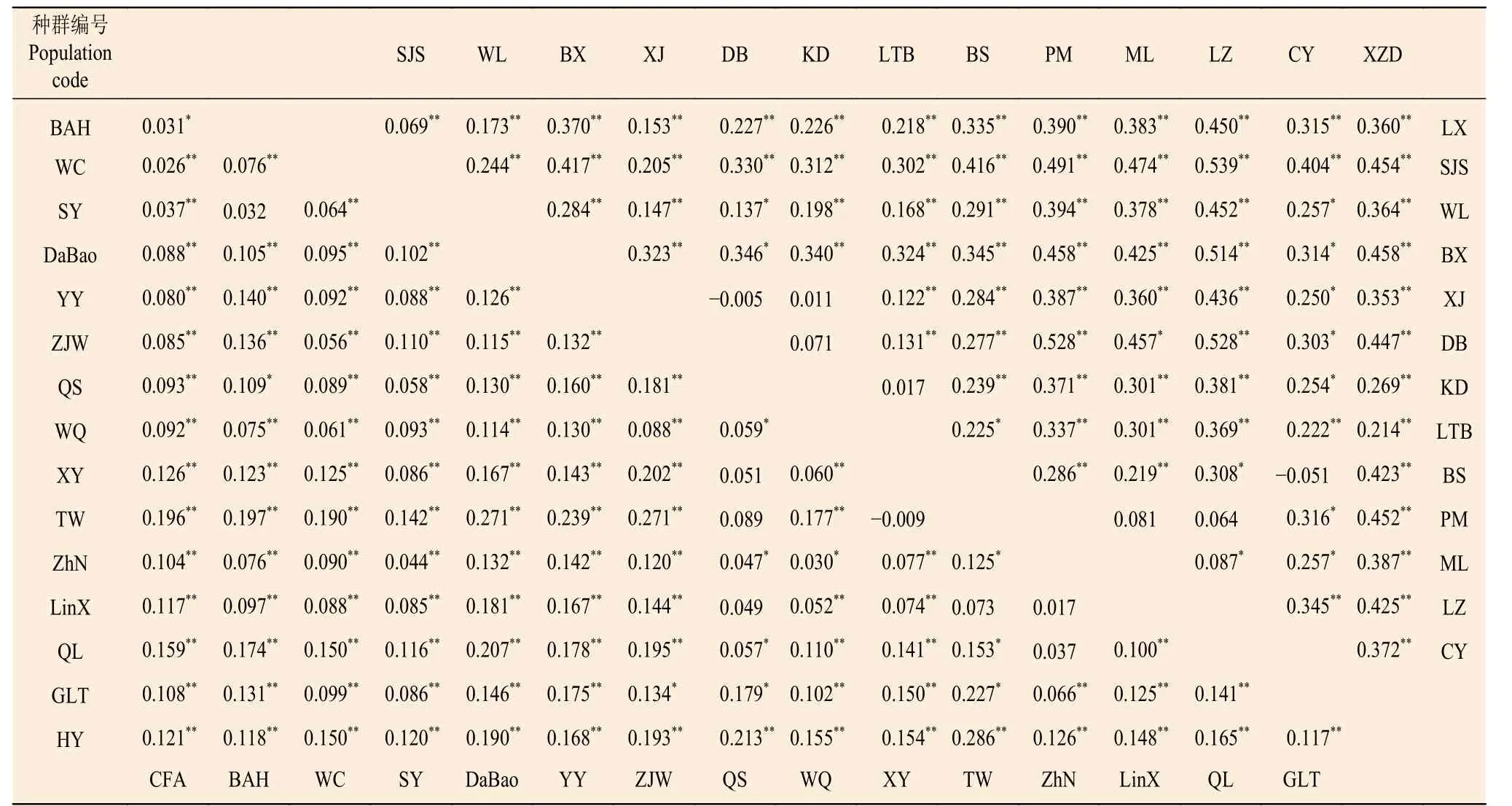
表3 中国沙棘(对角线左下方)和云南沙棘(对角线右上方)亚种内不同种群间遗传分化系数(FST)Table 3 Genetic differentiation coefficients (FST) between populations within H.rhamnoides subsp.sinensis (below diagonal) and within H.rhamnoides subsp.yunnanensis (above diagonal),respectively.
