云南省凤庆县野放大叶种茶越冬生化成分变化研究
2021-12-23肖涵,侯顺
肖 涵,侯 顺
(昆明学院 化学化工学院,云南 昆明 650214)
在自然条件下,茶树全年经历3次生长-休止轮替,于冬季休眠.目前,对茶树季节轮替变化的研究多基于行业习惯,并关注春夏秋3季[1-3],而有关茶树越冬期的研究尚不多见.研究[4-6]表明,生长周期的变化主要体现在日照、积温、水分含量的变化,导致茶树对多种元素的吸收分布出现差异,Na、Mg、K、V、Cr、Co、Rb、Pt、Se、Ce和Y可以作为不同季节茶叶的指纹因子.物候变化也干扰叶片次生代谢产物,特别是渗透性调节物质脯氨酸、可溶性蛋白和可溶性糖等[7-8].低温导致茶树抗冻耐冬机制启动,并参与生长、发育、蛋白质合成和细胞分裂的代谢组变化[9],使SOD/APX/GR等多种同工酶活性下降[10],以及代谢率大大降低;低温干燥促使茶多糖蓄积为其他季节的2~3倍,酯型儿茶素含量下降,低聚糖类分解为棉子糖、葡萄糖和果糖[11].此外,以上特殊周期变化使冬茶耐泡性提高、滋味醇厚、回甘升高、苦涩感下降,呈现蔗糖甜与花果香,后期转化表现良好.受物候影响,目前我国只有福建、皖南、岳西、贵州和云南凤庆有冬茶出产史,普洱近期也开始研制冬红茶.
云南省临沧市凤庆县属滇红茶代表性产区.凤庆大叶种茶树(Camelliasinensis(Linn.) var.assamica(Masters) Kitamura)种植时间长、面积广,其茶汤滋味强烈刚劲,内含物高[3].滇红创始人冯绍裘先生在《滇红史略》[12]记述:“沿长江南北一带地区不产冬茶,而云南迤西顺宁(凤庆)初冬尚能生产这样的高级红、绿茶叶,诚属可贵”.
为了解凤庆大叶种茶树入冬后叶片生化成分变化,以凤庆县5个野放茶园为样地,按节气采摘一芽两叶至三叶新梢,使用多种技术手段,对12种重要茶叶生化指标(包括6种益生长元素Mg、Al、Mn、Fe、Zn、Cu[13],以及6种重要滋味因子(茶氨酸、脯氨酸、总游离氨基酸(TFAA)、水浸出物、茶多酚、水溶性糖))进行测定,对测定数据进行统计分析.本研究结论不仅可为凤庆县典型大叶种茶树的生长轮替特性累积原始数据,也可为茶行业指导茶园茶树种植、充分开发茶树资源提供参考.
1 材料与方法
1.1 样品采集
2020年10—2021年1月,参考茶行业行规[14-15],即秋茶采摘节气:白露到中秋;冬季:小寒到大寒,于2020-10-02及2021-01-05,在云南省临沧市凤庆县5个大叶种野放茶园向阳坡面中部,5~20棵茶树采摘一芽二叶至三叶新梢(冬季芽叶稀少,其他季节采样以冬季茶株分布中心为起始点,向外辐射).由于冬季各样品芽叶显著缩小,叶片革质化,部分呈现猫耳朵状,色泽发黄、水分含量偏低,且部分叶片有焦边现象.
1.2 实验方法
样品采集后使用纯净水快速冲刷,甩干水分摊晾干燥,冰箱密封冷藏.临用前70 ℃烘干后磨碎至60目.水浸出物含量测定方法参照GB/T 8305—2013[16];游离氨基酸总量及脯氨酸测定采用氨基酸分析仪法(GB/T 8314—2013)[17-18];茶多酚含量测定采用福林酚比色法(GB/T 8313—2018)[19];水溶性总糖含量测定采用蒽酮-硫酸比色法[20];元素测定采用混酸[V(高氯酸)∶V(硝酸)=1∶5]消解后的ICP-MS法(GB/T 30376—2013)[21-22].
1.3 数据分析
用SPSS23.0软件对实验数据进行统计分析.
2 结果与讨论
2.1 鲜叶特征生化成分含量
入冬后野放大叶种茶树鲜叶样品相关生化成分测定结果见表1.
