重度火烧迹地兴安落叶松(Larix gmelinii)根际土壤真菌群落研究
2021-12-25邱钰明王中斌汪海霞曲来叶
杨 寅,邱钰明,王中斌,汪海霞,曲来叶,*
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085 2 中国科学院大学,北京 100049 3 北京林业大学,北京 100083 4 天津市林业调查规划设计院,天津 300112
林火是森林中重要的扰动因子,其对森林的影响程度首要取决于火烧严重程度[1],重度火烧会显著地改变森林生态系统的结构以及演替过程[2]。土壤微生物在森林生态系统的物质循环和能量流动过程中发挥着重要作用[3],真菌作为土壤微生物的重要组成成分,对火烧干扰的响应迅速[4],对土壤理化性质的改变十分敏感,真菌群落的组成与功能是火烧干扰后土壤恢复的重要生物指标[5]。真菌能与植物根系形成菌根真菌促进植物氮、磷元素的吸收[6],又可作为分解者加快土壤中植物可吸收养分的释放[7],也可成为病原体阻碍植物的生长发育[8]。同时,植物的养分运输也直接影响菌根真菌,植物凋落物的质量和数量也会对腐生真菌产生间接影响。因此,当植物受到火烧扰动时,真菌群落的组成与功能也会受到相应的影响[9],植物与真菌之间的反馈使得二者的变化存在显著的相关性[10],在火烧程度严重的情况下这种相关性会长期存在[11]。此外,植物主要通过根际分泌物影响真菌群落[12],根际与非根际土壤真菌群落存在显著差异。因此,基于根际土壤的研究将更为准确地反映火烧干扰后植物与真菌群落变化的相关性。
大兴安岭林区是森林火灾的频发区,也是我国森林火烧程度最严重的林区之一。兴安落叶松(Larixgmelinii)为松科(Pinaceae)落叶松属(Larix)乔木,其耐寒耐瘠薄,木材蓄积丰富,是该林区荒山造林和森林更新的主要树种[13]。重度火烧迹地兴安落叶松的生长和恢复是大兴安岭森林生态系统恢复的关键,对我国林业生产具有重要战略意义[14]。国内对火烧迹地兴安落叶松土壤真菌群落具有一定的研究基础,张敏等通过稀释平板法研究了火烧迹地兴安落叶松土壤真菌数量的变化趋势[15],彭瑶等利用磷脂脂肪酸方法探讨了火烧迹地兴安落叶松土壤真菌与细菌相对含量的变化特征[16];张虹通过变性梯度凝胶电泳法研究发现不同火烧程度和不同恢复年限的兴安落叶松林土壤真菌群落结构均存在显著差异[17]。但是,以上研究均以火烧迹地的非根际土壤为研究对象,且对于重度火烧后兴安落叶松生长状态与根际土壤真菌群落组成与功能相关性的研究尚未涉及。近年来,高通量测序技术的发展以及FUNGuild工具[18- 19]的开发极大地推进了真菌群落组成与功能的深入研究,为重度火烧迹地兴安落叶松生长状态与根际土壤真菌群落组成与功能相关性的研究奠定了基础。
综上所述,本研究在内蒙古根河大兴安岭林区重度火烧迹地中选取两种火烧后生长状态差异显著的兴安落叶松(火烧枯立木和火烧存活木)为研究对象,以未火烧样地中的兴安落叶松作为对照(未火烧对照木),利用磷脂脂肪酸方法分析了根际土壤真菌群落生物量的变化特征,并通过高通量测序手段探讨了根际土壤真菌群落组成与功能变化,探究重度火烧后兴安落叶松生长状态与根际土壤真菌群落组成与功能的相关性。旨在解决以下问题:重度火烧迹地兴安落叶松的生长状态如何影响根际土壤真菌群落组成与功能?本研究将为重度火烧迹地兴安落叶松的恢复与管理提供科学参考和决策依据。
1 研究方法
1.1 研究区概况
