植物-土壤反馈理论及其在自然和农田生态系统中的应用研究进展
2021-12-24王光州贾吉玉张俊伶
王光州, 贾吉玉,张俊伶
中国农业大学资源与环境学院, 北京 100193
土壤是陆地生态系统的核心,是生物与非生物环境进行物质循环和能量转换的重要场所。土壤是多组分的综合体,具有复杂的物理、化学和生物学性状,提供支撑植物所必需的生长介质以及养分、水分、空气等其它生长条件。土壤性状受到多种环境因素(温度、水分和氧气等)的影响,但植被仍然是影响土壤结构和性质的最重要生物因素。植物通过根系下扎、分泌物、凋落物及调节田间小气候等影响多种土壤性状,包括有机质转化、水分循环及生物群落组成等。这些土壤性状的变化又会反过来影响植物本身及与其共存的植物的生长和健康。这种植物先改变土壤性状,然后土壤反过来影响植物生长的过程,被称作“植物-土壤反馈 (Plant-soil feedback,简称PSF)”[1- 2]。植物-土壤反馈在时间尺度上反映了植物导致土壤中非生物和生物因素的总体变化,同时在空间尺度上反映了生态系统地上-地下部分的互馈效应,因此可以系统探究植物与土壤、资源、环境之间的互作[3]。这一优势使得该理论形成之后的二十多年得到迅速发展,并成为生态学领域一个重要的研究方向。
事实上,植物-土壤反馈理论的基础最早起源于1000多年前的农业生产实践,轮作体系是一个典型的代表[3]。中国古代最早的农学著作《齐民要术》、清朝的《农蚕经》,均记载了作物轮作或套作对提高产量的重要性[4]。在农业生产中,长期单一化种植容易造成连作障碍等问题。连作障碍的主要机制是作物自身根分泌物和凋落物产生的化感物质导致自毒作用、土壤养分钝化、病原微生物累积而产生负反馈作用,而轮作或套作可降低化感物质所引起的这些负反馈作用[5- 7]。在20世纪70年代,生态学家开始关注土壤病原菌的作用,最为知名的是Janzen-Connell假说[8-9],该假说提出成年树种周围容易累积天敌而不利于同种个体种子萌发和幼苗生长, 从而为其他物种提供生存空间,有利于多物种共存和植物多样性形成。1993年 Wim van der Putten通过交互移植试验,证明了土壤病原菌在驱动沿岸沙丘植物群落演替中的重要作用[10]。Bever于1997年正式提出植物-土壤反馈的概念模型及定量化研究方法[1]。之后,该理论得到迅速发展,现已成为生态学领域研究土壤微生物调节植物群落动态、组成和功能的前沿理论和研究方法,为理解和解释群落演替[11- 12]、物种共存和植物多样性形成[13- 15]、多样性-生产力关系[16- 18]等研究开辟了全新视角。当前全球面临着人口增加、粮食安全和生态环境等重大命题,如何更好发展和应用植物-土壤反馈理论,对促进农业和生态环境的可持续发展,提高自然生态系统调节、保蓄能力、增强生态系统服务与功能具有重要的理论和现实意义。
1 植物-土壤反馈理论概述
植物-土壤反馈效应可分为直接反馈和间接反馈[3]。直接反馈是指土壤性状的改变对植物本身及其后代生长发育和健康程度的影响(图1,αA和βB)。直接反馈类型可分为正反馈、负反馈和中性反馈,即土壤性状的改变会促进、抑制或不影响植物自身的生长。而间接反馈是指宿主植物导致的土壤性状的变化对其它共存植物的影响(图1,αB和βA)。间接反馈同样包括负反馈、中性和正反馈,其反映的是一个相对效应大小。例如,当宿主植物引起的土壤性状抑制共存植物生长时,对宿主植物为正反馈效应;反之,为负反馈效应。反馈作用的强度和方向受植物自身和相邻植物的影响。