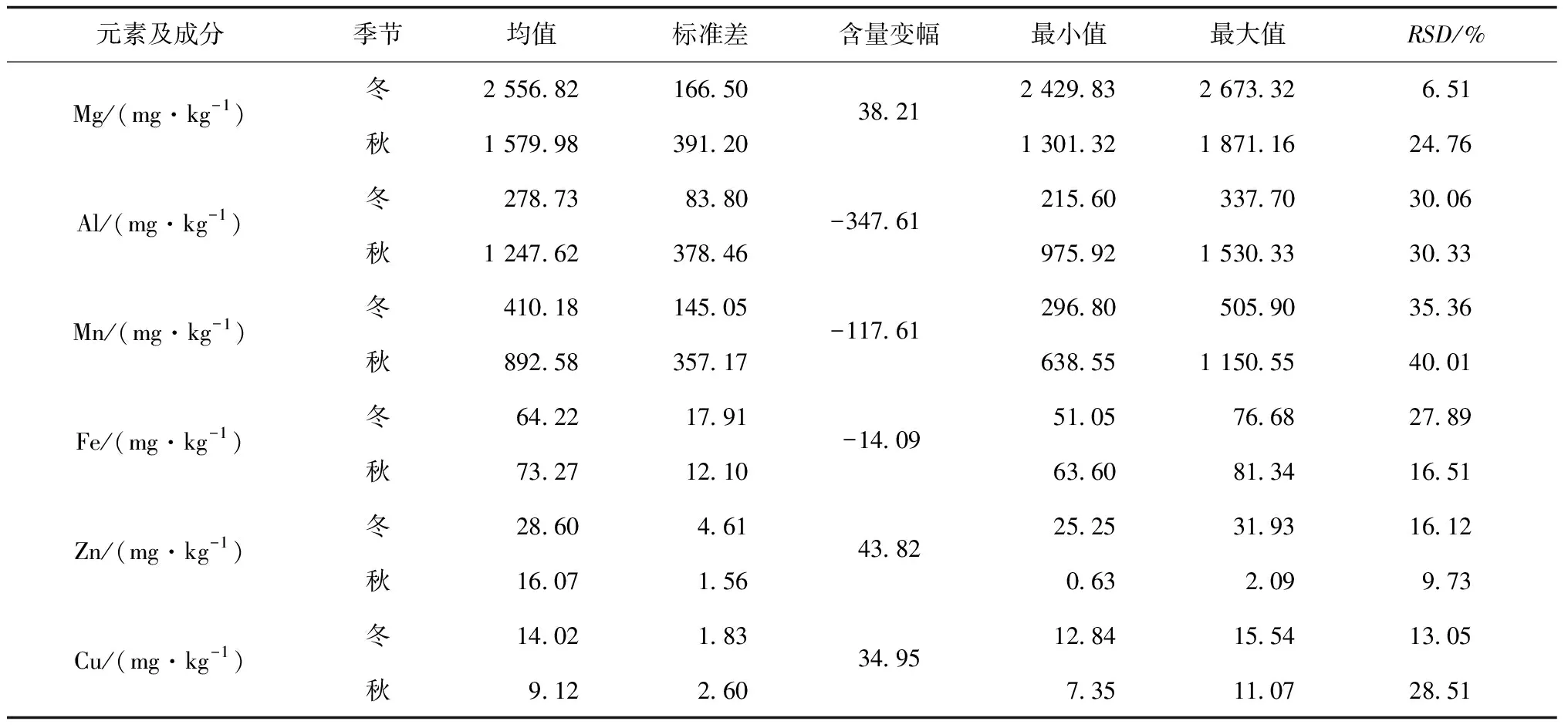
表1 入冬后大叶种茶树新梢特征生化成分含量(n=5)
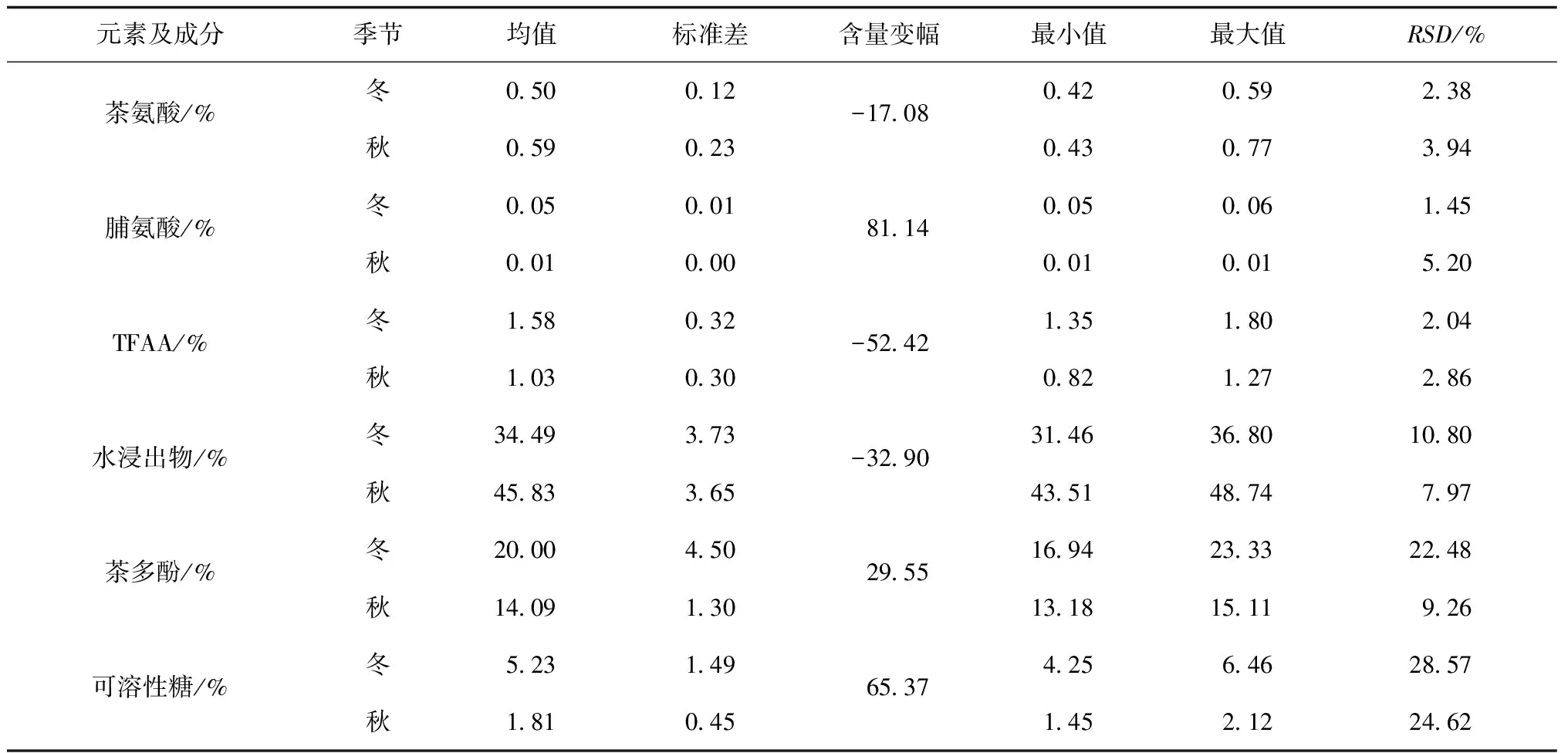
续表1
茶树新梢元素含量主要取决于元素背景值,而次生代谢产物含量主要取决于茶树品种和季节性节律,受茶树生理特性、管理方式等多种因素影响.冬季茶园特殊的水热条件显著改变叶片中次生代谢产物合成机制,供给茶树的氨态氮为茶氨酸、谷氨酸、精氨酸等主要形式,较为稳定地贮藏在茶树根部和叶部[3].
各指标含量均值变化幅度指征秋冬季节性差异.从表1可知,6种益生矿质元素的含量分异明显,各元素含量均值含量水平变异达2个数量级.各元素含量平均值的高低顺序为:w(Mg)>w(Al)>w(Mn)>w(Fe)>w(Zn)>w(Cu),此顺序与地球陆地植物矿质元素含量顺序相近[23].但Al和Mn含量有大幅提高(Al提高1~2个数量级,Mn提高1个数量级),体现了大叶种茶树的富集偏好[24].入冬后,Fe元素含量相对稳定,含量下降的元素为w(Al)>w(Mn)>w(Fe)(变化幅度排序),含量升高的元素为w(Zn)>w(Mg)>w(Cu)(变化幅度排序).入冬后,野放大叶种茶新梢中,除Fe、茶氨酸较为稳定外,脯氨酸、可溶性糖、总游离氨基酸质量分数显著升高,水浸出物质量分数整体下降.
