四川丘陵区水库浮游植物群落结构与蓝藻水华风险
——基于优势种生态位与种间联结研究
2021-12-24岳兴建代丽娜韩文文王永明
陶 敏,岳兴建,岳 珊,代丽娜,韩文文,王永明,刘 果,李 斌,*
1. 内江师范学院,生命科学学院, 内江 641112 2. 长江上游鱼类资源保护与利用四川省重点实验室, 内江 641112
物种生态位理论与种间联结性理论能解释自然群落中不同物种共存与竞争机制,对研究群落结构功能、群落内种间关系、群落演替、种群进化等方面具有指导意义[1-2]。生态位测度包括生态位宽度和生态位重叠两个指标,其中生态位宽度为反映物种所能利用的各种资源的一个综合指标[3],生态位重叠则反映两个物种对同一资源的利用效率及共享程度[4],二者被认为是物种多样性及群落结构的决定因素,也反映了其所在群落的稳定性[5]。种间联结体现了物种与物种之间在分布上的关联,是群落重要的结构特征与数量特征,也是群落结构形成与发生演化的基础[6]。浮游植物是水域生态系统中的重要初级生产者,其群落结构与动态直接影响并反映着生态系统的稳定性,关系着水体的健康与功能。对浮游植物优势种生态位与种间联结性开展相关研究,不仅可以帮助了解群落内各种群对资源的利用情况,而且有助于掌握种群的竞争机制和竞争规律,揭示浮游植物适应性特征与环境变化之间的关系[7- 9],预测环境变化乃至全球环境变化对群落结构的影响[10- 12]。目前关于浮游植物生态位相关研究较少,主要集中于海洋[7-8,11- 15]。另外在湖泊、河流等淡水水域中也有少量相关研究[2,9,16- 20]。
四川丘陵地区降水分布不均[21],资源性缺水及工程性缺水严重[22],故而水库在该地区人民生产生活中扮演着极其重要的角色。截至2011年底,四川省已建总库容10万m3及以上水库工程共8072座,其中大型水库51座,中型水库219座,小型水库7878座[23];多数水库承担着当地居民饮用水源和农业灌溉的重任[23]。与大型水库不同,中、小型水库由于水体容量小、自净力较弱、受工农业以及人类生活影响较大,水质情况往往不容乐观,富营养化与藻类水华爆发的风险较高。因此,本文选择四川丘陵区典型中、小型水库为研究对象,通过生态位理论和种间联结法,分析浮游植物群落中优势种利用资源的能力和在环境中的适应能力,探寻优势种间相互作用的生态关系与发展方向,并据此预测该类水库中发生蓝藻水华的可能性,为该地区水库水源保护与生态修复提供科学依据。
1 材料与方法
1.1 研究水库概况与采样点设置
研究的10座代表性水库位于四川省东南丘陵地区。该区域地处长江上游,坐标104°15′—105°26′E、29°11′—30°2′N,地势呈不规则状展布,海拔274—900m,属台状低山丘陵地形。气候类型为亚热带湿润季风气候,冬暖夏热,年均温15—28℃,年降雨量约1000 mm,60%分布在夏季[24-25]。
10座水库(图1)中有中型水库5座、小型水库5座,大坝坐标范围29°20′02″N—29°54′51″N,104°37′12″—105°15′42″E,总库容1.7×105—7.58×107m3(表1)。按面积与形状,在每个水库中设置1—5个采样点,其中库容大于500×105m3的设置5个采样点,100×105—500×105m3的设置4个,10×105—100×105m3的设置3个,库容最小的五星水库设置1个样点,于2016年8月(夏)、2016年11月(秋),2017年2月(冬)和2017年5月(春)分季节开展采样工作。