本研究于2018年7月选取内蒙古根河林业区2015年重度火烧(烧死木占蓄积量60%以上)的兴安落叶松林(火烧后自然恢复,无人为干预),以及附近未火烧的兴安落叶松林(对照)为研究区开展调查。研究区地理坐标为122°25′56.46″—122°26′3.68″E,50°59′22.63″—50°59′26.71″N,海拔913m。该地区年降水量450—500mm,年均温-5.4℃。土壤以棕色针叶林土和暗棕壤为主,森林覆盖率高达75%,主要树种为兴安落叶松(Larixgmelinii)、白桦(Betulaplatyphylla)、樟子松(Pinussylvestris)等。
1.2 样地选择与采样
初步调查发现重度火烧迹地内兴安落叶松的生长状态由着火点向外大致以同心圆形状呈阶梯分布。将其中树皮脱落超过50%且树枝没有当年生针叶的兴安落叶松标记为火烧枯立木(Burned dry wood, BDW);将树皮有轻微脱落(<20%)但树枝有当年生针叶的兴安落叶松标记为火烧存活木(Burned survival wood, BSW)。在重度火烧迹地内,根据兴安落叶松的生长状态,设置两个20m×20m的样方:第一个样方距离火场中心较近(100m以内),样方内兴安落叶松多为火烧枯立木;第二个样方距离火场中心较远(100m以外),样方内兴安落叶松多为火烧存活木。同时,在未火烧的对照样地中设置20m×20m的对照样方。在第一个样方内选取火烧枯立木9棵,在第二个样方内选取火烧存活木9棵,在对照样方内选取9棵生长状态良好的未火烧对照木。综上,共选取27棵兴安落叶松,其胸径基本一致。
对27棵兴安落叶松的根际土壤(0—10cm)进行采集。采样前,先扫去土壤表面的枯枝落叶等杂物,用铁锹挖土找到兴安落叶松的树根,用手抖落根际土。在每棵兴安落叶松根部四个方向取土,并混合为一个土样,作为该兴安落叶松的根际土壤样品。新鲜土壤过2mm筛后放于4℃冰盒里短暂保存,带回实验室。取部分新鲜土样保存于-20℃冰箱中用于磷脂脂肪酸(Phospholipid fatty acid, PLFA)、土壤DNA的提取,另一部分土样用于根际土壤理化性质的测定。
1.3 根际土壤理化性质与磷脂脂肪酸测定
土壤理化性质测定方法[20]:土壤含水量采用铝盒烘干法测定,土壤pH值采用酸度计(土水比1∶5)测定,土壤总碳、总氮含量采用元素分析仪测定,土壤总磷、总钾含量采用微波消解ICP-OES法测定,土壤速效磷含量采用碳酸氢钠法提取并用紫外分光光度计测定,土壤速效氮含量采用碱解扩散法测定。
土壤磷脂脂肪酸测定方法[21]:(1)提取:取4g冻干土(过100目筛)加3.6mL磷酸缓冲液、8mL甲醇、4mL氯仿,超声波10min,震荡1—2h,2500rpm/min离心10min。上清液加4mL氯仿、3.6mL磷酸缓冲液,混匀,分层过夜(避光),收集下层氯仿相,氮气吹干。(2)分离:用氯仿分两次洗涤转移吹干的样品,用氯仿润湿硅胶柱。加15mL丙酮过硅胶柱、加10mL甲醇(色谱纯)过硅胶柱,收集甲醇相,氮气吹干。(3)甲酯化:用甲醇-甲苯(1∶1,v/v)溶液溶解吹干的脂类物质,加1mL氢氧化钾(甲醇作溶剂,0.2mol/L),混匀,37℃温水浴15min。冷却至室温,加2mL氯仿-正己烷(1∶4,v/v)、0.3mL醋酸(1mol/L)、2mL超纯水,混匀,2000rpm/min离心5min,收集上层正己烷相。加2mL氯仿-正己烷(1∶4,v/v)重复提取一次,合并两次提取的正己烷相,氮气吹干,-20℃保存。(4)上机测定:用19C做内标,通过气相色谱仪得到磷脂脂肪酸特征图谱,根据内标计算含量,单位用nmol/g表示。根据已发表文献中的磷脂脂肪酸标记物[22- 23],对真菌(18:2ω6c, 18:1ω9c)的磷脂脂肪酸含量进行表征。