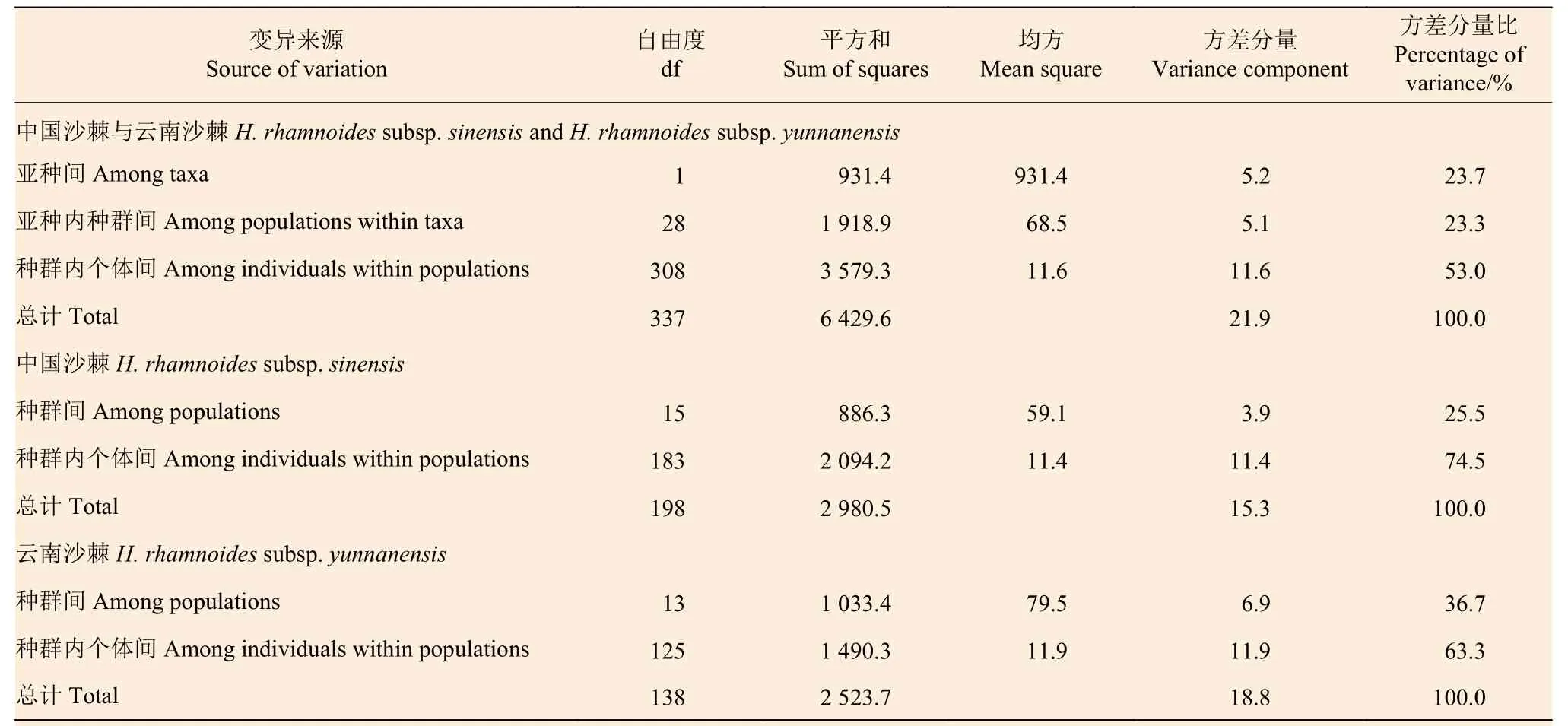
表4 中国沙棘和云南沙棘基于SSR 基因型的AMOVA 分析Table 4 AMOVA analysis based on SSR genotypes of H.rhamnoides subsp.sinensis and H.rhamnoides subsp.yunnanensis
3 讨论
3.1 中国沙棘和云南沙棘的“边界线”
依据标本、文献资料的记载,在四川省部分地区,中国沙棘和云南沙棘的分类存在争议。本研究基于18个SSR 位点的基因型,系统发育树和Structure 2.3[16]软件分组结果都显示,中国沙棘和云南沙棘可分成明显的2 组,种群MWS 和QY 的大部分个体表现为2 组间杂交。这2个种群分别位于四川北部的红原县和松潘县,该区域北侧种群的遗传组成中含更多的中国沙棘遗传成份,南侧种群的遗传组成中含更多的云南沙棘遗传成份,因此,推测该地区为中国沙棘和云南沙棘的分布边界带。Wang等[8]利用叶绿体片段序列研究中国沙棘的空间遗传分布时,发现重叠分布区的中国沙棘的2个单倍型也存在于云南沙棘中,并且单倍型在若尔盖盆地有明显的分界。这与本研究确定的分布边界带区域相符,这一结果可为沙棘的物种鉴定分类提供初步依据。要进一步确定分界线的地理位置,还需要更多的数据,如形态学数据、地形数据、气候数据等。
中国沙棘和云南沙棘的系统发育树中,分支支持率低,可能存在种间杂交;同一亚种内部各分支支持率大多不高,同一地理种群个体并不能完全聚在一起,说明地理种群间存在基因流;Structure 结果也表明,亚种间有基因渐渗现象,这与前人研究结果一致[6,11]。中国沙棘与云南沙棘亲缘关系较近,亚种间和种群间可能存在基因交流。但要进一步明确基因交流的程度,还需要更多的SSR 位点等遗传信息的支持,以及覆盖范围更广的样品收集。
