遗失于巴西国博火灾中的南方“猛禽”
—— 水妖龙与半鸟亚科
2021-12-23王然巍
王然巍
作者单位:沈阳师范大学古生物学院
驰龙科(Dromaeosauridae)是一类中小型的兽脚类恐龙,因电影《侏罗纪公园》,使之成为大众所熟知的明星级恐龙。这种敏捷而凶猛的掠食者,具有锋利的牙齿和特化的第二趾爪,是与鸟类亲缘关系最为密切的非鸟恐龙之一。2021年8月,学术刊物Papers in Palaeontology报道了一个驰龙类新物种——洛氏水妖龙(Ypupiara lopai)。这种新驰龙类恐龙属于驰龙科中的半鸟亚科(Unenlagiinae)——一个无论是地理分布还是形态特征,都与其他驰龙类存在巨大差异的类群。
来自巴西的“猛禽”
这次新命名的驰龙类成员来自巴西南部的乌贝拉巴市郊区。化石材料非常少,仅包括一块不完整的右上颌骨和右齿骨,以及与之关联的若干牙齿。非常遗憾的是,即便是如此稀少的材料,也已经在2018年的巴西国家博物馆火灾中遗失,幸得古生物学家及时对它进行了研究,才使这种来自远古时代的奇特生物能够与更多的人见面。

水妖龙复原图(图源:A.S.Brum等,2021)
古生物学家将这种新恐龙命名为洛氏水妖龙。属名“水妖龙”,来自当地神话故事中的一种水生生物,参与研究的学者们认为,这种新驰龙类是一种在水环境周围以鱼类为食的恐龙。种名“洛氏”则是为了纪念化石的发现者——阿尔伯特·洛帕(Alberto Lopa)。保存水妖龙化石的地层名为马里利亚组,该地层目前还缺乏精确的年代学数据,但地层中的微体化石表明其时代应该是晚白垩世的马斯特里赫特期,介于7200万-6600万年之间。

兽脚类恐龙头骨(图源:网络)
水妖龙的化石虽然保存不完整,但是相关骨骼具有一些区别于其他驰龙类的特征。需要了解的是,在兽脚类恐龙中,眼眶前方存在一个名为眶前窝的结构,在眶前窝下方的颌骨边缘,通常具有神经和血管穿过的微小孔洞——滋养孔。水妖龙的上颌骨仅保存有眶前窝前方的部分,但这部分骨骼上的滋养孔数量极少,仅存在一个,而其他大部分驰龙类以及兽脚类通常具有数量较多的滋养孔。再结合其他种种差异,参与研究的学者相信这是一个新物种。最终的系统发育分析表明,水妖龙属于驰龙科中的半鸟亚科。作为半鸟亚科中的一员,水妖龙具有与其他半鸟亚科成员相似的特征,例如牙齿较小且间距较大,牙齿形状呈圆锥形,齿冠光滑,缺乏隆脊和锯齿,表面具有纵向凹槽。除此之外,上颌骨的形态表明,水妖龙似乎具有一个宛如“鸟头”一样的细长头骨。

水妖龙的上颌骨
驰龙类谱系中的水妖龙与半鸟亚科
驰龙类恐龙可能早在侏罗纪时期就已经存在,但保存下来的化石都是一些零散的牙齿。到了白垩纪时期,驰龙类才迎来了发展的黄金期,并演化出6个主要支系。哈兹卡盗龙亚科(Halszkaraptorinae)是目前已知最原始的驰龙科类群,成员包括哈兹卡盗龙(Halszkaraptor)、胡山足龙(Hulsanpes)和大黑天神龙(Mahakala),化石产自蒙古国南戈壁省的晚白垩世地层。小盗龙亚科(Microraptorine)大部分生存于早白垩世,成员包括中国辽西地区发现的小盗龙(Microraptor)和中国鸟龙(Sinornithosaurus)等,加拿大晚白垩世的西爪龙(Hesperonychus)是该类群唯一已知东亚以外的成员,且生存于晚白垩世时期。真驰龙类(Eudromaeosauria)则代表了驰龙科中最衍化的类群,由蜥鸟盗龙亚科(Saurornitholestinae)、驰龙亚科(Dromaeosaurinae)和伶盗龙亚科(Velociraptorinae)三大支系组成,分别以斑比盗龙(Bambiraptor)、犹他盗龙(Utahraptor)和临河盗龙(Linheraptor)为代表。不过蜥鸟盗龙亚科是否是一个有效的分类群仍存在较大争议。驰龙类的最后一个类群就是以水妖龙为代表的半鸟亚科。
半鸟亚科是一个靠近驰龙科基干位置的类群,在哈兹卡盗龙发现以前,不少学者认为它可能是驰龙类家族中最原始的一个亚科。除了水妖龙以外,半鸟亚科家族的成员还包括鹫龙(Buitreraptor)、半鸟(Unenlagia)、内乌肯盗龙(Neuquenraptor)和南方盗龙(Austroraptor)。新的系统发育表明半鸟亚科与哈兹卡盗龙亚科属于同一个支系,二者互为姊妹类群,并将这个新支系命名为“半鸟类(Unelagiinia)”,较长的头骨和缺乏锯齿的牙齿是它们的共同特征。水妖龙则与半鸟亚科中的南方盗龙构成姊妹类群的关系。