图1 研究水库及采样点分布图Fig.1 Distribution of reservoirs and sampling sites surveyedHLK: 葫芦口水库; GYH: 古宇湖; LJ: 龙江水库; SL: 松林水库; HH: 黄河水库; BY: 八一水库; CZG: 菜子沟水库; YJT: 严家滩水库; WGS: 乌龟山水库; WX: 五星水库; 1—5: 各水库采样点编号

表1 研究水库概况
1.2 样品采集、处理与测定
采样点透明度(Secchi Depth, SD)、水温(Water temperature, WT)、pH等理化指标使用塞氏盘、便携式pH计(METTLER TOLEDO Seven2Go)等设备进行原位测定。总氮(Total nitrogen, TN)、总磷(Total phosphorus, TP)样品采集与测定参考相关文献[26]在实验室中进行。浮游植物定量以5L有机玻璃定深采水器采集,将1L表层(水下0.5m)水样装入1L样品瓶并立即加入鲁哥试剂现场固定。浮游植物定性样品用25号浮游生物网(0.064mm)在水面下作“∞”字形捞取并过滤收集,加入4%甲醛现场固定。回到实验室后将浮游植物定量样品在1L规格浮游生物沉淀器中避光静置沉降48h, 以虹吸法去除上层清液后, 浓缩定容至50mL,加入数滴甲醛溶液保存。种类鉴定参考胡鸿钧等[27],计数方法参照章宗涉等[28]。
1.3 数据处理与分析
1.3.1优势种确定
浮游植物优势种以Mcnaughton优势度指数(Y)值进行鉴别[2]:
Y=Pi×fi
式中,Pi为第i种浮游植物丰度与样品中浮游植物总丰度的比值(ni/N),fi为第i种在各采样点出现的频度。Y≥0.02的物种为优势种。
1.3.2生态位宽度
采用修正后的Levins指数计算优势种生态位宽度[2]:
式中,Bi为第i种的生态位宽度,Pij=nij/Ni,为第i种在第j个样点出现的个体数占该种所有个体数的比例,r为样点数。
1.3.3生态位重叠与生态响应速率
采用Petraitis指数计算优势种之间的生态位重叠[2]:
SOik=eEik
式中,SO为成对种间生态位特定重叠,k为不同于i的另一浮游植物物种,SOij表明物种对资源的利用程度,Ri为生态响应速率。
1.3.4物种间总体关联性
物种间总体关联性测定采用Schluter的方差比率法(VR)[2],VR计算分为以下几步:
式中,S为浮游植物优势种的总种类数;Pi=ni/N,其中N表示样方总数,ni表示物种i出现的样方数;Tj表示样方j内出现的浮游植物优势种种数;t表示样方中浮游植物优势种物种数的平均数t=(T1+T2+…+Tn)/N;δT2为所有采样点物种数的方差;ST2表示总种数出现频度的方差。
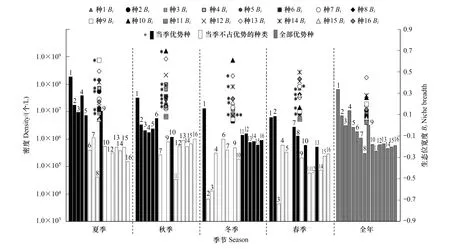
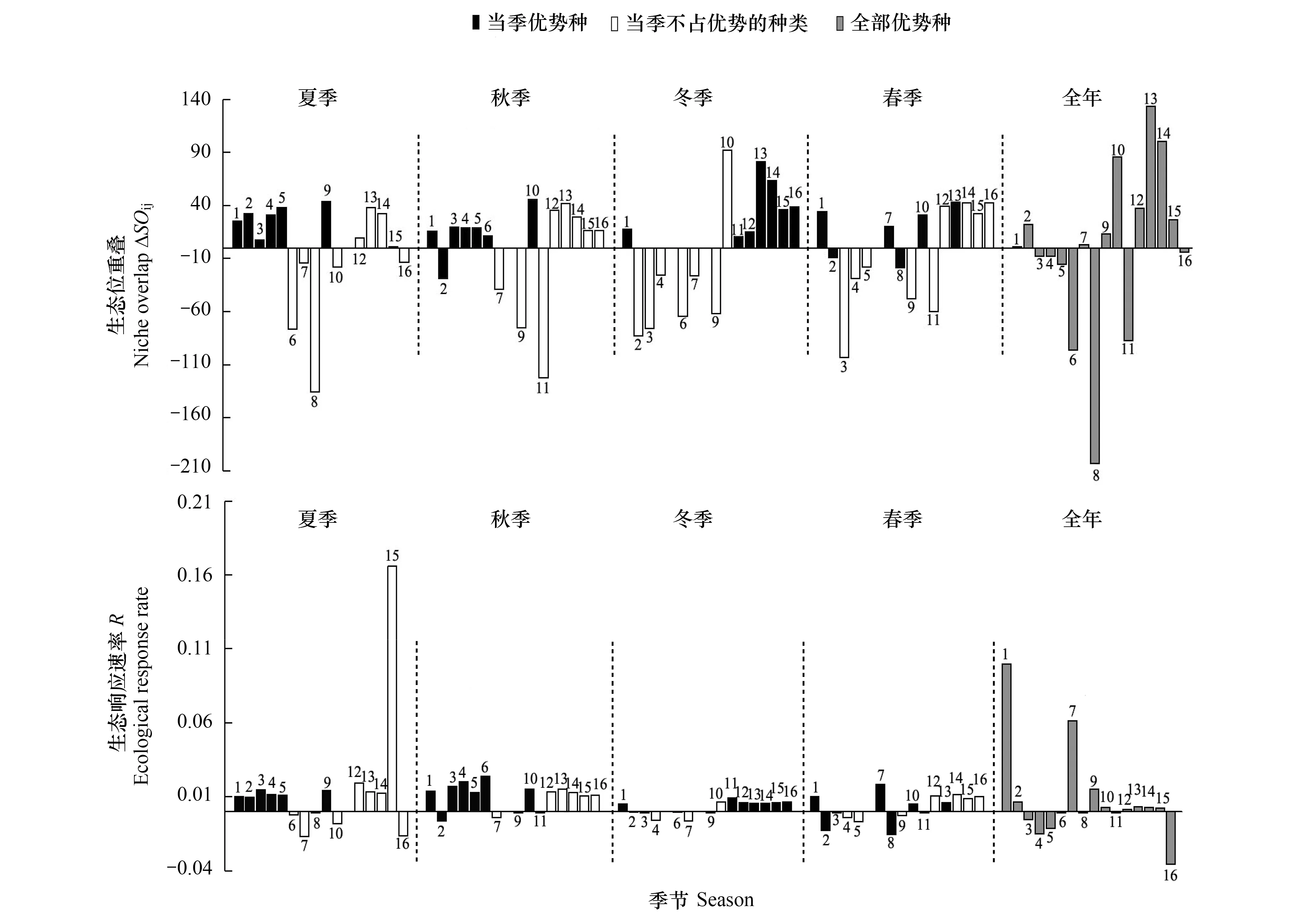