在植物群落中,目标植物及其相邻共存植物与其对应的土壤性状之间往往相互影响,直接反馈和间接反馈同时存在,形成整个体系的“互反馈效应”(pairwise feedback)。互反馈效应是土壤性状对宿主植物及其共存植物影响的净效应,可用公式计算净效应值 (Net feedback=αA-αB+βB-βA)。互反馈效应能够比较群落中植物受土壤性状影响的相互作用关系的强弱。通常,当整个体系中宿主植物引起的土壤性状的改变对同种植物生长的促进作用更高,或者对不同种类植物生长的抑制作用更高时,为正反馈效应;相反地,当土壤性状的改变抑制了同种植物的生长,而有利于其它植物的生长时,则是负反馈效应。

图1 直接和间接植物-土壤反馈效应Fig.1 Direct and indirect plant-soil feedback effects 植物种类A和B分别引起土壤性状SA、SB的变化;同时SA的变化分别影响宿主植物A (αA) 和共存植物B (αB)。同理,SB的变化也会影响宿主植物B (βB) 和共存植物A (βA);两种植物A和B的生长同时受自身种群密度和相互竞争关系 (CA, CB) 的影响;改自文献[19]
2 植物-土壤反馈的作用机制
在广义范围内,植物-土壤反馈包括各种物理、化学、生物学反馈,是生物因素和非生物因素共同作用的结果[2,20]。土壤中非生物因素如养分、水分、分泌物等,以及生物因素如土壤微生物、土壤动物等均可直接或间接影响反馈作用的强度和方向。植物生长对土壤中养分的摄取导致养分含量下降,养分的降低进一步限制植物生长,是一个典型的负反馈[21]。但是,这种由养分差异造成的反馈效应与植物特性以及种间互作相关。例如,生长速率快的植物产生凋落物的数量高,且凋落物容易分解,通常这类植物对高养分供应表现出正反馈作用;相反,生长速率慢的植物对养分需求量较少,凋落物的数量少且分解较慢,低养分供应可以满足生长速率慢的植物的生长,但对生长速率快的植物来说,便会导致负反馈[22-23]。
在狭义范围的植物-土壤反馈,主要指的是植物-土壤生物之间的互作(图2)。因为植物根际可以形成专性土壤生物群落,反过来,土壤生物通过与植物形成共生、寄生、偏利共生等影响植物生长和健康等[24- 25],这种植物-土壤生物互作关系的特异性,为理解和运用植物-土壤反馈提供了更直接的证据和理论支撑。根据土壤生物对植物生长产生的反馈作用的不同,可以将土壤生物分为有益生物和有害生物。一般来说,有益生物促进宿主植物生长,产生正反馈效应(图2a)。例如,丛枝菌根真菌(Arbuscucular mycorrhizal fungi, 简称AMF)能够与80%以上的陆地植物共生,是一种在土壤中分布最广泛的能够促进植物氮、磷营养[26- 27],改善土壤结构[28- 29],提高植物抗病性[30- 31]的有益菌。当宿主植物供给碳源供AMF增殖和扩繁,AMF反过来促进植物生长时,是正反馈作用。然而,AMF也可能更加促进了相邻植物生长,对宿主植物产生负反馈作用(图2c)。同样地,土壤有害微生物如病原菌,对宿主植物为负反馈作用(图2d),但是当宿主植物引起病原菌的变化更能抑制邻居植物的生长时,对宿主植物便是正反馈作用(图2b)。由上可知,有益菌和病原菌均可导致正反馈或者负反馈,取决于植物和微生物的互作方式和强度。