1.4 根际土壤DNA提取、真菌ITS基因扩增及测序
对兴安落叶松根际土壤进行冷冻干燥和过100目筛处理,采用Qiagen土壤DNA提取试剂盒(DNeasy Power Soil Kit)对兴安落叶松根际土壤进行DNA抽提。将提取好的DNA送至上海美吉生物公司进行真菌片段PCR扩增和纯化,真菌ITS引物为[24]:ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA- 3)和ITS2R(5′-GCTGCGTTCTTCATCGATGC- 3′)。PCR产物大小在300—500bp之间,Illumina测序。
1.5 生物信息学分析及数据处理
采用Qiime2软件(qiime2 2019.1)对测序原始数据进行生物信息分析:利用DADA2对下机数据进行质控[25],生成OTU表和代表性序列,过滤掉在Illumina测序数据中用于提高扩增子测序质量的任何phiX序列,同时过滤嵌合体序列。将测序得到的序列进行随机抽样标准化,不同处理样品的稀释曲线均趋于平缓,OTU数目趋于饱和,能够准确反应真菌群落特征。将重抽样标准化后的OTU表转化为相对丰度,从Unite中下载真菌数据库sh_refs_qiime_ver8_dynamic_02.02.2019用于比对,在Qiime2中训练Naive Bayes分类器,进而对各个OTU进行物种注释并分级汇总可视化。利用Qiime2计算α多样性指数[26](Chao1指数、Pielou均匀度指数、香农多样性指数)。使用R语言vegan包进行基于Weighted-Unifrac距离的主坐标分析(Principal coordinates analysis,PCoA)比较各样地真菌群落的β多样性并进行群落相似性分析(Analysis of similarities, ANOSIM)[27]。通过LEfSe(LDA Effect Size)分析寻找组间差异物种(LDA阈值为4.0)[28],使用FUNGuild对真菌群落进行营养方式(Trophic mode)与功能群(Guild)划分[18- 19]。使用SPSS24.0对根际土壤理化性质、磷脂脂肪酸、真菌群落多样性指数、组成与功能进行单因素方差分析,用LSD方法比较不同组之间的差异。使用R语言vegan包进行冗余分析(Redundancy analysis,RDA)探讨影响根际土壤真菌菌门相对丰度的主要环境因子[27],通过Pearson相关性分析真菌菌属相对丰度与土壤理化性质的相关性。所有制图通过Origin 9.1、R 3.5.1版本完成。
2 结果与分析
2.1 火烧后不同生长状态的兴安落叶松根际土壤理化性质与真菌磷脂脂肪酸含量
根际土壤理化性质如表1所示,火烧枯立木与火烧存活木根际土壤总磷含量显著高于未火烧对照木(P<0.05),但火烧枯立木与火烧存活木根际土壤总磷含量差异没有达到显著水平。火烧枯立木根际土壤速效磷含量显著高于未火烧对照木(P<0.05),总钾含量显著低于未火烧对照木(P<0.05),而火烧存活木速效磷、总钾含量与未火烧对照木均不存在显著差异。此外,火烧枯立木与火烧存活木的土壤pH、含水量、总碳、总氮、速效氮含量与未火烧对照木均不存在显著差异。

