基于RNA-Seq分析氨气对肉兔卵丘卵母细胞复合体的影响
2021-12-23吴峰洋杨新宇樊长林陈宝江
崔 嘉,吴峰洋, 杨新宇,樊长林, 陈宝江,3*
(1.河北农业大学 动物科技学院,河北 保定 071000;2.保定市动物检疫队,河北 保定 071001;3.河北省牛羊胚胎技术创新中心,河北 保定 071000)
氨气(NH3)是兔舍中主要的有害气体之一,兔舍中氨气浓度一般为2.28~38.00 mg/m3,且大多数兔舍氨气浓度可达7.6 mg/m3[1-2]。环境中高浓度的氨气会增加器官和组织中氨的浓度[3],并引起许多不利影响,包括生长减缓[4]、代谢异常[5]、免疫抑制[6]、组织和器官损伤[7]以及生殖功能损伤等[8-9]。卵巢是主要的生殖器官,负责产生成功受精和早期胚胎发育所需的感受态卵母细胞[10]。研究发现,7.6,57.0 mg/m3的氨气可显著降低产蛋鸭的卵巢质量及产蛋率[4]。50,100 mg/m3的氨气环境会降低鸡只的产蛋率[11]。12.92~19.00 mg/m3氨气会显著降低大小鼠的产仔量、仔鼠的初生体质量、仔鼠的离乳率及胎次间隔[12]。目前,关于整个卵巢的研究越来越多[13-14]。吴梦等[15]通过对武定鸡卵巢转录组SNP进行分析发现,与繁殖相关的GnRH信号通路中的 PLA2G4A和MAPK12基因可能与武定鸡的停产和产蛋有密切关系。张杉杉[16]通过Illumina 测序平台对高产、低产番鸭卵巢组织进行转录组测序,筛选出了7个与繁殖性状相关的关键基因。吴贤锋等[17]利用RNA-Seq技术获得了不同品种山羊不同繁殖性状的基因表达模式。
卵巢作为一个动态发育的生殖器官,卵母细胞和周围卵丘细胞(CC)之间通过间隙连接的双向通讯对于卵泡区的发育和卵母细胞能力的获得至关重要[14]。因此,将卵丘-卵母细胞复合体(COCs)视为相互依赖的区室的研究非常重要。赵智锋等[18]通过比较川中黑山羊和雷州山羊大小卵泡的mRNA表达图谱,发现GADD45B、TC2和MSMO基因与卵泡发育密切相关。陆婷婷等[19]采用高通量测序技术对发情期川中黑山羊的卵巢基质、大卵泡和小卵泡进行转录组测序,筛选出INHA、TNFRSF19等15个与卵泡发育相关的基因,主要富集在内质网蛋白质加工、类固醇生物合成、卵母细胞减数分裂等信号通路。谢梦圆等[20]采用Illumina测序技术对添加OrexinA的黄体颗粒细胞进行测序后发现,cGMP-PKG信号通路可能在OrexinA调控绵羊卵巢黄体化颗粒细胞发育中发挥重要作用。兔具有“不争地、少争粮、易管理”等优点,且多胎高产,因此使其卵巢保持活跃状态从而促进卵泡发育、诱导母兔发情,对肉兔的繁殖具有重要作用,然而关于氨气对肉兔的卵丘卵母细胞转录组研究鲜见报道。
本研究以肉兔为研究对象,应用高通量测序技术对COCs进行转录组测序,比较分析氨气对COCs的差异表达基因,揭示氨气对COCs的毒性机理,为氨气影响肉兔繁殖性能相关候选基因筛选奠定基础。
1 材料与方法