当VR=1时,符合所有物种无关联的零假设;当VR<1 时,物种间总体为负协变关系;当VR>1 时,物种间总体为正协变关系。利用统计量W=VR×N来检验VR值偏离1的显著程度,在物种间总体无关联的情况下,χ20.95N 1.3.5优势种种对间关联 根据物种在样方内是否存在,将全年207个样点× 16个优势种的数量数据矩阵转化为 0,1 形式的二元数据矩阵[29]。基于2×2列联表,采用 Yates连续校正法, 进行χ2卡方检验[30], 其计算公式如下: χ2<3.84(P>0.05),表示物种间无显著关联性;3.841≤χ2≤6.635(0.01≤P≤0.05)表示物种间关联显著;χ2>6.635(P<0.01)则表示物种间关联性极显著。 调查期间共检出浮游植物9门104属188种,其中绿藻门出现的种类最多为48属84种,占44.7%,其次为硅藻门(15属35种,占18.6%)和蓝藻门(19属34种,占18.1%)。其余门类种类数均较少,包括裸藻门5属12种,金藻门8属10种,甲藻门4属7种,隐藻门2属4种,种类数最少的黄藻门和定鞭藻门,均只出现1属1种,仅占0.5%。从种类数来看,研究水库浮游植物群落组成与长江上游其他河流型水库近似,以绿藻+硅藻+蓝藻为主[31- 36],绿藻、蓝藻等静水种类的增多,暗示着水体的富营养化[33,37]。 从现存量季节分布来看(图2),夏季浮游植物密度达到最大,为(2.85±3.54)×108个/L,其后依次为秋季(6.04±8.52)×107个/L、冬季(2.38±4.92)×107个/L和春季(2.11±2.84)×107个/L。蓝藻在四季均占据优势地位,其中占比最高的为夏季(2.78±3.53)×108个/L,占97.6%;其次为秋季(4.95±8.19)×107个/L(81.9%)与春季(1.81±2.67)×107个/L(85.8%),冬季最低(1.56±4.65)×107个/L(64.1%)。从现存量空间分布上看(图2),10个水库中浮游植物总密度超过108个/L的有4个,超过107个/L的有5个;其中最高的为WGS,年均值为(2.39±5.18)×108个/L,最低为HLK,年均值为(5.11±6.51)×106个/L。各水库均以蓝藻占优势,所占比例从55.3%(HLK)到96.7%(WGS)不等,其中蓝藻所占比例超过95%的有3个(WGS、SL、LJ),其他藻类仅有绿藻在HLK中占一定优势(38.1%)。与长江上游大型水库如紫坪铺、长寿湖、升钟湖、二滩水库等相比[31- 35],本研究中的中小型水库浮游植物丰度高出2—3个数量级,按照相关评价标准[38],夏季已达到极富营养水平,秋季为富营养水平,冬春两季为中营养水平,富营养化比大型水库更为明显。尽管蓝藻种类数明显少于绿藻和硅藻,但其丰度占据了绝对优势,这一现象与长江上游流域部分水库类似[36,39],爆发蓝藻水华的风险较高。 图2 研究水库浮游植物密度时空分布Fig.2 The temporal and spatial distribution of phytoplankton abundance 四季共出现4门16个优势种(Y≥0.02),其中蓝藻9种、绿藻4种、硅藻1种、隐藻2种;以全年数据进行分析,结果显示优势种仅3种,均为蓝藻(表2)。与蓝藻(Y值跨度0.020—0.435,均值(0.124±0.136))相比,其他藻类(Y值跨度0.020—0.056,均值(0.031±0.011))优势度指数均不高(P=0.013,独立样本t检验)。优势种组成说明了研究水库污染现状不容乐观:种类数最多的蓝藻是最典型的富营养水体优势类群[37],其他种类则多为中污带指示生物或耐有机污染种类[40]。 