图2 植物-土壤反馈类型Fig.2 Plant-soil feedback types以有益菌和病原菌为例,阐释了植物-土壤反馈的正反馈和负反馈效应;箭头和黑色圆圈分别表示有益或有害作用,实线表示植物-土壤生物之间存在互作,虚线表示两者之间无作用或互作较弱,作用强度的大小由线条的粗细表示;其中,正反馈来自于与宿主植物结合的有益微生物(a),负反馈来自宿主专性病原菌的效应(d);另外,植物对有益微生物不对称的响应也可导致负反馈(c);例如,植物A能够促进有益微生物Y的扩繁和生长,而Y对植物B的促进作用更大,因此植物A相对植物B就是负反馈效应;类似的,对病菌不对称的互作关系也可产生正反馈(b)。例如,植物A对病原菌Y的抗性较高,且促进作用更强,而Y主要抑制植物B的生长,因此植物A相对植物B就是正反馈效应;改自文献[19]
土壤生物和非生物因素间的反馈作用是相互影响的。土壤生物影响有机质降解、团聚体形成、氮循环等诸多生物地球化学过程,反过来,土壤非生物因素也影响着土壤生物功能的发挥。例如,在养分较低的情况下,菌根真菌可以帮助植物吸收养分,产生正反馈;在养分较高的条件下,植物对于菌根真菌的依赖性会降低,向地下投入的碳源会减少,菌根真菌的数量以及活性会降低,菌根真菌会与植物竞争养分,产生中性甚至负反馈[32]。干旱影响土壤细菌和真菌的群落组成[33-34],且干旱对细菌群落组成的影响比真菌更大[35]。但在干旱条件下,植物会选择适应干旱环境的微生物类群,如增加放线菌的丰度[36],当再次受到干旱胁迫时,这些微生物的正反馈作用能够帮助植物更好生长[37- 38]。
3 植物-土壤反馈理论在自然和农田生态系统的应用
3.1 外来植物入侵
在入侵我国农林水生态系统的520余种外来有害生物中,入侵植物就占51.5%[39]。外来植物的大量繁殖和蔓延,会抑制当地植物的生存和发展,降低生物多样性,严重危害入侵地生态系统体系的稳定和安全。天敌逃逸是外来物种成功入侵的主要原因之一。一个外来种被引入到新的生境后,逃避了原生地病虫害的侵害,相对于本地物种,呈现出中性或者正的反馈作用,从而导致其在新栖息地大面积生长、繁殖和扩散[40]。在一些情况下,入侵种也会受到本地病原菌的侵害,然而,相对于本地物种,它们的感病性更弱,这样即使在负反馈的效应下,同样可以形成入侵优势[41]。此外,外来物种也可以通过和有益微生物共生促进其正反馈作用。例如,加拿大一枝黄花(Solidagocanadensis)在我国是一种危害极大的外来入侵植物。这种植物改变了AMF的群落组成,促进了Glomusgeosporum的扩繁和生长,提高了一枝黄花对本地物种鸡眼草(Kummerowiastriata)的竞争能力[42]。AMF促进外来植物入侵定殖的作用机制包括改善入侵植物的营养成分,提高它们的抗逆性等[43]。也有研究表明,有些入侵植物对AMF的依赖性较低,降低了土著AMF的丰度,抑制了对AMF依赖性较高的本地植物的生长,使入侵植物间接获得竞争优势[44- 45]。
进一步研究发现,随着时间推移,入侵植物也可产生负反馈,使其失去入侵能力[46]。例如,对新西兰12种入侵植物的研究表明,随着其在当地定殖时间的延长,负反馈作用越大[47]。在对豕草(Heracleummantegazzianum)不同定殖年限(11—48a)的研究中,同样发现了随时间延长,负反馈作用发生变化[48]。这些负反馈作用的作用机制可能是多方面的,如病原菌的积累,有益菌作用的减弱,养分的消耗和化感物质积累等[49- 50]。同时,许多的外来植物并没有形成入侵优势,这可能受入侵地生态环境因子的影响。土壤类型、温度差异、养分高低、气候干旱或湿润,均可能会直接或间接影响植物与土壤环境之间的相互作用,以及外来植物的入侵和演替。
3.2 植物群落演替