1.1 试验设计将100只体质量为(1 311.15±144.69) g的35日龄雌性IRA肉兔随机分为CG和LAC两组,分别置于氨气浓度为0.0,7.6 mg/m3的环境控制仓中,每组50个重复,每个重复1只,单笼饲养。自由饮水和采食,光周期为12L∶12D,仓内温度为18~24℃,试验期28 d。
1.2 样品采集试验结束后,称肉兔活体质量,每组随机选取10只肉兔用电击方法实施安乐死。取卵巢,并用含双抗的生理盐水清洗3次,用剪刀剪去卵巢多余组织,用一次性注射器抽取卵巢表面卵泡,捡出COCs,迅速放置于液氮保存。
1.3 卵巢组织学观察卵巢组织经过脱水透明、浸蜡、包埋、载玻片明胶处理、切片、贴片、烤片、脱蜡、HE染色与封片等步骤后得到组织石蜡切片。将石蜡切片置于光学显微镜下拍照,观察组织结构变化并对不同级别卵泡计数。
1.4 RNA文库的构建和测序按照制造商的程序,使用Trizol试剂(Invitrogen,CA,USA)提取总RNA。使用RIN数>7.0的Bioanalyzer 2100和RNA 6000 Nano LabChip试剂盒(Agilent,CA,USA)分析总RNA数量和纯度。根据Epicenter Ribo-Zero Gold Kit(Illumina,San Diego,USA)的说明,纯化RNA。纯化后,在高温下使用二价阳离子将poly(A)-或poly(A)+RNA片段裂解。然后,按照mRNA-Seq样品制备试剂盒(Illumina,San Diego,USA)的操作规程,将裂解的RNA片段反转录以创建最终的cDNA文库,配对末端文库的平均片段大小为300 bp (±50 bp)。然后,按照说明在Illumina NovaseqTM6000(lc-bio,China)上进行配对末端测序。首先,使用Cutadapt除去低质量碱基和不确定碱基的片段,然后使用FastQC(http://www.bioinformatics.babraham.ac.uk/projects/fastqc/)验证序列质量。使用Bowtie2和hisat2将片段映射到兔的基因组,用StringTie组装每个样品的映射片段,用StringTie和edgeR评估所有转录本的表达水平。
1.5 定量逆转录聚合酶链反应(qRT-PCR)验证提取样本中总RNA后用超微量核酸蛋白测定仪(Scandrop100)检测总RNAD值,计算D260/D280比值。采用Aidlab公司反转录试剂盒(TUREscript 1st Stand cDNA SYNTHESIS Kit)进行反转录操作。采用20 μL反应体系:总RNA 500 ng,5×RT Reaction Mix 4 μL,Rondam primer/oligodT 1 μL,TUREscript H-RTase/RI Mix 1 μL,加RNase Free dH2O至20 μL。反转录反应条件如下:25℃ 10 min,42℃ 50 min,65℃ 15 min反应结束后,得到cDNA,-80℃保存。Real-time PCR反应荧光定量反应体系:2×SYBR®Green Supermix(5 μL),Forward primer(0.5 μL),Reverse primer(0.5 μL),cDNA(1 μL),ddH2O(3 μL)。扩增条件:95℃预变性5 min;循环反应95℃ 10 s,60℃ 30 s,40个循环;融解曲线95℃ 15 s,60℃ 60 s,95℃ 15 s。引物序列见表1。