表2 研究水库优势种优势度指数 尽管优势种种类数较多,其优势度和密度在不同季节存在差异。除冬季以绿藻和隐藻为主要优势藻种外,其余3个季节均以蓝藻占多数,尤其是丝状蓝藻(表2)。湖泊假鱼腥藻(Pseudanabaenalinmnetica)在四季都具有最高的优势度,其年均密度((6.26±18.73)×107个/L)和最高密度(1.84×108个/L)也均是最高,成为绝对优势种。拉氏拟柱孢藻(Cylindrospermopsisraciborskii)除冬季外在其余3个季节均占优势地位,尤其在春季其密度为16种中最高((6.69±19.28)×106个/L)。表2显示了藻类在不同季节中的优势分布情况,大部分种类(9种)仅在某一季节成为优势种属(表2)。在三峡库区开展的研究也发现,丝状蓝藻的优势普遍存在[36,39],尤其是夏季,但冬春两季蓝藻优势度明显下降,且优势种间优势度指数差异不大[39],未形成如本研究中湖泊假鱼腥藻这样的绝对优势,可能跟三峡库区水体较大,自我调节能力较强有关。 研究水库面临着同样的问题,即丝状蓝藻的普遍优势,仅在程度上有所不同(表3):SL、LJ、HH、YJT、WGS五个水库中的优势种组成较简单,均以丝状蓝藻为主,其密度高达7.2×107—20.7×107个/L,占浮游植物总密度比例为78.9%—86.7%;其中SL、LJ和WGS三个水库假鱼腥藻优势度指数Y高达0.63—0.85,优势种组成单一,爆发丝状蓝藻水华风险较高。HLK、GYH和BY中丝状蓝藻的优势度则相对较低,密度在2.1×106—13.3×106个/L之间,占比18.2%—41.5%,优势种较多且优势度均不高,表明其群落结构相对来说较复杂也较为完整,水华风险相对较低。相关分析发现不同水库丝状蓝藻优势与营养水平无关(P>0.05),可能更多地取决于水库大小、周边生产生活干扰、灌溉与泄洪频率等因素的综合作用。 表3 各水库优势种密度/(×106个/L) Mean±SD与优势度指数Y 生态位宽度(niche breadth)是反映群落中物种所能利用的各种资源的一个综合指标[18,41]。生态位宽度大的物种利用资源的能力强,竞争力强,被称为泛化种或广布种;生态位宽度小的物种则相反,被称为特化种[42-43]。 从全年数据来看,绿藻生态位宽度稳定且较大,其中全年Bi值以小球藻(Chlorellavulgaris)为最高(图3)。四季中,夏季以蓝藻生态位宽度值较大,Bi最高的为当季优势种细浮鞘丝藻(Planktolyngbyasubtilis)。调查期间各季节理化指标均值见表4。相关分析显示细浮鞘丝藻密度与水温显著正相关(R=0.62,P<0.01),夏季高水温可能是其占据较宽生态位的主要原因[19]。其余三个季节生态位宽度最大的均为绿藻,其中秋、冬季均为二形栅藻(Scenedesmusdimorphus),春季为球衣藻(Chlamydomonasglobosa)。研究发现,冬季和春季Bi值最高的种类均非当季优势种,密度也较低(图3,表2);尽管这些藻类藻数量少,但由于其相对较宽的生态适应幅度与顽强的竞争力,保证了它们在各种水体环境中生存与发展的可能性。而四季中均占有优势的湖泊假鱼腥藻Bi在春季最大(0.345),冬季最小(0.09),则可能与水温有关。有研究发现,富营养化水体中适宜的水温(24—30℃) 是蓝藻暴发性增殖的必要条件[19,44]。