在原生演替阶段的早期,由于成土母质中养分的缺乏,能够与根瘤菌、AMF共生的植物成为建群种,呈现正反馈作用[51]。早期演替植物一般生长速率较快,而使其抗病能力减弱,随着时间推移,病原菌定殖于早期演替植物中,因此容易呈现负反馈作用;而晚期演替物种由于对病原菌有较强的抗性,能够在演替后期占据竞争优势。因此,不同演替阶段植被对病原菌响应的不同,在一定程度上影响群落演替的方向[3]。例如,对沙茅草(Ammophilaarenaria)的研究表明,其在新形成的风积沙滩上生长良好,而当沙丘稳定之后,在其根系中就会有病原菌的积累,而使其相对后续植物失去竞争优势[10]。
在次生演替阶段,植物-土壤反馈同样驱动了群落演替,而且作用速度相对于原生演替阶段可能更快。研究表明,早期演替物种呈现负反馈效应,而晚期演替物种呈现正反馈效应[11]。晚期演替植物除了对病原菌具有较强的抗性外,对AMF等有益微生物的响应较强[52],有益和病原菌共同驱动了植物群落的演替[12,53]。
3.3 物种共存和植物多样性
土壤的正反馈作用会导致优势物种的形成,不利于植物共存和体系多样性的维持[19, 54]。正反馈效应的一个典型例子是当优势物种更好结合有益微生物时,便会产生竞争排除,不利于其它从属植物生存。例如,在北美大草原中C4植物是优势物种,且具有较高的菌根响应,使其它植物的生长受到限制[55]。在森林生态系统中存在“本土优势”,即森林中的优势种在分解过程中发挥了自身的主场优势,促进其凋落物在自身生长的群落环境中分解[56- 57]。影响凋落物分解的因素可分为生物因素(树种、土壤微生物活性等)和非生物因素(气候因素、土壤理化性质等)等两类。“本土优势”的作用机制可能是物种选择了能够快速高效降解其自身凋落物的微生物群落,且建立了适合这些腐生微生物生长和繁殖的环境。
在自然生态系统中,病原菌产生的负反馈作用在物种共存、群落构建和物种多样性维持方面的作用越来越受到人们的关注[58]。最早Janzen[9]和Connell[8]撰文系统地描述了热带森林维持较高物种多样性的机理, 形成了Janzen-Connell假说。这一假说认为种子扩散以母株为中心, 邻近母株的种子和幼苗存在较高的死亡率,有助于维持热带森林物种多样性。植物这种负反馈作用一般源自植物本身根系分泌的自毒物质或病原菌的积累。因此,植物群落中的优势物种受到病原菌的影响时,较难形成绝对的生长优势,导致从属物种也能共存生长,利于群落中物种共存和植物多样性的形成[14-15, 59]。然而,负反馈作用不只源于病原菌,有益菌如AMF也可产生负反馈作用。例如,草地植物车前草(Plantagolanceolata)对AMFScutellosporacalospora亲和性高,而Panicumsphaerocarpon则与另两种AMFAcaulosporamorrowiae和Archaeosporatrappei亲和性较高,当两种植物共同生长时,与Panicum结合的两种AMF对车前草的生长正效应更高,导致负反馈效应[60]。因此,在整个植物多样性体系中,AMF可能促进某些从属物种的生长,提高其对优势种的竞争能力,促进整个群落多样性的提高[61]。
3.4 植物多样性-生产力关系