指标含量的RSD值指征不同样地间差异.入冬后,Al元素RSD保持稳定,RSD增加的元素为RSD(Fe)>RSD(Zn);含量变幅降低的元素为CV(Mg)>CV(Cu)>CV(Mn);次生代谢产物中,RSD增加的是RSD(茶多酚)>RSD(水浸出物)>RSD(可溶性糖),RSD减少的是RSD(脯氨酸)>RSD(茶氨酸)>RSD(TFAA).
总体而言,秋冬季节相比,Fe和茶氨酸含量变幅小,RSD小,在茶树体内分布较为稳定,环境影响较少;Zn和茶多酚含量虽有变化,但其RSD亦大幅增加,表明其对多重干扰响应明显,不适宜作为指纹图谱研究对象;TFAA和水浸出物虽有变化,但较为轻微,尚不足以成为季节影响的溯源因子;Mg、Al和脯氨酸含量大幅变化,同时其RSD稳定或减少,说明其受土壤环境影响较小,而对季节变化敏感,可作为冬茶特征因子加以研究.
2.2 特征生化成分成对比较T检验结果
对样品涉及的5个样地的12个指标数据进行成对比较T检验,结果见表2和表3.
由表2可知,入冬后,野放大叶种茶新梢中,除Fe外,其余元素均发生极显著变化.其中,Mg和Al含量极显著下降, Zn含量显著升高,可作为冬茶代表性元素加以研究,其机理可能与蛋白质基因组中A20/AN1-zinc-finger的作用有关[9].
可溶性次生代谢产物与植物越冬期抗寒性相关.此外,在茶产品冲泡过程中,它们又能迅速进入茶汤,成为水浸出物决定茶汤品鉴结果.
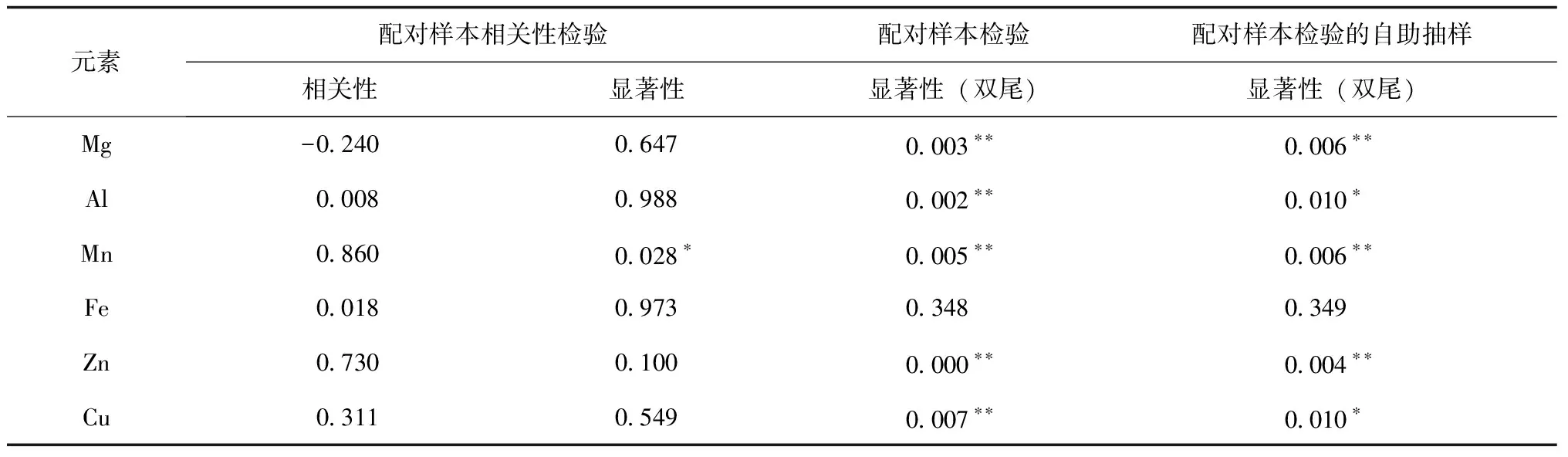