半鸟亚科的谱系位置(图源:自制)
独特的南方“怪龙”
半鸟亚科在驰龙科家族中是一个另类的族群。大部分驰龙类成员来自于北美和欧亚地区,这些地区在中生代时期属于劳亚古陆的一部分。而半鸟亚科则只分布于另外一块超级大陆上,即劳亚古陆南方的冈瓦纳古陆。除了巴西的水妖龙,已知确切属于半鸟亚科的恐龙全部来自阿根廷北部的内乌肯盆地,如鹫龙、半鸟、内乌肯盗龙和南方盗龙。马达加斯加岛的胁空鸟龙(Rahonavis)是否属于半鸟亚科仍有争议,但它所在的地理区域仍属于冈瓦纳古陆的一部分。
鹫龙生存于晚白垩世的早期,因化石产地位于阿根廷的拉鹫地区而得名。鹫龙仅有一种,即模式种——戈氏鹫龙(Buitreraptor gonzalezorum),其化石也是半鸟亚科中保存最完整的,包括一具几乎完整的成年个体骨骼。鹫龙是一种小型驰龙类恐龙,体长约1.5米。它最大的特点是具有一个又细又长的头部,牙齿特征与水妖龙相似。半鸟则是半鸟亚科在晚白垩世中期的代表,这个类群目前已经发现了两种,分别是模式种——科马约半鸟(Unenlagia comahuensis)和佩氏半鸟(Unenlagia paynemili),但二者的化石均不完整,仅保存有少量四肢骨、带骨和椎骨等骨骼。与半鸟处于同一地层层位的半鸟亚科恐龙还有阿根廷内乌肯盗龙(Neuquenraptor argentines),其化石更为稀少,仅有一些跖骨和趾骨,其骨骼的尺寸及比例与半鸟几乎相同,因此不排除内乌肯盗龙是半鸟同物异名的可能性。到了晚白垩世的晚期,半鸟亚科的体型演化到了顶峰,体长超过5米的南方盗龙不但是最大的半鸟亚科,也可能是已知最长的驰龙类恐龙。这种大型驰龙类也具有较长的头部以及与水妖龙相似的圆锥形牙齿。

阿根廷地区的半鸟亚科恐龙(图源:自制)

上图:南方盗龙齿骨;下图:水妖龙齿骨
综合看来,低而长的头骨,以及表面具有凹槽、间距较大且缺乏锯齿的锥型齿,可能是半鸟亚科所具有的共同特征,这些特征在驰龙科家族中极为少见。相比之下,很多其他驰龙类的牙齿扁平宛如刀片,边缘锋利且具有锯齿。同时,鹫龙和南方盗龙的下颌齿骨的外侧存在一条沟槽,内部发育有滋养孔结构。由于水妖龙的齿骨不完整,因此是否具有同样的特征不得而知,但该特征在其他驰龙类中从未发现,但却是驰龙科的“近亲”——伤齿龙科(Troodontidae)的重要特征。
驰龙类家族中的食鱼者
半鸟亚科的食性,一直是古生物学家和恐龙爱好者们津津乐道的话题。不少学者认为,半鸟亚科是一群以鱼类为食的恐龙,或者鱼类至少是它们的猎物之一。但是有关半鸟亚科食性的推测,大部分来自形态学的分析。主要依据包括牙齿的形态、牙齿的数量,等等。
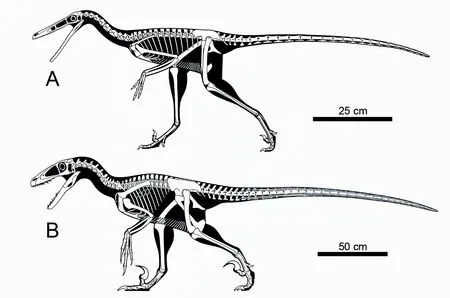
鹫龙(半鸟亚科)与恐爪龙(真驰龙类)

南方盗龙的装架(图源:网络)
棘龙(Spinosaurus)和一些鳄类具有与半鸟亚科相似的牙齿形态。对于体长可达15米的棘龙来说,大部分化石表明它是一种半水生动物,以鱼类为食,特别是2020年棘龙尾部新化石的发现更加支持了棘龙适应水环境的观点。其次是较高的牙齿数量。由于水妖龙的上颌骨保存极不完整,因此学者们无法推测其牙齿数量。鹫龙和南方盗龙的下颌骨可能具有25枚牙齿,一般来说,兽脚类恐龙的下颌齿数通常不会超过20枚,大部分驰龙类一般在11-16枚左右,较多的牙齿虽然不一定反映食鱼习性,但这种特征在具有食鱼习性的四足动物中比较常见。
在驰龙科家族中,被认为具有食鱼习性的类群除了半鸟亚科以外,还包括哈兹卡盗龙亚科中的哈兹卡盗龙和小盗龙亚科中的小盗龙。与半鸟亚科一样,哈兹卡盗龙的食鱼习性也是基于骨骼特征推算而来,例如,同样缺乏锯齿状构造的牙齿、较多的牙齿数量(下颌齿数在25枚左右)和较长的头部,等等。小盗龙的食鱼习性则具有明确的化石证据,该证据来自其胃容物中的鱼类残骸,但另外一些小盗龙化石的胃容物中含有反鸟类和蜥蜴类的残骸,表明小盗龙是一种不局限于鱼类的“全能”捕食者。
由于半鸟亚科的食鱼习性是根据形态特征推测出来的,因此争议较大。有些学者根据后肢骨骼的比例,认为相比于一些后肢相对粗壮的真驰龙类,后肢纤细的半鸟亚科在陆地上奔走的能力更强,因此可能会捕食一些小型陆生动物,例如哺乳类、鳄形类和鳞龙类,等等。但是,一些现代涉禽类显然与该假说不相一致。通过同样具有半夹跖型的小盗龙也许可以推测,半鸟亚科的食性可能比我们想象的要复杂,也许鱼类仅仅是其“食谱”中的一部分,也许它同样也是个“全能”捕食者。