在自然生态系统中,随着植物多样性增加,体系生产力提高[62]。之前对植物多样性-生产力关系机制的解释集中在非生物因素方面即选择效应和补偿效应。选择效应是指在一个物种数目多的植物群落中,相比于一种或物种数目低的植物群落,优势或高产物种出现的概率较高,这些物种通常生长速率快、竞争能力强,在混作中易获得生长优势,提高了群落生产力[63]。补偿效应包括资源利用生态位的分化和种间互惠作用[62, 64-65]。研究发现,植物-土壤反馈作用也影响植物多样性-生产力关系。植物多样性的增加可以提高土壤中碳、氮的含量[66],其中的作用机理可能是植物多样性促进了土壤中微生物的活性,提高了有机质和氮的矿化速率[67]。这些养分资源的增加反过来又促进了植物多样性体系生产力的提高。土壤中的有益微生物如AMF和根瘤菌,也可以通过参与植物氮、磷等养分资源的吸收和利用而影响种间互作和体系生产力。AMF对植物亲和力不同,同时,菌丝网络的存在可调节C[68]、N[69]和P[70- 71]等资源在植物间的分配。在黑麦草 (Loliummultiflorum) 与红车轴草 (Trifoliumpretense) 混作中,AMF促进了红车轴草的生长,降低了黑麦草对其抑制作用,提高了混作生产力[72]。而且,随着AMF多样性增加,草地生态系统生产力增加[61]。根瘤菌的作用主要体现在豆科植物中。例如,在红三叶草和蓝草的混作中,红三叶草可固氮并促进蓝草的氮营养,提高了体系生产力[73]。
此外,土壤病原菌同样能驱动物种多样性-生产力正相关关系[16- 18]。长期单作土壤中往往会积累宿主专性的病原菌,抑制植物生长。在草地[17, 74- 75]和森林[76]生态系统中均观察到病害的发生率和严重程度在物种多样性低的植物群落中较高的现象。植物多样性体系通过稀释或抑制病原菌,减弱负反馈作用的强度,提高混作体系生产力[77- 79]。
3.5 多营养级之间的相互作用
除了植物-微生物之间的直接相互作用,地上-地下互作系统中还存在大量的其他生物,如原生动物、食草性昆虫、分解者、捕食者等,这种多营养级之间的相互作用,参与和介导植物-土壤反馈的过程,影响植物生长与健康[80]。例如,植物根系与土壤生物之间的互作通过改变植物的养分状况、初生及次生代谢产物等,影响地上部昆虫及其捕食者的响应。研究表明,蒲公英植株饲养的毛虫微生物群落与其驯化的土壤微生物群落类似,表明了植物对土壤微生物群落的影响可以通过土壤记忆传递到以后代植物为食的昆虫中[81]。AMF也能通过反馈防御信号调节地下-地上互作。当植物受到害虫的侵害时,AMF的菌丝起到信号传导的作用,可将这种防御信号传递给相邻植物,提高相邻植物的防御能力[82- 83]。
地上-地下部食草性昆虫还能通过影响根系分泌物和代谢产物影响地下部生物群落[84],进而影响植物-土壤反馈响应。例如,地下部植食性昆虫诱导改变根系中生物碱合成以及土壤中真菌的群落组成,当害虫再次取食时,相对于前期没有诱导的植物,诱导植物生长的负反馈作用更小[85]。人工模拟的植食作用,还可以通过刺激植物根系分泌来促进微生物生物量和活性增强,提高土壤的氮矿化速率,从而有利于植物在植食压力下的补偿生长[86]。这种地上部植食性昆虫介导的反馈作用不仅会发生在同物种之间,也会发生在不同种之间,影响后续植物对昆虫的抗性和生长[87]。
3.6 气候变化与植物-土壤反馈间的相互作用
在区域尺度上,植物-土壤互作受温度、降水以及其他气候因素影响。因此,气候变化会影响植物向土壤输入凋落物和根系淀积物,也影响植物根系与有益菌、腐生菌和病原菌相互作用关系[88]。温度升高能够加速植物的生理和代谢,提高植物光合和呼吸效率,同时也会提高微生物活性,促进有机质的降解,对植物生长产生正反馈效应。但是长期的增温会导致干旱,干旱会降低矿化速率和养分释放,影响植物养分的吸收和利用,从而对种间互作和植物多样性造成影响[89-90]。研究表明,干旱会通过影响土壤微生物群落组成产生“遗产效应”(legacy effect),使植物再次受到干旱胁迫时获益[38],进而影响植物种间互作[91],或使植物趋于中性反馈作用[92]。