表2 入冬后大叶种茶树新梢元素含量显著性差异(n=5)
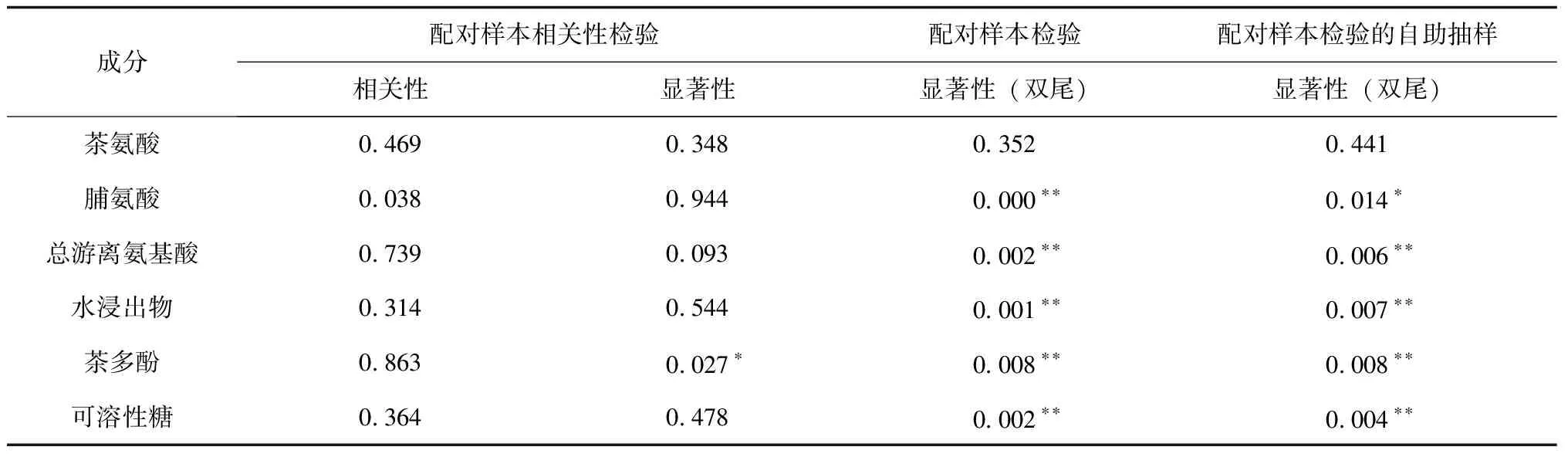
表3 入冬后大叶种茶树新梢次生代谢物质量分数显著性差异(n=5)
在茶树耐冬机制下,茶叶呼吸作用下降,细胞内大分子分解为可溶性物质(蛋白质分解为氨基酸,多糖水解为二糖和单糖等)的速度增加,小分子可溶性次生代谢物聚合速度减缓,以调节细胞渗透压,束缚植物水分,供给碳源和反应底物,提高寒冷抗性.多种氨基酸贮藏在茶树根部和叶部以作为茶树的氨态氮供给源,这使得入冬后总游离氨基酸上升.脯氨酸、可溶性糖在茶树受环境胁迫时,作为渗透调节物质,含量动态增加,以保护植物,安全避冬[7].茶多酚是茶叶中抗氧化、色素合成的必须中间体,大叶种茶茶多酚含量与光照、水热、碳源多个环境因素相关.冬季茶园温度大幅下降,且光照强度较弱,导致茶树体内多酚合成速度明显下降[3].
水浸出物与茶产品耐泡度、茶汤滋味强度密切相关.TFAA不仅体现茶汤良好口感,也促进香气产生和发酵过程.茶氨酸是茶汤中主流风味氨基酸,给茶汤带来鲜爽滋味.茶多酚含量可指征茶汤苦涩程度.水溶性糖指征茶汤甜醇度、回甘度及厚度感.
总体而言,入冬后野放大叶种茶新梢除茶氨酸外,其余次生代谢产物均发生了极为显著的变化.这些滋味物质分布特征与秋茶相去甚远,有必要进行针对性的工艺开发.越冬期茶新梢茶多酚、水浸出物质量分数显著降低,脯氨酸、可溶性糖、总游离氨基酸质量分数极显著升高,其趋势与部分研究成果[7]一致.因此,Mg、Al和脯氨酸可作为越冬茶代表性生化指标进行深入研究.
