Cloning, Functional Characterization and Expression Analysis of the elovl4a Gene in the Large Yellow Croaker (Larimichthys crocea)
2021-12-22ZHANGMengLIWanboZHUYangyangWANGWeijiaandWANGZhiyong
ZHANG Meng, LI Wanbo, ZHU Yangyang, WANG Weijia,and WANG Zhiyong, 2), *
Cloning, Functional Characterization and Expression Analysis of theGene in the Large Yellow Croaker ()
ZHANG Meng1), #, LI Wanbo1), 3), #, *, ZHU Yangyang1), WANG Weijia1),and WANG Zhiyong1), 2), *
1),,,361021,2),,266235,3),,361021,
is a fatty acyl elongase which participates in the biosynthesis of long-chain polyunsaturated fatty acids (LC- PUFA). Herein we isolated and functionally characterized thegenein thelarge yellow croaker, and in- vestigated the regulatory effects of transcription factors Hnf4α, Lxrα, Pparα on the activity ofpromoter. Tissue expression pattern revealed thatwaswidely expressed in several tissues and predominantly in brain. Heterologous expression in yeast showed thatElovl4a could effectively elongate both C18 and C20 PUFA substrates to C22 fatty acid. In addition, the affi- nity of large yellow croaker Elovl4a to n-6 series fatty acids is weak and cannot prolong 18:2n-6 and 18:3n-6. Moreover,reporter activities were elevated by 1.31-, 1.39- and 1.48-foldover-expression of Lxrα, Pparα and Hnf4α, respectively. The above findings can enrich the knowledge of biosynthesis pathway of LC-PUFA in the large yellow croaker and contribute to elucidate the LC-PUFA anabolism mechanism in fish.
;; long-chain polyunsaturated fatty acids; promoter
1 Introduction
Long-chain polyunsaturated fatty acids (LC-PUFA) are defined as polyunsaturated fatty acids with ≥20 carbon chainand ≥3 double bonds (Belland Tocher, 2009), such as eico- sapentaenoic acid (EPA, 20:5n-3), docosahexaenoic acid (DHA, 22:6n-3), and arachidonic acid (ARA, 20:4n-6), play- ing pivotal roles in maintaining the integrity and function of biofilm structure (Xiao., 2001; Stulnig and Zeyda, 2004; Marsh, 2008), regulating gene expression (Calder, 2012), promoting cognitive function, protecting eyesight, increasing intelligence (Lauritzen., 2001; Das, 2003; Halver and Hardy, 2003). The proper amount of LC-PUFAs in organisms also minimizes the risk of a number of dis- eases such as nephropathy, diabetes, senile dementia and cardiovascular disease (Kang and Leaf, 1996; Nord., 2001). The desire of meeting the nutritional needs of hu- man kind efficiently has facilitated the studies of bioche- mical and molecular mechanisms underlying LC-PUFA bio-synthesis in fish.