CO2的升高影响反馈作用。在针叶林生态系统的野外模拟试验中,利用开放式CO2增肥试验技术 (Free air CO2enrichment, FACE),发现CO2浓度增加后增加了根系向土壤中的碳输入,提高了微生物活性和有机质的矿化,加速碳的流失;然而该过程也同时提高了氮的转化和释放,促进了树木对氮的吸收利用、生长和碳的累积,形成正反馈[93]。在美国Cedar Creek试验站的BioCon试验中,同样利用FACE技术,经过20a的长期定位试验后,结果表明,在C3植物群落中,CO2的增加使氮循环由正变负,而在C4植物群落中由负变正,驱动了体系由C3向C4植物的转变,但是具体的作用机制仍不清楚[94]。同时,CO2的升高往往伴随着增温、干旱等其他气候变化因素的产生,因此这些多重因素如何共同影响植物-土壤间复杂的相互作用,有待于进一步探究[95]。
气候变化最直接的后果可能是植物会像动物一样,随着其最适环境的变化发生生境偏移。当植物到达一个新环境后,这些迁徙植物具备类似于入侵植物的特征,遭受的负反馈作用较小[96];同时与有益菌和腐生菌之间的相互作用关系也较弱,因此受温度、降水、养分等环境因素的影响可能较大。但随着迁移时间延长,特异性病原菌积累,负反馈作用会加重[88]。尽管迁徙植物是气候变化的一个结果,在新的生态气候区定殖后,迁徙植物同样能够改变生态系统过程,例如改变养分循环和地上-地下相互过程[97]。在当前全球气候变化背景下,在不同生态气候区,准确预测植物迁徙类型和特征,进行科学管理和干预,对于保障生物多样性具有重要意义[98]。
植物-土壤反馈在受到气候变化影响的同时,也反过来调节气候变化。通常,植物多样性可提高土壤生物多样性[99],反过来,土壤生物多样性的提高能够提高植物群落抵抗干旱、冻害、增温、氮沉降等环境胁迫的能力[100- 101],有利于维持生态系统的稳定性。植物多样性的提高可通过调节土壤微生物的代谢活动[102]、提高AMF多样性[103]等方式,促进土壤团聚体的稳定性和碳的固存[28- 29],减少CO2的排放潜力。此外,禾草类植物与豆科植物混作后可改变土壤中N循环相关微生物,即降低与N2O排放相关的基因表达,减少N2O的排放[104]。
3.7 植物-土壤反馈在集约化农业中的应用
集约化农业生产造成了土壤酸化[105]、土壤结构恶化[106]、环境污染[107]、病虫害加剧[108]等诸多问题[97- 98],严重威胁到土壤健康和农业绿色发展。例如,在农田生态系统中,长期单一化种植使作物病虫害发生严重,是一种典型的负反馈效应。据估算,病虫害每年造成全球粮食产量减产幅度可达20%—30%[109]。作物多样化种植,包括轮作、间作和覆盖作物等可从时间和空间上缓解负反馈,增加正反馈效应[5,110- 111]。对我国作物种植体系的整合分析表明,轮作体系的产量比单作平均提高20%[112]。一方面,轮作显著提高土壤肥力,例如豆科作物的生物固氮作用增加土壤可利用的氮,提高后续小麦的产量和蛋白质含量[113]; 另一方面,轮作通过非寄主作物对病原菌的稀释效应[114],以及提高拮抗土壤微生物的数量[115],缓解土传病原菌的负反馈效应。