春季调查期间水温(26.5±2.6)℃(表4),适宜其生长故生态位宽度最大,冬季水温最低(14.1±1.0)℃(表4)影响其增殖,故生态位宽度最小。而其密度则在后一个季节达到极值,如夏季最大(1.84±3.26)×108个/L,春季最小(6.08±8.55)×106个/L,可能是由于其种群发展存在时滞所致。 图3 优势浮游植物密度与生态位宽度(Bi)Fig.3 Abundance and niche breadth of dominant phytoplankton species 表4 研究水库环境因子季节变化(Mean±SD) 综合各种在不同季节的出现频率与生态位宽度,将研究水库的优势种分为3类: 第一类为研究水库中相对泛化的种群,包括二形栅藻、小球藻和湖泊假鱼腥藻,为全年生态位最宽或在各个季节均作为优势种出现的种类。小球藻与栅藻在较多的研究中被证实具有较宽的生态位,属于广布种[2,18]。尽管湖泊假鱼腥藻生态位宽度值并不最高(0.090—0.345),甚至冬季Bi为所有优势种中最低值(图3),但在优势度和密度方面,其季节均值和年均值均为所有藻类中最高。综合优势度、现存量、出现频率等因素,将湖泊假鱼腥藻也归为研究水库中相对泛化的类群。这三种藻类高度适应四川丘陵地区小型水库生态环境,拥有强的适应力,其变化可对水库中藻类群落的结构及水环境状况产生重要影响。 第二类为中生态位优势种,包括拉氏拟柱孢藻、微小平裂藻(Merismopediatenuissima)、拟短形颤藻(Oscillatoriasubbrevis)、中华小尖头藻(RaphidiopsisSinensia)4种蓝藻,在2—3个季节中占优势,且其优势度指数与生态位宽度指数处于中等水平,生态适应幅度小于前一类(表2,图3),而与第三类相比,这一类在研究水库中也具有较强的竞争力。 第三类为窄生态位优势种,为剩余的9种,包括4种蓝藻、2种绿藻、1种硅藻和2种隐藻,仅在某一季节占优势,对水温、营养条件等生态因子依赖性较高,竞争力较小,利用资源和适应环境的能力相对较弱。就在研究水库中的竞争力而言,相对较强的前两类的7个种中,有5种蓝藻,蓝藻的优势不言而喻。 生态位重叠(niche overlap)表示物种间对资源利用的相似性,反映不同物种对同一资源的共同利用程度,也体现了物种的空间分布情况[14,45]。生态位重叠值SOij表示i种群占用j种群资源的大小,亦即j种群被i种群侵占的资源量。一般情况下,生态位宽度与生态位重叠表现为正相关;ΔSOij的正负表明处于发展或衰退状态,ΔSOij差值大小表示发展或衰退的程度,ΔSOij为0时表明是一个中性种[2,14,18]。 本研究中,较大的生态位宽度大多伴随着较高的SOij值,符合一般规律[29]。从全年来看,16个优势种中9种处于发展状态,ΔSOij正值最大为小球藻(种14,133.6,图4),生态位重叠最大的则是拟鱼腥藻(种8,-203.4,图4)。四个季节的当季优势种ΔSOij多数大于0,处于发展状态,其中ΔSOij正值最大的是冬季的小球藻(种13,81.4),相关分析显示小球藻密度与TP显著正相关(R=0.395,P<0.01),冬季的高TP条件((0.896±0.533) mg/L,表4)适宜其生长,表现出最强的发展状态;少量当季优势种处于衰退状态,以秋季拟柱孢藻(种2,-28.6,图4)负值最大,可能与秋季氮磷比高((51.7±52.1),表4)有关,拟柱孢藻为固氮蓝藻[46],高氮磷比限制了其在秋季的发展。