表1 火烧后不同生长状态的兴安落叶松根际土壤理化性质
根际土壤真菌磷脂脂肪酸含量如图1所示。相较于未火烧对照木,火烧枯立木与火烧存活木根际土壤真菌磷脂脂肪酸含量显著降低(P<0.05),且火烧枯立木根际土壤真菌磷脂脂肪酸含量显著低于火烧存活木(P<0.05)。

图1 火烧后不同生长状态的兴安落叶松根际土壤真菌磷脂脂肪酸含量Fig.1 Fungi PLFA contents in rhizosphere soil of Larix gmelinii in different growth status after fire字母不同代表处理间差异显著(P<0.05)
2.2 火烧后不同生长状态的兴安落叶松根际土壤真菌群落多样性分析
根际土壤真菌群落的α多样性如表2所示。相较于未火烧对照木,火烧枯立木与火烧存活木根际土壤真菌群落Chao1指数显著降低(P<0.05),但Pielou均匀度没有发生显著变化。火烧存活木的香农多样性指数显著低于未火烧对照木(P<0.05),而火烧枯立木的香农多样性指数没有发生显著变化。
真菌群落β多样性的主坐标分析结果如图2所示。前两个排序轴的解释率分别为38.28%与17.92%。其中,未火烧对照木聚集在第一排序轴的正半轴,而火烧枯立木与火烧存活木聚集在第一排序轴的负半轴。同时,火烧枯立木聚集在第二排序轴的负半轴,而火烧存活木在第二排序轴的正负半轴均有聚集。真菌群落的相似性分析(ANOSIM)结果显示,火烧枯立木、火烧存活木、未火烧对照木根际土壤真菌群落存在极显著差异(P<0.001)。

图2 真菌群落β多样性主坐标分析以及群落相似性分析 Fig.2 PCoA of beta diversity and ANOSIM of rhizosphere soil fungal community
2.3 火烧后不同生长状态的兴安落叶松根际土壤真菌群落组成及功能差异
从重度火烧后不同生长状态的兴安落叶松根际土壤测序样品中共划分出2230个真菌OTU,隶属8门21纲50目87科146属。真菌群落门水平的相对丰度如表3所示,未火烧对照木根际土壤中担子菌门(Basidiomycota)为优势菌门(相对丰度60%)。相较于未火烧对照木,火烧枯立木与火烧存活木根际土壤中担子菌门(Basidiomycota)相对丰度显著降低(P<0.05),优势菌门转变为子囊菌门(Ascomycota),相对丰度显著升高(P<0.05)。火烧枯立木与火烧存活木各真菌菌门的相对丰度不存在显著差异。
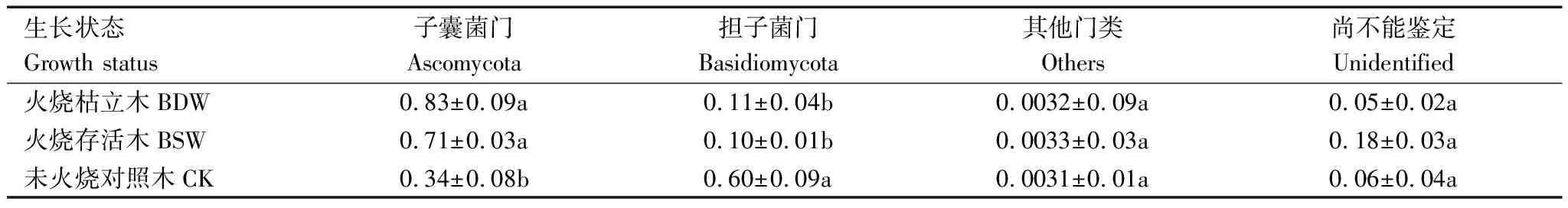
表3 火烧后不同生长状态的兴安落叶松门分类水平下根际土壤真菌群落组成
表2 火烧后不同生长状态的兴安落叶松根际土壤真菌群落α多样性

Table 2 Alpha diversity of fungal communities in rhizosphere soil of Larix gmelinii in different growth status after fire