间套作是作物多样化种植的另一种重要形式,其在提高作物产量、资源利用效率,强化生态系统服务功能方面发挥着重要的作用[4]。间套作可通过阻挡作用、田间微气候改善、根际分泌物和改变根际微生物群落等物理、化学和生物方面的作用抑制病害[116],降低土壤的负反馈作用[117]。例如,当易感病水稻和抗病性水稻种植在一起时,由于抗性水稻可阻隔稻瘟病菌的扩散,易感病水稻相对单作可以提高89%的产量,同时降低了94%的稻瘟病发病率[118]。小麦与西瓜间作降低了西瓜根际丙二醛含量,增加了多酚氧化酶、苯丙氨酸解氨酶、类黄酮和总酚类含量,减轻了尖孢镰刀菌对西瓜根系的侵染[119]。旱稻通过根系分泌物改变细菌和真菌的丰度,可以帮助抑制西瓜的枯萎病[120- 121]。在间套作体系中,作物多样性的增加还可通过提高土壤有益菌的多样性和丰度等促进土壤正反馈效应。例如,玉米与不同树木间作体系中玉米根系AMF丰富度显著提高[122]。在南方低磷土壤上,玉米与鹰嘴豆、大豆、小麦间作后,菌根真菌的数量随不同体系而发生变化[123]。在玉米/蚕豆间作中,玉米分泌物可提高蚕豆根瘤数量和氮的固定效率[124]。反过来,这些有益微生物的改变又能通过调节氮、磷等养分资源在种间的利用,提高整个体系的养分利用效率和生产力。如AMF可以提高玉米/蚕豆间作体系中玉米对磷的吸收利用和生长[125],根瘤菌和AMF可提高玉米/大豆间作体系中大豆对氮的固定效率,同时促进氮从大豆向玉米的转移[126],促进体系增产。
近年来,我国土壤污染问题日益突出,重金属类、残留农药、挥发性有机物、石油类等各类有害物质严重威胁土壤健康,对作物生长产生负反馈作用。据统计,2014年全国受污染的耕地面积约有0.1亿hm2,土壤点位超标率高达19.4%[127]。在污染土壤修复技术中,植物修复具有成本低、适用性广及协同改善水质、大气等周围环境的特点,因此是土壤污染修复的一项重要技术。植物修复是利用植物吸收、固定、挥发、降解的机能去除或分解转化污染物质。如在重金属修复方面,蜈蚣草可以超累积砷,龙葵可以超累积镉等[128]。土壤微生物是污染土壤修复的另外一项重要技术。例如,恶臭假单胞菌 (PseudomonasputidaF1) 和施氏假单胞菌 (PseudomonasstutzeriOX1) 能分别降解乙基苯和二甲苯,且两者的混合体系能同时实现对两种苯污染物的高效降解[129]。植物-土壤反馈理论可用于指导土壤污染修复。在污染土壤中可通过筛选高效的适应性强、修复效率高的微生物类群,提高修复效率[130]。随着基因组学、培养组学等技术的发展,合成微生物组成为可能。通过筛选功能性高的植物-微生物组合,不仅可促进污染物的降解,还能促进植物生长,形成高效的植物-微生物联合修复体系[131- 133]。
由于人为措施的影响,农田生态系统反馈作用的发挥还受到管理措施的影响。如土壤中过量的氮和磷会降低AMF的多样性和丰度,影响其相应功能[134],增加氮供应提高作物遭受病虫害的风险[135]。而一些合理的管理措施,如施用有机肥、覆盖作物等保护性耕作措施,能够提高土壤微生物的丰度、多样性和活力,改善土壤质量,促进微生物的正反馈作用[136]。同时,土壤微生物反馈效应的方向可能与植物特性有关。例如,从根系形态角度,比根长低的植物对AMF的依赖性较高,且根系表面积较低,降低了病原菌侵染的机率,可能会呈正反馈效应;反之,比根长高的植物具有较大的表面积,更多依赖于根系获取养分,提高了病原菌侵染的概率,易呈现负反馈效应[137-139]。因此,在不同的农业生态气候区,进行合理的作物组合搭配,有助于更好的发挥有益微生物的作用潜力。目前对农田生态系统微生物反馈作用机制的认识,主要集中在根瘤菌、丛枝菌根真菌和某些病原菌等,未来随着高通量测序、规模化微生物分离鉴定和微生物组学研究技术的发展,需要从微生物群落以及其他生物群落角度开展系统研究[140]。