As in most vertebrates, fishes lack Δ12 and Δ15 desa- turases to biosynthesize polyunsaturated fatty acidsand thus they can only convert essential fatty acids (EFA) linoleic acid (LA) and α-linolenic acid (ALA) into LC- PUFAsequential desaturation and elongation reactions of fatty acyl chains (Tocher., 1998; Cook and Mc- master, 2002; Miyazaki and Ntambi, 2008; Park., 2009). However, the capability of LC-PUFA synthesis varies in fishes. In general, freshwater and diadromous fishes are with higher capability in LC-PUFA synthesis compared to marine fish (Sargent and Tacon, 1999). Most of marine fisheslack or have limited capability in LC-PUFA synthe- sis (Gatlin andDelbert, 2003). They need exogenous LC- PUFA to ensure survival, normal growth and development(Sargent., 2003; Lee and Cho, 2008). Lacking the capability of LC-PUFA synthesis is partly due to an un- complete set of key enzymes required in the synthesis pa- thway in marine fish. To date,, an enzyme mainly elongates C20 and C22 PUFA, has only been found in ze- brafish (Tan and Zon, 2011), rainbow trout (Gregory andJames, 2014) and Atlantic salmon (Morais., 2009). Marine fishes appear to lack, which is an essential enzyme for DHA biosynthesis (Jakobsson., 2006).is close toin function and can participate in the biosynthesis of C20, C22 PUFA and especially C24 (Carmona-Antoñanzas, 2011; Monroig, 2011;Castro, 2016).has been isolated and studied in zebrafish (Oscar, 2010), large yellow croaker (Li,2017), yellow drum (Kabeya, 2015), Atlantic salmon (Carmona-Antoñanzas, 2011), rabbitfish (Li, 2017), cobia (Monroig, 2011), orange-spotted grou- per (Monroig, 2012), spotted scat (Xie, 2016) and other fishes. Studies in zebrafish have shown that Elovl4has two subtypes, Elovl4a and Elovl4b, and their functions are differentiated to a certain extent. Elovl4a has the abi- lity to catalyze C20 and C22 substrates producing polyenesup to C24, while Elovl4b may effectively convert eico- sapentaenoic (20:5n-3) and arachidonic (20:4n-6) acids toelongate polyenoic products up to C36 and thus partici- pate in the biosynthesis of very long-chain PUFA (VLC- PUFA) (Oscar, 2010). In addition, other than some bifunctional Fads, such as Δ6/Δ5 Fad or Δ4/Δ5 Fad foundin(Tocher., 1999),(Aga- ba., 2005)and(Li., 2010), while no Δ5 Fad has been identified from marine fishes. Anotherpossibleexplanationisthat marine fisheshavelower enzymatic activities and the conversion efficiency of LC-PUFA is low. For example, the Δ5 Fad activity inand turbot is degenerated, and is unable to convert 20:4n-3 and 20:3n-6 to 20:5n-3 (EPA) and 20:4n-6 (ARA),or the conversion efficiency cannot meet their own physi- ological needs (Tocher., 1999; Huang., 2007). Furthermore, the activity of the newly found Δ4 Fad was also low in(Li., 2010) and(Navarro-Guillén., 2013)Moreover,LC- PUFA is easily accessiblein the wild circumstances.Thismight be thespeculative reason of the genetic degradation in LC-PUFA synthesis in fishes living in such environment(Ghioni, 1999; Mourente., 2000).
Large yellow croaker (), a typical carnivorous marine fish withimportanteconomic value, distributed widely in southeast China. However, thesyn-theticactivitiesof LC-PUFA is weak and need dietary nu- trients supply of fish oil and fishmeal for normal growth in aquaculture. Given the low yields of fish oil and fish meal globally, this will not only increase the cost but also be unsustainable. In order to understand the biosynthesis of LC-PUFA in the large yellow croaker, we herein cloned the full-length cDNA ofand studied its function using heterologous expression in yeast. In addition, the underlying regulatory effects of transcription factors (Hnf4α, Lxrα and Pparα) on thepromoter activity were also investigated. Our study enriched the knowledge of bio- synthesis pathway of LC-PUFA in the large yellow croa- ker and laid a foundation for elucidating the anabolism mechanism of LC-PUFA. Furthermore, it may contribute to reducing the consumption of fish meal and oil in aqua- culture in the future through enhancing theLC-PUFAsyn- thesisability of the large yellow croaker.
2 Materials and Methods
2.1 Ethics Statement
Large yellow croakers used in this study were collected from a mariculture farm in Ningde, Fujian, China. The pro- tocols of fish experiments involved in the study were ap- proved by the Animal Care and Use Committee of Fish- eries College of Jimei University. The brain, eye, gonad, li- ver, kidney, head kidney, spleen, muscle, intestine, stomachand swim bladder tissues were harvested and stored at −80℃ until RNA extraction.