作为唯一的四季优势种,湖泊假鱼腥藻ΔSOij与其生态位宽度季节变化类似,春季最高(种1,34.2,图4),秋季最低(种1,15.6,图4),也是由于水温的季节变化而引起[19,44]。 与其他优势种之间的生态位重叠程度(ΔSOij绝对值)最大的种类ΔSOij均为负值(图4),说明与其他种群资源需求相似性最强的种群往往处于衰退状态,且衰退空间较大[2]。当共享资源丰富时,物种间的生态位重叠只表明种间的生态相似性;共享资源不足的情况下,生态位重叠则还暗示着二者存在竞争[47]。在研究的10个水库中,冬季物种间生态位重叠值(ΔSOij绝对值)普遍较高(图4),其可能原因:由于冬季低温不适合大部分优势藻类的发展,当资源(即水温等)不足时,一方面大部分藻类生长缓慢,密度降低,另一方面生态位重叠的物种间在环境相对适宜的点位上发生竞争,导致优势种生态位宽度降低,物种间生态位重叠加剧[13,19]。本研究数据显示,冬季的7个优势藻种,有5个种(湖泊假鱼腥藻、短小塔胞藻、小环藻Cyclotellasp.、啮噬隐藻Cryptomonaserosa、尖尾蓝隐藻Chroomonasacuta)有超过90%的数量分布在50%的采样点中,点位竞争激烈,其生态位宽度也低于其余2个物种(二形栅藻、小球藻)(图3)。 ΔSOij表征物种与他种竞争的结果,生态位宽度代表种群在群落中的生态优势程度,利用速率(R)将二者结合,可用以衡量物种对生境的生态响应,指示优势种种群发展性或衰退性强弱[2,17]。四个季节当季优势种中发展性最强的多为蓝藻(图4):夏季为微小平裂藻,秋季为大螺旋藻(Spirulinamajor),冬季为短小塔胞藻(Pyramimonasnanella),春季则是卷曲鱼腥藻(Anabaenacircinalis)。从全年数据来看,发展性最强的仍然为蓝藻,其中湖泊假鱼腥藻R值最高(0.100),其次为卷曲鱼腥藻(0.061)和细浮鞘丝藻(0.015),而尖尾蓝隐藻衰退性最强(-0.035)。在与泊假鱼腥藻、卷曲鱼腥藻等的持续竞争中,处于衰退状态的种类生态空间会有逐渐缩小的趋势(例如尖尾蓝隐藻),而湖泊假鱼腥藻的优势度可能会进一步的扩大,甚至发生水华。湖泊假鱼腥藻、卷曲鱼腥藻等发展性最强的种类具有产毒潜力[48- 50],而衰退性的种类大多不产毒,如微小平裂藻、尖尾蓝隐藻等[51]。一旦发生产毒蓝藻水华,该地区水库的水质安全和水体功能将遭遇严峻考验。 图4 研究水库浮游植物优势种生态位重叠(ΔSOij) 和生态响应速率(R)Fig.4 Niche overlap (ΔSOij) and ecological response rate (R) of dominant phytoplankton species 群落物种间的总体关联性反映了群落演替的进行阶段和群落的稳定性,一般而言,随着演替的持续进行,群落结构将逐渐趋于完善和稳定,种间关系也将从负关联逐步趋向于正关联,直至种间达到稳定共存的局面[30]。在同一环境中,种对间的正关联指示着对某一方或双方有利的相互作用[52],负联结则表明不利于某一方或双方的相互作用[30]。 