4 展望
当前日趋加剧的气候和环境变化严重影响全球生物多样性格局和分布。如何理解和掌握植物-土壤互作机制,运用植物-土壤反馈理论应对全球气候变化、保护濒危植物、控制外来植物入侵,以及进行可持续的食物、饲料和生物质能源生产,是未来科学研究和实际生产应用中面临的巨大挑战。尽管植物-土壤反馈理论已经在生态领域得到了广泛应用,但对其普遍适用性、以及地上-地下部生物和非生物因素之间的相互作用机制和规律还不清楚;同时,在我国农业绿色发展中利用植物-土壤-微生物互作强化正反馈作用的机理和途径还不明确。未来还需从以下几个方面开展工作(图3)。
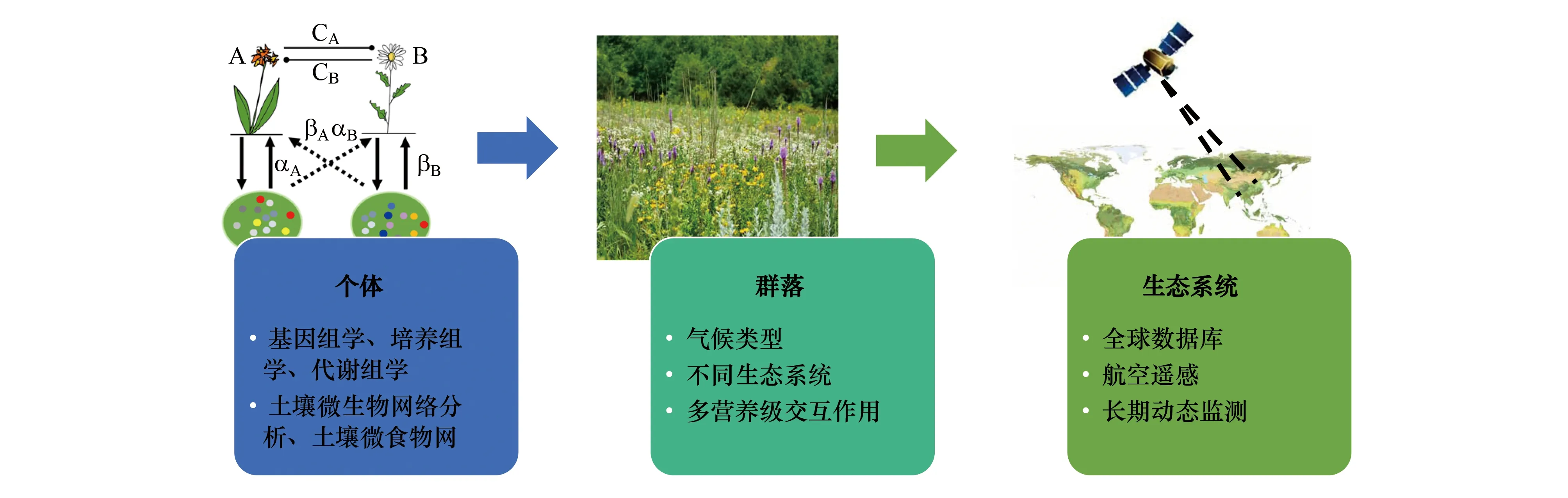
图3 植物-土壤反馈在不同尺度上的研究展望Fig.3 Research prospects on plant-soil feedback at different scales
(1)植物-土壤反馈机制还需进一步深入。土壤作为一个多组分、多维度、多功能的复杂生态系统,在前期研究中,对植物种间互作和群落动态的影响更多是依据植物生长表现而形成的推测性描述,或者利用模型进行的推理性预测。近年来随着基因组学、培养组学、代谢组学等多组学技术的发展,有必要在植物-土壤反馈研究中结合相关技术,揭示植物代谢与菌群功能发挥之间的互作关系,阐明土壤微生物对生态系统地上部种群、群落组成和功能的作用机制。土壤微生物网络结构分析以及微生物食物网是近年来微生物领域比较热门的分析方法。土壤中的微生物并不是单独存在的,而是通过直接或间接的相互作用形成复杂的共生网络。土壤微食物网的生态功能主要体现在微生物和微动物间的相互作用对土壤的C、N 循环过程和植物生长以及农业生态系统中病虫害发生等的影响方面。通过对土壤微生物网络结构和食物网的分析,可以深度解析植物-土壤反馈的机制和过程。
(2)目前对植物-土壤反馈的研究主要集中于对单一生态系统的研究,缺乏不同生态系统之间的比较研究。不同生态气候区的演替阶段以及环境条件如何影响土壤中的碳、氮循环以及分解者、有益菌、病原菌作用和功能的发挥,特定生态气候区的关键微生物类群,如何与食物网中多营养级之间的其它微生物或者土壤动物相互作用,对这些问题进行系统研究,有助于更好地理解、预测和调控生态系统中植被群落动态和生物多样性,提升生态系统服务与功能。
(3)当前植物-土壤反馈的研究多集中在控制条件的室内模拟试验中,如何在更大尺度范围内理解和运用植物-土壤反馈理论是一个巨大的挑战。在未来的研究中,需要借助全球资源数据库、航空遥感技术等的发展[141],在更大区域甚至全球尺度上,对地上-地下互作生态系统,进行长期试验和观测,探究植物-土壤互作的动态变化特征和规律,为生态系统的科学管理和修复提供重要的科学依据。
(4)在农业生产实践中,如何进行作物-土壤-微生物的综合调控,在不同农业生态区,选择适宜的作物基因型和种植方式,提高土壤微生物学潜力,加快土壤内部物质循环和养分周转,减少外源投入,降低土壤污染,增强作物抵抗病虫害的能力,构建良性健康、可持续的植物-土壤互作系统,是未来农业可持续发展亟需重视和解决的问题。随着科学技术的进步,通过对植物-土壤反馈定向调控,可以充分利用和发挥作物多样化种植和土壤生物多样性驱动生态系统多功能性的优势,实现农业生产与生态系统服务的协同。