表1 本试验中使用的引物信息
1.6 统计分析使用SPSS 23.0通过One-Way ANOVA和LSD对所有试验数据进行统计分析。数据表示为平均值±标准差。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 氨气对肉兔卵巢组织形态的影响由图1可知,氨气刺激后肉兔卵巢卵泡腔塌陷,卵泡细胞坏死脱落于卵泡腔,部分坏死细胞核溶解凝固。与对照组相比,原始卵泡与初级卵泡数量极显著降低(P<0.01),闭锁卵泡增多(P>0.05)。
2.2 COCs差异表达基因的筛选采用DEseq2法筛选DEGs,与CG组(对照组)相比,LAC组(处理组)COCs共有3 345个显著差异表达基因,其中显著上调DEGs 1 093个,显著下调DEGs 2 252个。表2列出了氨气处理后上调倍数最大的前20个DEGs,表3列出了氨气处理后下调的20个DEGs。

A.CG组卵巢的组织形态(100×);B.LAC组卵巢的组织形态(100×);C.卵巢组织各级卵泡数。黑色箭头(↑)表示原始卵泡;红色箭头(↑)表示初级卵泡;绿色箭头(↑)表示次级卵泡;蓝色箭头(↑)表示卵泡状卵泡;*,**.分别表示差异显著(P<0.05)和差异极显著(P<0.01);CG.0.0 mg/m3氨气;LAC.7.6 mg/m3氨气图1 氨气对肉兔卵巢组织形态的影响
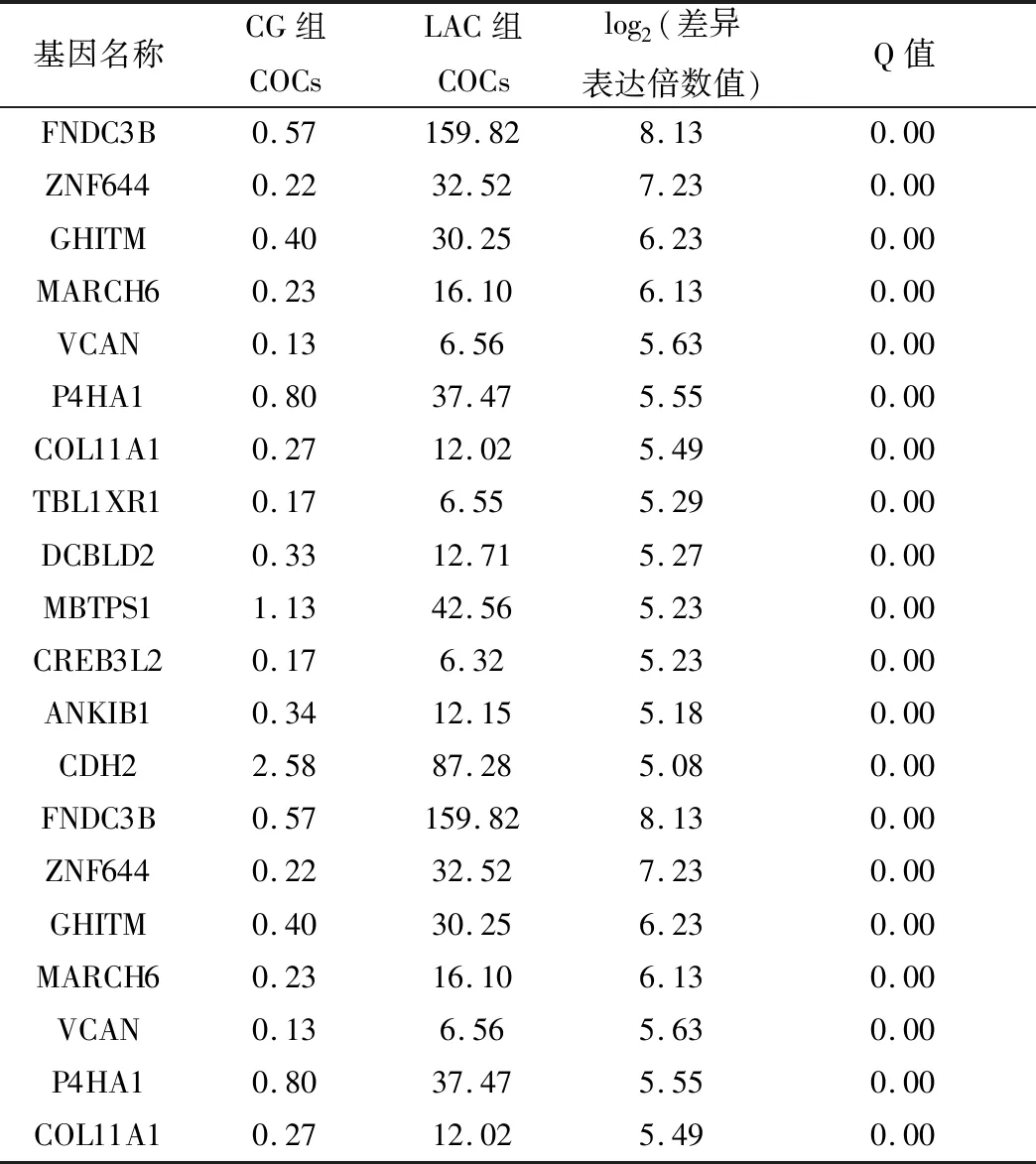
表2 COCs中上调倍数最大的前20个差异表达基因

表3 COCs中下调倍数最大的前20个差异表达基因
2.3 差异表达基因的GO功能富集分析GO分析结果显示,3 345个DEGs归类涉及生物过程(biological process)、细胞组分(cellular component)和分子功能(molecular function)3大类49个二级条目(图 2)。在生物过程分类25个二级条目中占比例最大依次为以DNA为模板的转录调控(regulation of transcription,DNA-templated)、DNA为模板的转录(transcription,DNA-templated)、GTPase活性的正向调节(positive regulation of GTPase activity)和蛋白质磷酸化(protein phosphorylation)。在细胞组分分类15个二级条目中占比例最高依次为细胞质(cytoplasm)、核(nucleus)、胞质溶胶(cytosol)及核质(nucleoplasm)。在 分子功能分类10个二级条目中占比例最大的依次为蛋白质结合(protein binding)、金属离子结合(metalion binding)、ATP 结合(ATP binding)及DNA 结合(DNA binding)。