2.2 Molecular Cloning of L. crocea elovl4a and Sequence Analyses
We aligned themRNA (NM_0011 95552.1) against the assembled genome (GenBank acces- sion: GCA_003711585.2) of the large yellow croaker us- ing local blastn program, and found two most significant hits on the chr14 and chr12, respectively. Thegene on the chr14 has been reported previously in the large yel- low croaker (Li., 2017). The newly discoveredon the chr12 was with high sequence similarity toin other fish species and we designated it asThe retained nucleotide sequence ofwas used to de- sign primers (shown in Table 1). A fragment was obtainedpolymerase chain reaction (PCR) and further extended by SMARTer®RACE 5’/3’ Kit (Clontech, USA) to pro- duce full-lengthcDNA using the total RNA iso- lated from brain tissue of the large yellow croaker using TRIZOL Reagent (Invitrogen, USA).

Table 1 Primer name and sequences
The amino acid (aa) sequence deduced from thecDNA was aligned with orthologues from otherspecies using MegAlign software. TMHMM 2.0 (http:// www.cbs.dtu.dk/services/TMHMM/) was used for mem- brane protein structure prediction. The phylogenetic tree of amino acid sequences was constructedneighbor-joining method in MEGA5 software with 1000 times of bootstrapping.
2.3 Quantitative Real-Time PCR (qRT-PCR) Analysis
Total RNA was isolated from brain, eye, gonad, liver, kidney, head kidney, spleen, muscle, intestine, stomach and swim bladder using TRIZOL Reagent (Invitrogen, USA), respectively. First-strand cDNA was synthesized from 2μgof total RNA using GoScriptTM Reverse Transcription Kit (Promega, USA) according to the manufacturer’s instruc- tions. The cDNA template was diluted 20 times, andwas used as internal reference gene. Specific primers were designed using Primer Premier 5.0 (Premier Biosoft). Reaction procedure was performed on a thermal cycler LightCycler 480 (Roche, USA) as follows: 95℃ for 5min; 95℃, 10s; 60℃, 30s for 45 cycles.The relative expression levels were calculated following the 2−ΔΔCtmethod (Livak and Schmittgen, 2002).
2.4 Heterologous Expression of elovl4a in Yeast S. cerevisiae
The ORF sequence ofwas amplified from brain cDNA using primers contained restriction en- zyme sites ofI andI listed in Table 1 and Prime STAR®GXL DNA polymerase (Takara, Japan). The am- plified fragment was ligated into the pYES2 vector (Invi- trogen, USA) to form the recombinant plasmid pYES2-Then the recombinant plasmid pYES2-and pYES2 (control) were separately transformed into(strain INVSc1) competent cells using. EasyCompTMTransformation Kit (Invitrogen). Yeast selection and culture was carried out inminimal medium minus uracil (SCMM-uracil) using galactose induction of gene expression as described in pre- vious reports (Hastings, 2002; Li., 2010).Re- combinant yeast cultures were supplemented with the po- tential fatty acid (FA) substrates (ANPEL, China),inclu- ding α-linolenic acid (18:3n-3), linoleic acid (18:2n-6), γ-li-nolenic acid (18:3n-6), eicosapentaenoic acid (20:5n-3), ara- chidonic acid (20:4n-6), docosapentaenoic acid (22:5n-3), and adrenic acid (22:4n-6). The FA substrates were added to the yeast cultures at a final concentration of 0.5mmolL−1(C18), 0.75mmolL−1(C20) or 1.0mmolL−1(C22), respec-tively, considering the uptake efficiency decreases when chain length of FA increases.
2.5 Fatty Acid Analyses
After culturing for 48h at 30℃, yeast samples were col- lected in a 50mL tube by cryogenic centrifuge at 2500rmin−1for 5minat 4℃. The supernatant was discarded, and the pellet was rinsed twice with 5mL ice-cold Hanks’ ba- lanced salt solution to harvest yeast cells (Sigma, USA). The cells were then homogenized in chloroform/methanol (2:1, v/v) containing 0.01% butylated hydroxytoluene (BHT)as antioxidant (Folch., 1957). Fatty acid methyl esters (FAME) were prepared with BF3-methyl alcohol and ana- lyzed using Agilent 6890N gas chromatography equipped with a CD-2560 capillary column (CNW, China) and hy- drogen was used as the carrier gas. The oven thermal gra- dient was set from an initial temperature of 140℃ and reach- ed to 240℃ by increasing of 4℃min−1and then remained at 240℃ for 30min,and the injection port and detector temperatures were both set to 260℃. FAMEs were identi- fiedcomparing the retention time with each known stan- dard.