LEfSe分析结果及系统发育树如图3所示,在LDA阈值为4.0的显著差异条件下,火烧枯立木、火烧存活木、未火烧对照木从纲到属存在显著差异的真菌类群有21个,其中属分类水平上有10个。将存在显著差异的真菌菌属进行营养方式(Trophic mode)与功能群(Guild)划分,结果如表4所示。未火烧对照木根际土壤中真菌主要营养方式为共生型,担子菌门(Basidiomycota)的蜡伞属(Hygrophorus)、红菇属(Russula)、丝膜菌属(Cortinarius)均为外生菌根真菌,相对丰度显著高于火烧枯立木与火烧存活木(P<0.05)。火烧枯立木根际土壤中真菌主要营养方式为共生型以及病原-腐生-共生复合型。其中,子囊菌门(Ascomycota)的Leptodontidium属、Phialocephala属为内生真菌,Coniochaeta属为子囊菌门(Ascomycota)的动物病原-粪腐生-内生复合功能真菌,这三个菌属的相对丰度均显著高于火烧存活木与未火烧对照木(P<0.05)。火烧存活木根际土壤中真菌主要营养方式为共生型以及腐生-共生复合营养方式。其中,子囊菌门(Ascomycota)的长毛盘菌属(Trichophaea)为粪腐生-外生菌根复合功能真菌,Calyptrozyma属为木质腐生-外生菌根复合功能真菌,这两个菌属的相对丰度显著高于火烧枯立木与未火烧对照木(P<0.05)。

表4 火烧后不同生长状态的兴安落叶松根际土壤真菌菌属功能群

图3 火烧后不同生长状态的兴安落叶松根际土壤真菌线性判别分析结果与系统发育树Fig.3 LEfSe analysis and cladogram of rhizosphere soil fungi of Larix gmelinii in different growth status after fire图中每一个节点代表一个真菌类群,节点的直径代表相对丰度的大小,组间相对丰度差异不显著的真菌类群用黄色节点标注,差异显著的真菌类群跟随组别进行着色;a:Calyptrozyma:Calyptrozyma属;b:Leptodontidium:Leptodontidium属;c:Phialocephala:Phialocephala属;d:Vibrisseaceae:Vibrisseaceae科;e:Trichophaea:长毛盘菌属;f:Coniochaeta:锥毛壳属;g:Coniochaetaceae:锥毛壳科;h:Coniochaetales:锥毛壳目;i:Trichoderma:木霉属;j:Hygrophoraceae:蜡伞科;k:Sordariaceae:粪壳菌科;l:Cortinarius:丝膜菌属;m:Cortinariaceae:丝膜菌科;n:Hygrophorus:蜡伞属o:Hypocreaceae:肉座菌科;p:Inocybe:丝伞盖属;q:Inocybaceae:丝伞盖科;r:Russula:红菇属;s:Russulaceae:红菇科;t:Russulales:红菇目;u:Thelephorales:革菌目
2.4 影响兴安落叶松根际土壤真菌群落的环境因子
真菌群落门水平组成与根际土壤理化性质的冗余分析结果如图4所示(仅展示相关性显著的根际土壤理化性质),前两个排序轴的解释率分别为44.56%与1.60%。根际土壤总磷与总钾含量是影响真菌群落门水平组成的主要环境因子。其中,根际土壤总磷含量与子囊菌门(Ascomycota)相对丰度显著正相关(P<0.05),与担子菌门(Basidiomycota)相对丰度显著负相关(P<0.05)。根际土壤总钾含量与子囊菌门(Ascomycota)相对丰度显著负相关(P<0.05),与担子菌门(Basidiomycota)相对丰度显著正相关(P<0.05)。