图2 mRNAs的GO功能注释
2.4 差异表达基因KEGG通路分析KEGG通路分析结果显示,3 345个DEGs被富集到316条KEGG通路,其中PI3K-Akt信号通路(PI3K-Akt signaling pathway)、Rap1信号通路(Rap1 signaling pathway)、Hippo信号通路(Hippo signaling pathway)、 FoxO信号通路(FoxO signaling pathway)和TGF-beta信号通路(TGF-beta signaling pathway)等31条通路为显著富集,各信号通路的基因调控关系见图3。
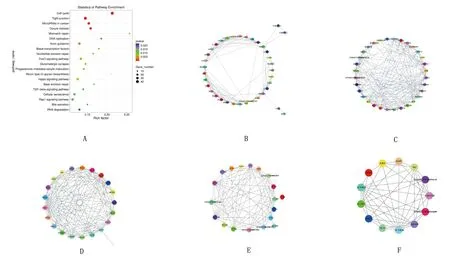
A.mRNAs的KEGG分析;B.PI3K-Akt信号通路;C.Rap1信号通路;D.Hippo信号通路;E.FOXO信号通路;F.TGF-beta信号通路图3 KEGG通路富集分析及蛋白互作网络
2.5 q-PCR验证采用qRT-PCR验证5个DEGs的转录水平(图4), 结果表明,通过qRT-PCR确定所选DEGs的表达模式与通过RNA-Seq的表达模式一致,表明通过RNA-seq确定的DEGs的表达谱是可靠和准确的。
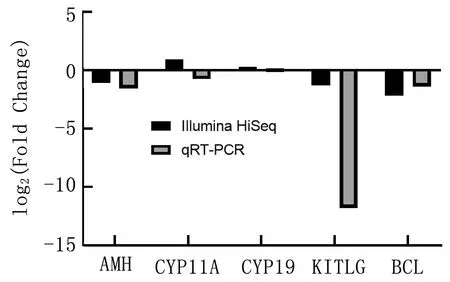
图4 部分差异表达基因的结果
3 讨论
卵巢最基本的结构与功能的单位是卵泡,它们的数量与质量直接决定了卵巢的功能与寿命[21]。卵泡的发育实际上是卵母细胞和周围的颗粒细胞及卵泡膜细胞互相调控的过程,卵泡的形态和功能也随着发育时间而不断变化,卵泡闭锁的潜在机制之一就是颗粒细胞凋亡[22-23]。本试验结果显示,氨气处理后肉兔卵巢的完整性被破坏,原始卵泡与初级卵泡数量极显著降低(P<0.01),闭锁卵泡增多(P>0.05)。卵泡数量的显著降低及闭锁卵泡的数量增多表明氨气损伤了卵巢影响了其功能与寿命。
SIRT1、KIRREL和IGFBP2均是与繁殖相关的基因。SIRT1在生殖细胞和体细胞中均有表达,参与机体多项生理过程[24],并通过影响细胞自噬和推动颗粒细胞凋亡来调控卵泡发育,在遗传毒性、氧化或代谢胁迫下,SIRT1活性增加[25]。KIRREL存在于不同的卵巢细胞中,在颗粒细胞中表达较高,KIRREL与颗粒细胞中MAPK1/3和MAPK14磷酸化的快速增加致使细胞增殖,通过MAPK信号通路对黄体化颗粒细胞产生影响[26]。IGFBP2属于IGFBPs家族的重要成员之一,具有多种生物学功能,在卵泡闭锁时表达水平显著提高[27]。本试验结果显示,氨气处理后的COCs中SIRT1、KIRREL和IGFBP2等基因表达上调,从而影响颗粒细胞的发育能力,促进颗粒细胞凋亡。
Pik3r1主要是通过PI3K-AKT-mTOR信号途径促进基质细胞增殖、抑制细胞凋亡,参与调控胚胎植入[28]。在卵丘颗粒细胞中,KDM2B基因的表达水平随减数分裂的进行而显著增加[29]。LHCGR主要通过改变卵泡类固醇激素生成来影响卵泡的发育[32]。LHCGR接受LH信号开启腺苷酸-蛋白激酶(Camp/PKA)信号通路,激活下游类固醇生成[30],并通过抑制BMP细胞因子来促进卵泡发育[31]。本试验结果表明,氨气处理后的COCs中Pik3r1、KDM2B和LHCGR等基因表达下调,影响类固醇激素生成,促进颗粒细胞凋亡,从而影响卵泡发育。