2.6 Expression and Reporter Plasmids Construction
The ORF regions of transcription factors Hnf4α, Lxrα and Pparα were amplified using cDNA from brain and li- ver with primers containing restriction enzyme sites forI andIII. The promoter region ofwas amplified using genomic DNA with primers contain- ingI andIrestriction sites (Table 1). An in-fu- sion clone kit (Takara) was used to construct the expres- sion plasmids pCS-, pCS-, pCS-and re- porter plasmid pGL-. Recombinant plasmids were extracted and purified using TransGen Plasmid Mini Kit (Beijing, China).
2.7 Cell Culturing, Transfection and Luciferase Assay
Zebrafish liver cell line (ZFL cells) was used to evaluate the regulatory effects of transcription factors Hnf4α, Lxrα and Pparα on the promoter of. The ZFL cells were cultured in DMEM/F12 (1:1) medium (Gi-bco, USA) supplemented with 10% fetal bovine serum (FBS, Gibco) and penicillin/streptomycin at 25℃ with 5% CO2.The plasmids were transfected using the Lipofectamine 3000 transfection reagent (Invitrogen) according to the manu- facturer’s instruction,and the Renilla luciferase plasmid pRLTK (Promega, USA) was used as an internal control. Briefly, ZFL cells were inoculated into a 24-well plate at a volume of 1.5×105cells per well and transfected when the cell confluence was about 90%. 1.2μg expression plas- mid, 0.4μg reporter plasmid, 0.4μg pRLTK plasmid and 4.0μL P3000 reagent were mixed thoroughly at room tem-perature. Then 100μL diluted Lipofectamine 3000 was add- ed for co-transfection. All assays were performed with threeindependent transfections (transfection efficiency above 86%). After cultured at 25℃ for 24h, dual-luciferase was detected using Dual Luciferase Reporter Assay System (Promega). The details were as follows: ZFL cells were washed twice with 500μL 1×PBS, then were lysed with 100μL 1×passive lysis buffer (PLB) at room temperature for 15min. 20μL cell lysate was transferred onto a plate and then 100μL LAR II and 100µL Stop&GLo® Reagen was added.Firefly and Renilla luciferase activities were mea- suredemploying a Varioskan™ LUX (Thermo scien-tific, USA).
3 Results
3.1 Cloning and Sequence Analyses of the L. crocea Elovl4a
As shown in Fig.1, the full-length cDNA ofgene is 3230bp (GenBank Accession no. MK88 7188), including a 404bp 5’ untranslated terminal region(UTR), a 1863bp 3’UTR and a 963bp open reading frame (ORF). The deduced protein has 320 amino acids and shares higher homology with Elovl4a in other teleost species, suchas(98.7%),(85.9%),(85.6%),(83.8%),(83.4%). It was 67.9%, 70.8%, 71.5%, 72.9%, 71.1%, 69.4%, 68.6% and 72.0% similari- ty toElovl4,Elovl4,Elovl4,Elovl4b,Elovl4b,Elovl4b,Elovl4b andElovl4b, respectively. Additionally, the croakerElovl4a is 70.9% identical to the previously isolatedElovl4, and shares 87.15%, 94.25%, 80.23%,81.82%, 85.27% similaritieswithElovl4b,Elovl4b,Elovl4b,Elovl4b andElovl4b, respectively (Table 2).

Fig.1 Full-length sequence of cDNA and deduced amino acid sequence of elovl4a in L. crocea. Lowercase, cDNA; uppercase, amino acid; round, initiation codon and termination codon; grey shade, ELO superfamily; overstriking type, ployA signals; underline, polyadenylation signal; square frames, site of glycosylation; black shade, phosphorylation sites.