2.3 鲜叶中特征生化成分相关性分析结果
对越冬期茶叶的上述12个特征生化成分进行相关性分析,结果见表4.
由表4可知,元素中Al、Cu、Mg和Zn与其他成分相关性较强.其中:Al与Cu、Zn、脯氨酸、总游离氨基酸、水浸出物呈显著或极显著负相关,与可溶性糖呈显著正相关;Cu与Mg呈极显著正相关,与脯氨酸、水浸出物、Zn呈显著正相关,而与可溶性糖呈极显著负相关;Mg与Zn、脯氨酸、水浸出物呈显著正相关,与可溶性糖呈极显著负相关;Mn与Zn呈现显著负相关;Zn与脯氨酸呈极显著正相关,与水浸出物呈显著正相关,与可溶性糖呈显著负相关;茶氨酸与所有指标未体现显著相关性;脯氨酸与Al呈极显著负相关,与Cu 和Mg呈显著正相关,与Zn呈极显著正相关;茶多酚与脯氨酸、总游离氨基酸呈显著正相关,与水浸出物呈极显著正相关;可溶性糖与Al呈显著正相关,与Cu和Mg呈极显著负相关,与Zn和脯氨酸均呈显著负相关.
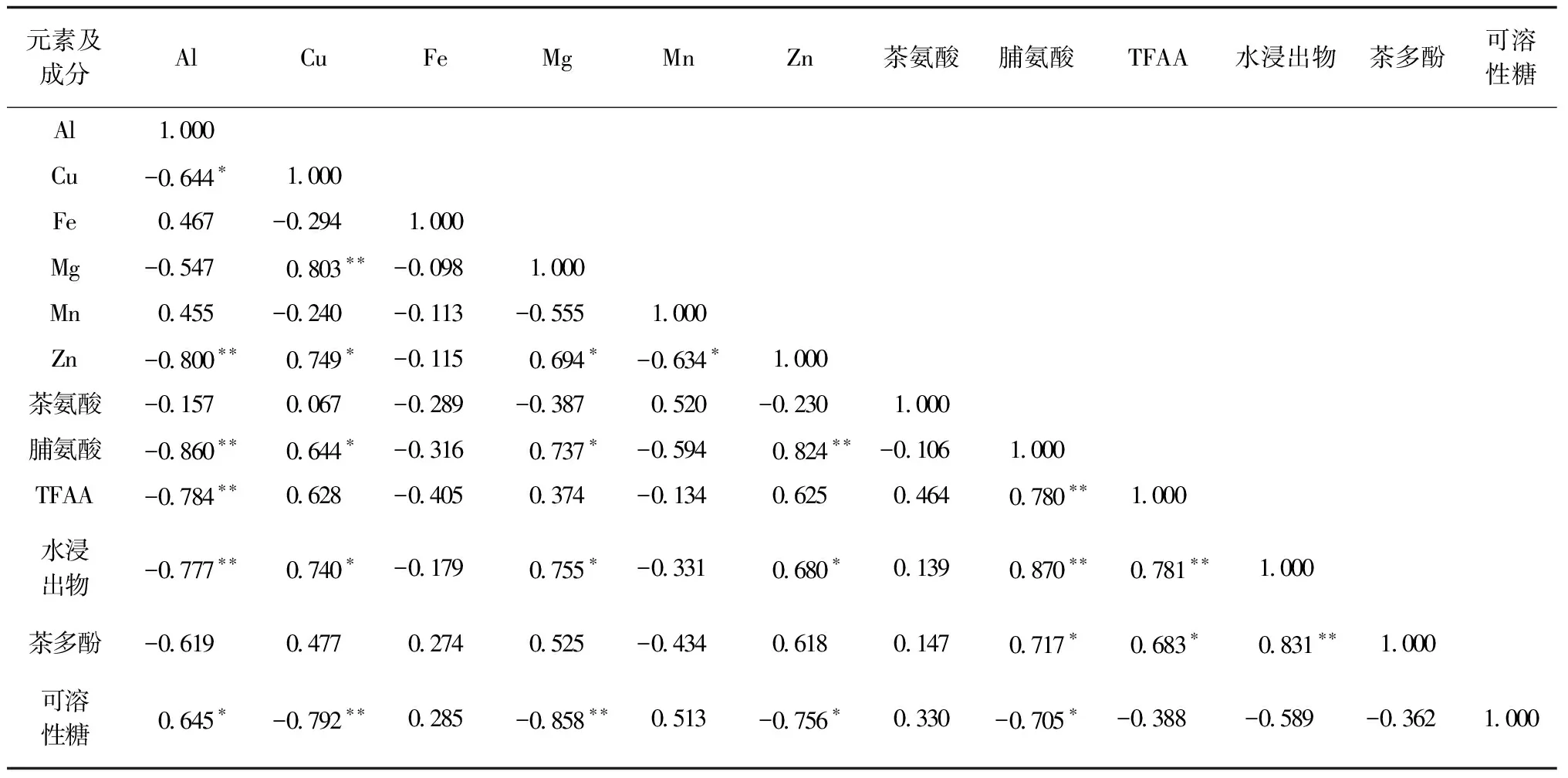
表4 入冬后大叶种茶树新梢特征生化成分相关性分析结果(n=5)
Al与可溶性糖两指标孤立于其他指标,多呈现负相关状态.研究认为,Al在茶树中强富集,可能会对其他元素产生竞争配位,影响其他元素的吸收转运,这也说明虽然茶树是喜铝型物种,但控制种植过程中铝元素背景值会影响其次生代谢过程,进而影响茶产品口感.Al与可溶性糖自身呈显著正相关,可溶性糖的生物合成过程与Al可能关系密切,而与其他次生代谢物的生物合成路径有所区别.Fe与所有指标相关性均未达显著性相关程度,可能在茶树以上指标的新陈代谢过程中未起到关键性作用.Mg和Zn与多个元素体现了相关性,同时也与脯氨酸、水浸出物、可溶性糖呈现相关性.脯氨酸与TFAA、茶多酚、水浸出物、可溶性糖均体现相关性.因此,它们均可以作为冬茶生化成分指纹因子.
值得注意的是,脯氨酸与Zn呈极显著相关性,提示Zn可能参与可溶性蛋白的水解过程,其代谢机理值得进一步研究.