3.2 中国沙棘和云南沙棘的遗传分化和遗传多样性
孙坤等[20]利用ISSR 标记研究了不同海拔中国沙棘的遗传变异情况,发现产生遗传分化的主要原因可能是基因流受限。本研究中,依据AMOVA和遗传分化系数,发现云南沙棘的遗传分化程度高于中国沙棘,造成这一结果的主要原因可能是云南沙棘采集自四川、云南、西藏等地形复杂地区,复杂的地形和气候使种群间的基因交流受限。另一个可能是,某些地区的云南沙棘种群可能与某种沙棘,如肋果沙棘[2],存在种间杂交,杂交后代的存在或杂交导致的基因渐渗使得这些种群与其他地区的云南沙棘种群的遗传差异升高。
沙棘被推测为起源于青藏高原地区,青藏高原的隆升和第三纪气候影响了它的分布范围和多样性,而后扩散到欧洲和中国其他地区[11,21]。中国沙棘在我国境内分布广,从西藏、云南到辽宁、内蒙均有自然种群分布,生境差异较大。通常分布范围广的物种有着更高的遗传多样性,而遗传多样性越高,越能为其适应当地环境提供有力支持[22-24]。本研究中,中国沙棘的各项遗传多样性参数均高于云南沙棘,代表中国沙棘的遗传多样性高于云南沙棘种群,这与前人研究结论一致[8,13],这可能是由于中国沙棘相较于云南沙棘分布范围广。
4 结论
本研究探讨了中国沙棘和云南沙棘的系统发育关系,发现中国沙棘的遗传多样性高于云南沙棘,与前者更广泛的分布范围有关;云南沙棘种群的遗传分化程度高于中国沙棘种群,推测是复杂的高山地形限制了云南沙棘种群间的基因交流,或区域性种间杂交渐渗提高了不同地理种群间的差异;确定了中国沙棘和云南沙棘的分布边界位于四川北部的红原县、松潘县一带。本工作为中国沙棘和云南沙棘遗传资源的保护和利用提供了重要的基础。