Table 2 GenBank accession numbers and the identity of Elovl4a
The deduced polypeptide sequence contains all the cha- racteristics shared by the microsomal Elovl family, includ- ing one histidine box (HxxHH), four typically conserved motifs and lysine (K) and arginine (R) residues at the car- boxyl terminus (Fig.2). Hydropathy analysis revealed that theElovl4a protein has seven α-helix transmem- brane regions. Phylogenetic analysis demonstrated that theElovl4a is clustered in the Elovl4a group andis more distant with Elovl4 and Elovl4b in the vertebrate lineage (Fig.3).
3.2 Expression Distribution of elovl4a in Tissues
Quantitative RT-PCR analysis revealed thatgene widely expressed in various tissues and the highest level ofexpression was found in brain,which was remarkably higher than in other ten tissues (Fig.4). In the different regions of brain, we found signi- ficantlyhigherexpressionlevelsofin cerebrum, cerebellum, thalamus and optic lobe compared to medulla oblongata (Fig.5).
3.3 Functional Characterization of Elovl4a in Yeast S. cerevisiae
Functional characterization of thewas carried out through heterologous expression ofin the yeast strain INVSc1,transformed with either the emptypYES2 vector or the recombinant plasmid pYES2-. The transformed yeasts weregrown in the presence of po- tential fatty acid (FA) substrates including 18:3n-3, 18:2n-6, 18:3n-6, 20:5n-3, 20:4n-6, 22:5n-3 and 22:4n-6. The chro- matograph analyses showed that the fatty acid composi- tion of the yeasts carrying the empty pYES2 vector were predominant FA normally found in, namely 16:0, 16:1n-7, 18:0, 18:1T (trans-9) and 18:1 (cis-9), toge- ther with the exogenously supplied fatty acids. This is con- sistent with the fact thatdoes not possess fat- ty acid elongase.
Based on GC retention time, 18:3n-3 substrate was con- verted to 20:3n-3 and 22:3n-3 in the FA profiles of the yeasts carrying pYES2-Furthermore, additional peaks corresponding to 22:4n-6 and 22:5n-3 were observed in the presence of 20:4n-6 and 20:5n-3, respectively. How- ever,Elovl4a has a poor affinity toward n-6 fat- ty acids and no additional peaks were observed in the presence of 18:2n-6 and 18:3n-6 (Fig.6).
3.4 Sequence Analyses of L. crocea elovl4a Promoter
Analyzing genomic DNA of the large yellow croakerwith Clontech’s SMARTer® RACE 5’/3’ Kit, a 1730bp (−1249bp–+481bp)gene promoter sequence was identified.Transcription factor binding sites were predict- ed to include specificity protein 1 (Sp1), sterol regulatory element (SRE), NF-Y, Hnf4α, Pparγ-rxrα,. (Fig.7).

Fig.2 Alignment of Elovl4a amino acid sequence of L. crocea with those of other vertebrates. Black shading, same amino acids; gray shading, similar amino acids; numerical, conservative motifs; arrow, residues of lysine (K) or arginine (R) at the c-terminal.