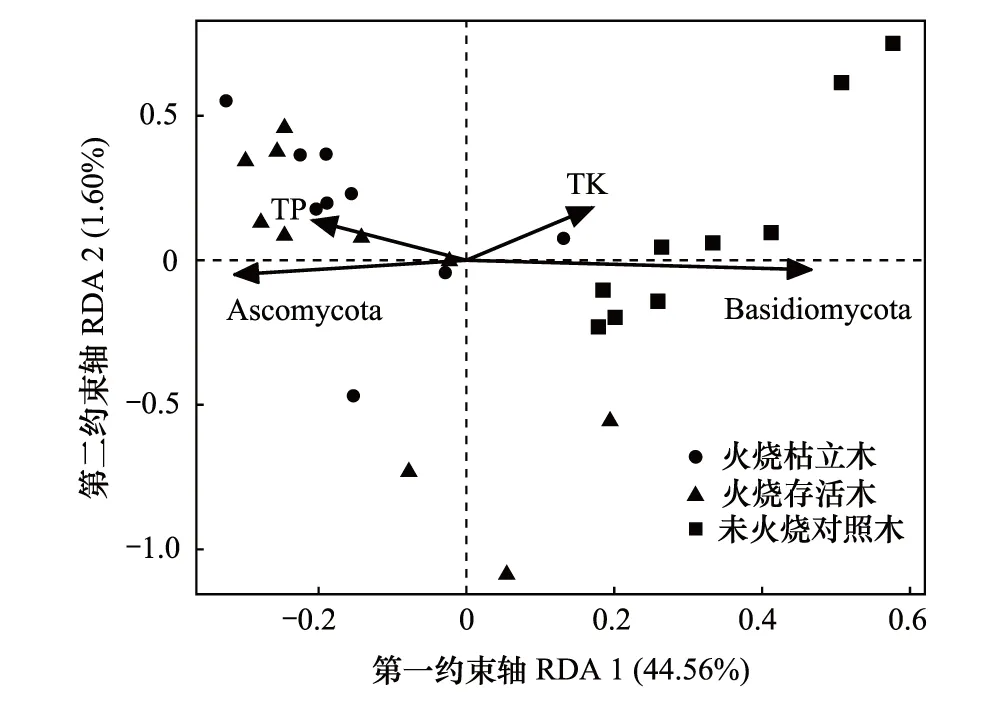
图4 真菌群落组成与土壤理化性质的冗余分析 Fig.4 RDA of fungal communities composition and physicochemical properties of rhizosphere soil
为探究造成火烧枯立木与火烧存活木根际土壤真菌群落差异的因素,对子囊菌门内相对丰度较高且存在显著差异的真菌菌属与根际土壤理化性质进行Pearson相关分析,结果如表5所示。Leptodontidium属相对丰度与总磷含量显著正相关(P<0.05),Phialocephala属相对丰度与总碳、总氮含量显著正相关(P<0.05),Calyptrozyma属相对丰度与总碳、总磷含量显著正相关(P<0.05),Trichophaea属、Coniochaeta属相对丰度与根际土壤理化性质不存在显著相关性。

表5 真菌菌属相对丰度与土壤理化性质的相关系数
3 讨论与结论
3.1 真菌群落磷脂脂肪酸含量与多样性
磷脂脂肪酸含量是反映土壤微生物量大小的重要指标[29]。火烧迹地真菌生物量变化的原因可归结为火烧过程中土壤升温对真菌的致死作用[30]、火烧后土壤水分养分条件的破坏[31]以及共生植物缺失。本研究中火烧枯立木与火烧存活木根际土壤真菌磷脂脂肪酸含量显著低于未火烧对照木,重度火烧显著降低了兴安落叶松根际土壤真菌群落生物量,与已有重度火烧迹地中真菌生物量变化的研究结果一致[32]。火烧枯立木真菌磷脂脂肪酸含量显著低于火烧存活木,这可能有以下两方面原因:一是因为重度火烧迹地中土壤升温不均匀导致火烧枯立木与火烧存活木根际土壤真菌生物量损失程度存在差异,二是因为真菌生物量受到兴安落叶松生长状态的影响,火烧存活木生长状态较好,促进了火烧后根际土壤真菌生物量的恢复。Chao1指数是反映物种丰富度的指标[33],Pielou均匀度指数反映物种分布均匀度[34]。结果显示,火烧枯立木与火烧存活木的Chao1指数显著低于未火烧对照木,但Pielou均匀度指数与未火烧对照木不存在显著差异,这说明重度火烧降低了兴安落叶松根际土壤真菌群落的物种丰富度但没有对物种分布均匀度产生影响。
3.2 真菌群落组成和功能变化特征以及环境因子影响机制
本研究中兴安落叶松根际土壤真菌门类以子囊菌门(Ascomycota)和担子菌门(Basidiomycota)为主,这与已有大兴安岭针叶林土壤真菌群落的研究结果一致[35]。其中,担子菌门(Basidiomycota)常与植物共生形成菌根[36],促进植物的养分吸收与生长发育。未火烧对照木根际土壤水分养分条件良好,故其根际土壤真菌群落中担子菌门(Basidiomycota)为优势菌门。子囊菌门(Ascomycota)可以产生胞外酶[37],更好地降解木质素、纤维素等难分解的物质,对植物残茬和碎屑十分敏感[38- 39],重度火烧后地表枯枝落叶与灰烬大量增加,所以子囊菌门(Ascomycota)在火烧枯立木与火烧存活木根际土壤真菌群落中占据了优势。冗余分析结果表明,重度火烧后根际土壤总磷、总钾含量是影响真菌门类的主要环境因子,有研究表明在富含磷元素的土壤中担子菌门相对丰度较低[40- 41],与本研究结果一致。真菌功能群分析结果表明,未火烧对照木根际土壤中担子菌门(Basidiomycota)的优势菌属多为外生菌根真菌,这类真菌被认为是对火烧敏感的真菌类群[42],且外生菌根真菌具有宿主植物专一性,火烧后外生菌根真菌群落的变化会受到恢复过程中定植植物的影响[43]。火烧枯立木与火烧存活木根际土壤中外生菌根真菌相对丰度显著减少,这说明兴安落叶松的生长状态以及土壤环境尚未恢复到适宜外生菌根真菌繁殖的水平。外生菌根真菌是森林生态系统中重要的菌类,它可以扩大兴安落叶松根系吸收营养的面积、增强植物的抗逆性、促进植物生长[44],可考虑在重度火烧迹地兴安落叶松的恢复过程中接种外生菌根真菌来促进恢复。