卵泡的发育过程由原始卵泡的启动开始。原始卵泡启动机制可能与卵泡内外分泌的各种因子、神经递质或卵母细胞-颗粒细胞间的信号传递等有关[33]。本试验中GO功能注释发现,COCs差异表达的基因在生物过程主要富集在DNA转录调控、DNA模板,细胞组分主要富集在细胞质、核、胞质溶胶成分,分子功能主要富集在各种物质的结合;这表明氨气会影响卵丘卵母细胞形态结构,影响卵丘卵母细胞之间的物质交换及信号传递。
卵泡中的卵母细胞周围包裹着各种体细胞,体细胞合成的相关激素物质会通过卵泡液运送到卵母细胞中,以调控和促进卵母细胞的发育成熟和启动减数分裂[34-35]。本试验中对差异基因进行KEGG富集分析发现,氨气处理组与对照组COCs中细胞周期(cell cycle)、紧密连接(tight junction)、胞吞作用(endocytosis)、细胞衰老(cellular senescence)、卵母细胞减数分裂(oocyte meiosis)与孕激素介导的卵母细胞成熟(proge oocyte maturation)通路上的基因表达发生显著变化;这表明氨气处理后COCs中卵母细胞的发育成熟及减数分裂受到影响。
本试验发现,氨气处理后差异表达基因显著富集于PI3K-Akt、Rap1、Hippo、FoxO和TGF-beta信号通路,这些信号通路参与细胞增殖和分化[20]、激素调节等功能[36]。在卵泡中PI3K/AKT-β-catenin信号通路的调控参与FSH以及WNT2介导的卵母细胞和颗粒细胞间细胞连接的形成和解离[37],通路激活会减少颗粒细胞和卵母细胞间连接从而加速卵泡闭锁[38]。Hippo信号通路在调控卵巢颗粒细胞(GCs)与雌性生殖干细胞(female germ stem cells,FGSCs)增殖中均发挥了重要作用[39-41]。FOXO3a可能通过SCF-PI3K-FOXO3a-p27kip1/Bim通路介入新生大鼠卵巢中裸露卵母细胞及原始卵泡卵母细胞凋亡的调节[42]。FOXO3a可能同步诱导促凋亡蛋白Bim和FasL高表达,激活下游细胞凋亡途径,并引起卵母细胞凋亡[43]。TGF-beta信号通路可影响动物卵巢内卵泡的发育调节过程[44]。由此推测,氨气通过激活这些信号通路来影响卵母细胞及颗粒细胞的发育以调控卵丘卵母细胞的增殖与凋亡。
本试验结果表明氨气处理导致肉兔卵巢组织结构完整性被破坏,卵巢储备能力降低。氨气影响肉兔COCs转录组,通过调节SIRT1、KIRREL、IGFBP2、Pik3r1、KDM2B和LHCGR等影响卵丘卵母细胞发育及功能的基因,激活PI3K-Akt、Rap1、Hippo、FoxO和TGF-beta信号通路影响卵母细胞及颗粒细胞的发育,促进卵丘卵母细胞凋亡,进而抑制卵泡发育并影响肉兔卵巢正常功能发挥,最终导致繁殖性能降低。