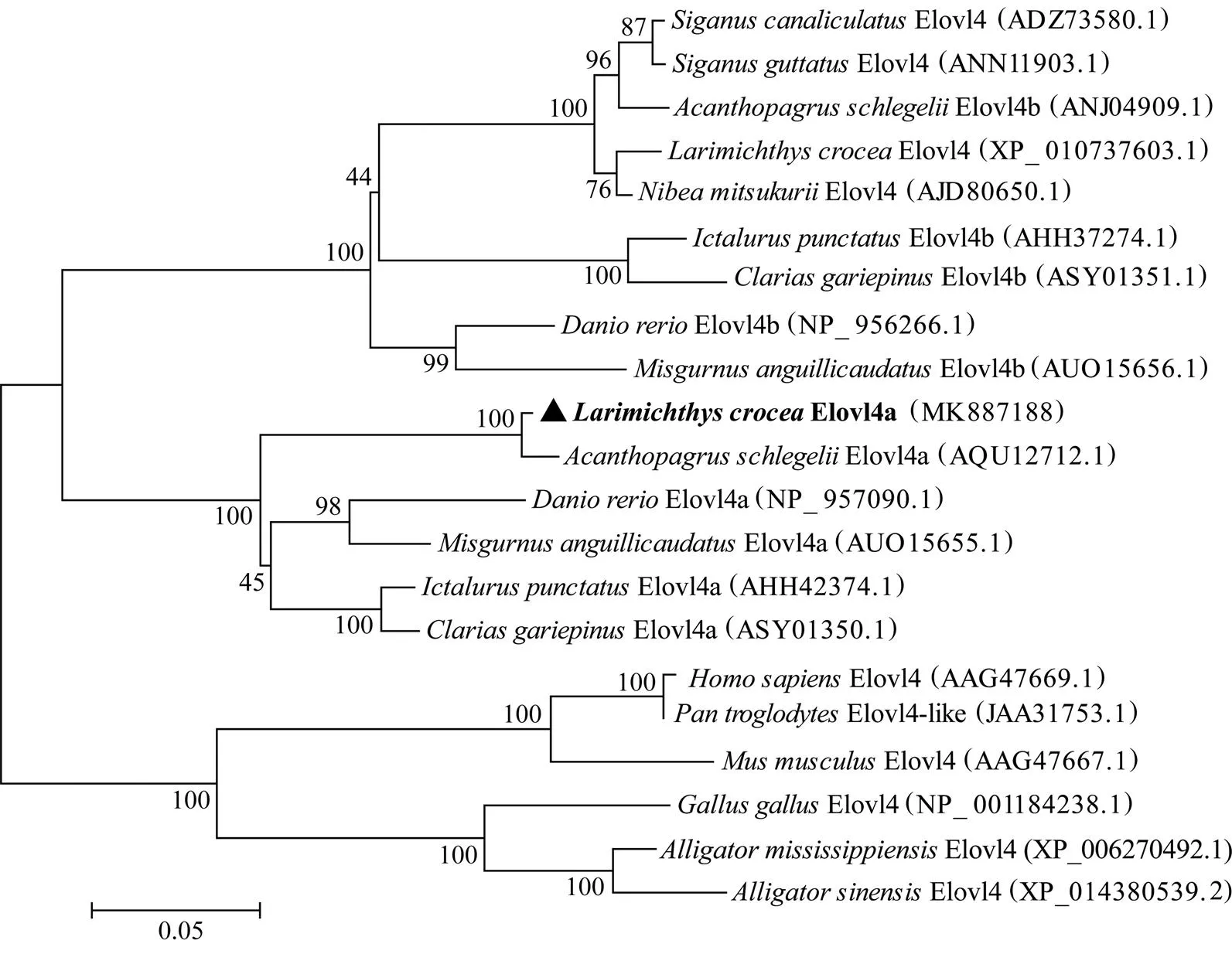
Fig.3 Phylogenetic analyses of amino acid sequences of Elovl4a.