火烧枯立木与火烧存活木根际土壤真菌群落中子囊菌门(Ascomycota)的优势菌属相对丰度及其所属功能群存在显著差异:火烧枯立木根际土壤中相对丰度显著增加的Leptodontidium属、Phialocephala属为内生真菌。其中,Phialocephala属为深色有隔内生真菌,可以在宿主根系定植并抑制致病菌的生长[45],同时增加植物生物量[46],而Leptodontidium属的生长繁殖特性与Phialocephala属十分相似[47- 48],Analie等通过盆栽实验研究发现Leptodontidium属的寄主植物十分广泛,其与Phialocephala属的共存可以显著增加植物茎重[49]。火烧存活木根际土壤中相对丰度显著增加的长毛盘菌属(Trichophaea)与Calyptrozyma属为腐生-共生复合型真菌。其中,长毛盘菌属(Trichophaea)属于盘菌目(Pezizales),这一真菌菌目具备一定的适应火烧的特性,如耐高温、火烧后不久会产生大量耐热孢子等[50],在轻中度火烧迹地中会作为火烧适应型真菌来替代减少的火烧敏感型真菌。火烧存活木根际土壤中相对丰度增加的菌属多为腐生-共生复合营养型真菌,真菌的营养方式与寄主植物生长状态以及土壤环境密切相关,真菌会改变自身的营养方式以维持正常的生理活动[51],当植物健康生长时为共生型真菌,当植物衰老或死亡时可转变为腐生真菌,这些真菌为应对不同的生存条件而采取了不同的营养策略[52]。由此可见,火烧枯立木与火烧存活木真菌群落的恢复策略存在差异,火烧枯立木根际土壤真菌群落通过Leptodontidium属、Phialocephala属这两种内生菌根真菌的共同作用来促进兴安落叶松根系与生物量的恢复,而火烧存活木根际土壤真菌群落对火烧的适应性显著增加,同时通过与兴安落叶松的腐生与共生关系促进其根际土壤物质循环与能量流动。此外,不同于火烧枯立木,火烧存活木根际土壤中病原型真菌相对丰度没有较未火烧对照木显著增加,这说明兴安落叶松良好的生长存活状态对其根际土壤真菌群落中病原型真菌有一定的抑制作用[53]。相关性分析表明,火烧枯立木与火烧存活木根际土壤中存在显著差异的真菌菌属中,Leptodontidium属、Phialocephala属、Calyptrozyma属相对丰度与根际土壤理化性质(总碳、总氮、总磷含量)存在显著相关性,而Trichophaea属、Coniochaeta属相对丰度与根际土壤理化性质不存在显著相关性,这说明除环境因子外,火烧后兴安落叶松的不同生长状态也对根际土壤真菌群落属水平上的组成及功能产生了不同的影响,造成了火烧枯立木与火烧存活木根际土壤真菌群落的差异。
综上所述,重度火烧后兴安落叶松根际土壤真菌群落生物量与多样性显著降低,火烧枯立木与火烧存活木的根际土壤真菌群落组成及功能存在显著差异。根际土壤总碳、总氮、总磷、总钾含量是影响真菌群落的主要环境因子。根际土壤真菌群落的变化特征对于兴安落叶松的恢复具有重要的指示意义,应在火烧恢复中引起重视。