图5 χ2检验半矩阵图 Fig.5 Half matrix of the association coefficients χ2 test among the dominant phytoplankton species ▲:极显著正相关 (P<0.01),且ad >bc;△:显著正相关(P<0.05),且ad >bc;▼:极显著负相关(P<0.01),且ad 16个优势种中有25个种对存在显著正关联,与其他相关研究[4]相比,正关联显著率较高,暗示了本研究浮游植物群落中物种优势度对物种关联的影响较为显著。 分季节看,4个季节除夏季表现为显著负关联外,其余3个季节均为显著正关联(表5)。负关联表明夏季浮游植物群落比较脆弱,结构简单,易发生波动甚至退化,这可能与夏季大量降雨与水库开闸泄洪导致的水环境剧烈变化有关[24-25],此时浮游植物群落结构稳定性较差,但仍表现为蓝藻的发展状态(夏季6种优势蓝藻ΔSOij>0,图4)。夏季浮游植物群落结构的不稳定性,结合目前已经出现的蓝藻高密度、高优势度与高生态响应速率,可以预测本区域中小型水库夏季出现间断性蓝藻水华的可能性较大,尤其可能出现假鱼腥藻水华。 研究水库中,不论是优势种种类数、优势度指数以及细胞密度,蓝藻始终占据优势地位,尤其是假鱼腥藻,其优势度指数全年最高,夏季平均密度高达1.84×108个/L,明显高于同类型其他水体[53-54]。从发展性来看,假鱼腥藻四季生态位重叠值均大于零,生态响应速率为16种中最高,说明其全年均为发展状态且发展性最强,处于不断掠夺其他物种资源的扩张状态。其他几种蓝藻,包括拟短形颤藻、拉氏拟柱孢藻、卷曲鱼腥藻、细浮鞘丝藻等也具有较为明显的数量优势或发展优势。研究水库种间关联整体较稳定,可在全年大部分时间维持目前的状态,即蓝藻优势,这是由于四川丘陵区人口众多,农业发达,随地表径流进入各类水体的大量营养物质导致其富营养化程度增加[55-56],有利于蓝藻的生存与发展。同时夏季由于大量降水和开闸泄洪导致的环境剧烈变化[24-25],出现了浮游植物群落结构的不稳定,但并不影响蓝藻的优势地位。故而研究水库在夏季很可能发生间断性的、以丝状蓝藻(假鱼腥藻、鱼腥藻、颤藻等)为优势的蓝藻水华。 (1)研究期间共检出浮游植物188种;四季共出现4门16个优势种,其中蓝藻9种,竞争力相对较强的7种藻类中有5种蓝藻,分别是湖泊假鱼腥藻、阿氏拟柱孢藻、微小平裂藻、拟短形颤藻、中华小尖头藻;以蓝藻的现存量为最高。 (2)冬季物种间生态位重叠值在四季中较高;不同季节优势种中发展性最强的多为具有产毒潜力的蓝藻,存在发生有毒蓝藻水华的风险;水温、营养条件等环境因子对优势种生态位宽度、种间生态位重叠值等有明显影响。 (3)全年群落种间总体关联性结果表明该地区水库群落结构较为稳定且存在正向演替的趋势;在夏季出现了浮游植物群落结构的不稳定,但四季均表现为蓝藻的发展状态。 综上所述,四川丘陵地区中小型水库夏季出现间断性有毒蓝藻水华的风险性高,需开展有效水质管理与水体修复。 致谢:内江师范学院生命科学学院张甜、刘天凤、贺雪、李文星、胡琼瑶、段炼同学在野外采样工作和样品处理中给予协助,特此致谢。2 结果与讨论
2.1 四川丘陵水库浮游植物群落结构

2.2 四川丘陵浮游植物群落优势种


2.3 优势种生态位宽度

2.4 优势种生态位重叠
2.5 优势种种间联结性
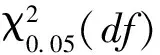

2.6 川东水库蓝藻发展与预测
3 结论