3 结果与讨论
1)样品的6种常量矿质元素的含量分异明显,各元素含量平均值的高低顺序为:w(Mg)>w(Al)>w(Mn)>w(Fe)>w(Zn)>w(Cu).Al和Mn含量在茶树生长过程中强富集.
2)入冬前后,矿质元素分布特征显著不同.入冬后含量下降的元素为w(Al)>w(Mn)>w(Fe)(变化幅度排序),含量升高的元素为w(Zn)>w(Mg)>w(Cu)(变化幅度排序).次生代谢产物除茶氨酸较为稳定外,脯氨酸、可溶性糖、游离氨基酸质量分数显著升高,水浸出物质量分数整体下降.
3) Mn和茶氨酸在茶树体内分布较为稳定,Zn和茶多酚对多重干扰响应明显,TFAA和水浸出物不足以成为季节影响的溯源因子;Mg、Al、脯氨酸受土壤环境影响较少,对季节变化敏感,可作为冬茶特征因子加以研究.
4)成对比较T检验结果表明,元素中Al、Cu、Mg、Zn与其他成分相关性较强.Al与可溶性糖两指标孤立于其他指标,常呈现负相关状态.Mg、Al、Zn、脯氨酸与多个指标均呈现相关性,可以作为冬茶特征生化成分加以研究.
5)Zn和脯氨酸入冬前后变化极为显著,且两者相关性极强,可作为越冬茶树的代表性生化指标进行代谢机理研究.
6)云南茶树冬季仍缓慢生长,但野放大叶种茶树的原生态管理使得嫩梢生长困难,芽头减少,从而降低了原料采收率.因此,建议在11月下旬入冬前对野放大叶种茶树进行轻修剪,以提高产率.
7)由于本实验涉及的冬茶树样品原料较老,水分含量低,氨基酸、多酚质量分数明显下降,可溶性糖较高,因此,建议将其加工为红茶及普洱熟茶.在加工过程中为促进多糖的分解和次生代谢产物的转化,建议提高揉捻程度,采用复揉工艺和发酵工艺,以促进内含物的浸出与转化过程.
8)今后的进一步研究将从茶多糖、儿茶素、黄酮、挥发性成分含量分布特征,以及元素与茶多糖、蛋白质、多酚聚合物等多种次生代谢产物入手,对野放大叶种茶冬季生化特征进行深入调查.