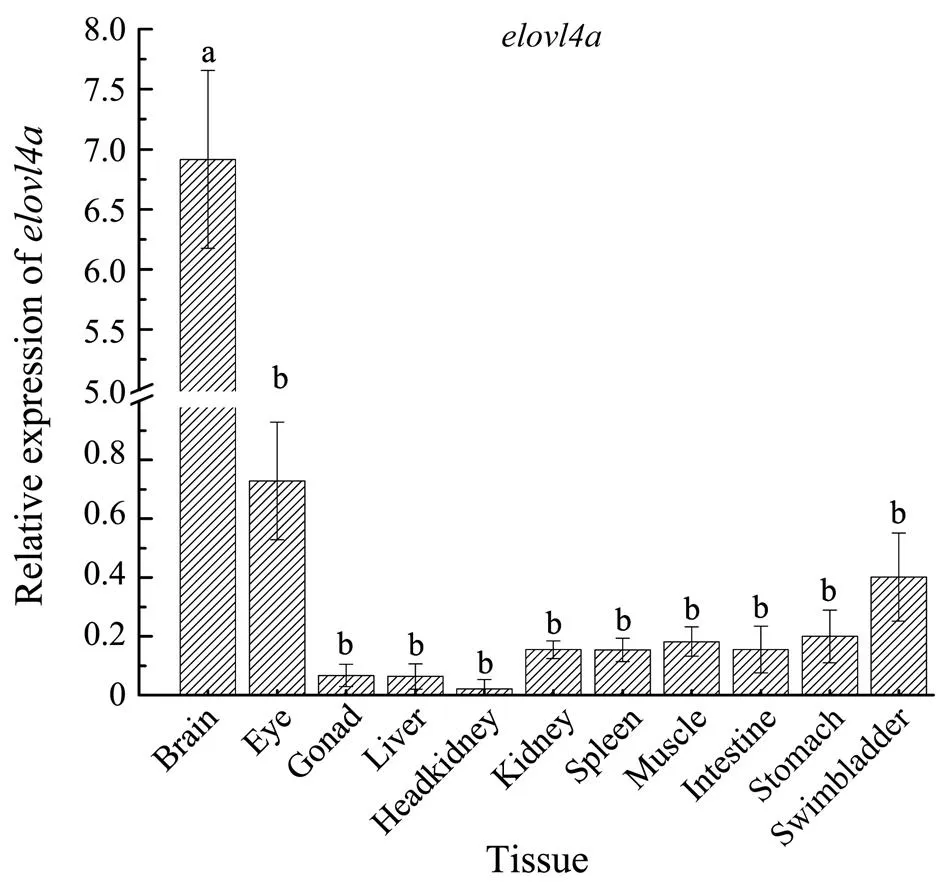
Fig.4 Expression profiles of L. crocea elovl4a indifferenttissues. The different lowercase letters above error bars indicate sig-nificant difference (P<0.05) and the same letters indicate no significant differences.The same rule is applied in Fig.5 and Fig.8.
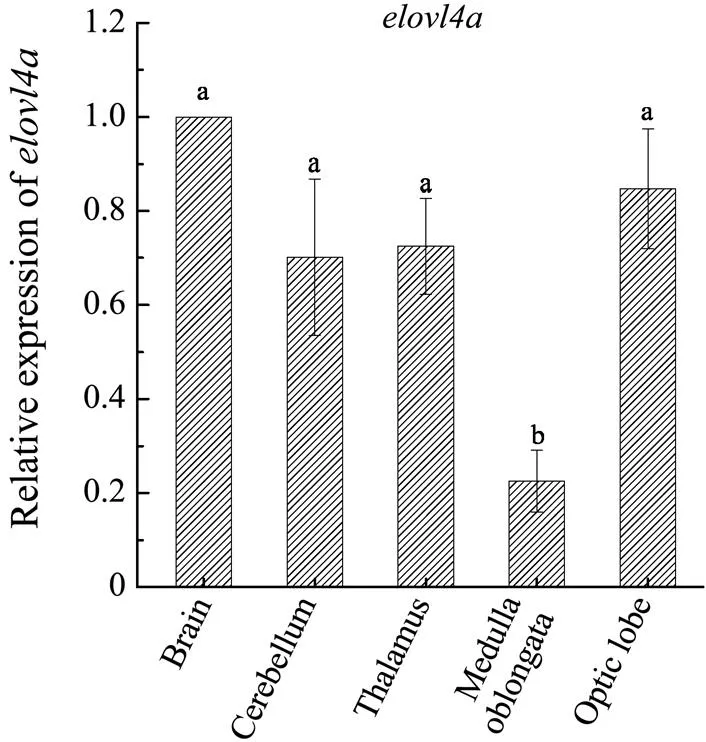
Fig.5 Expression pattern of L. crocea elovl4a in the brain.
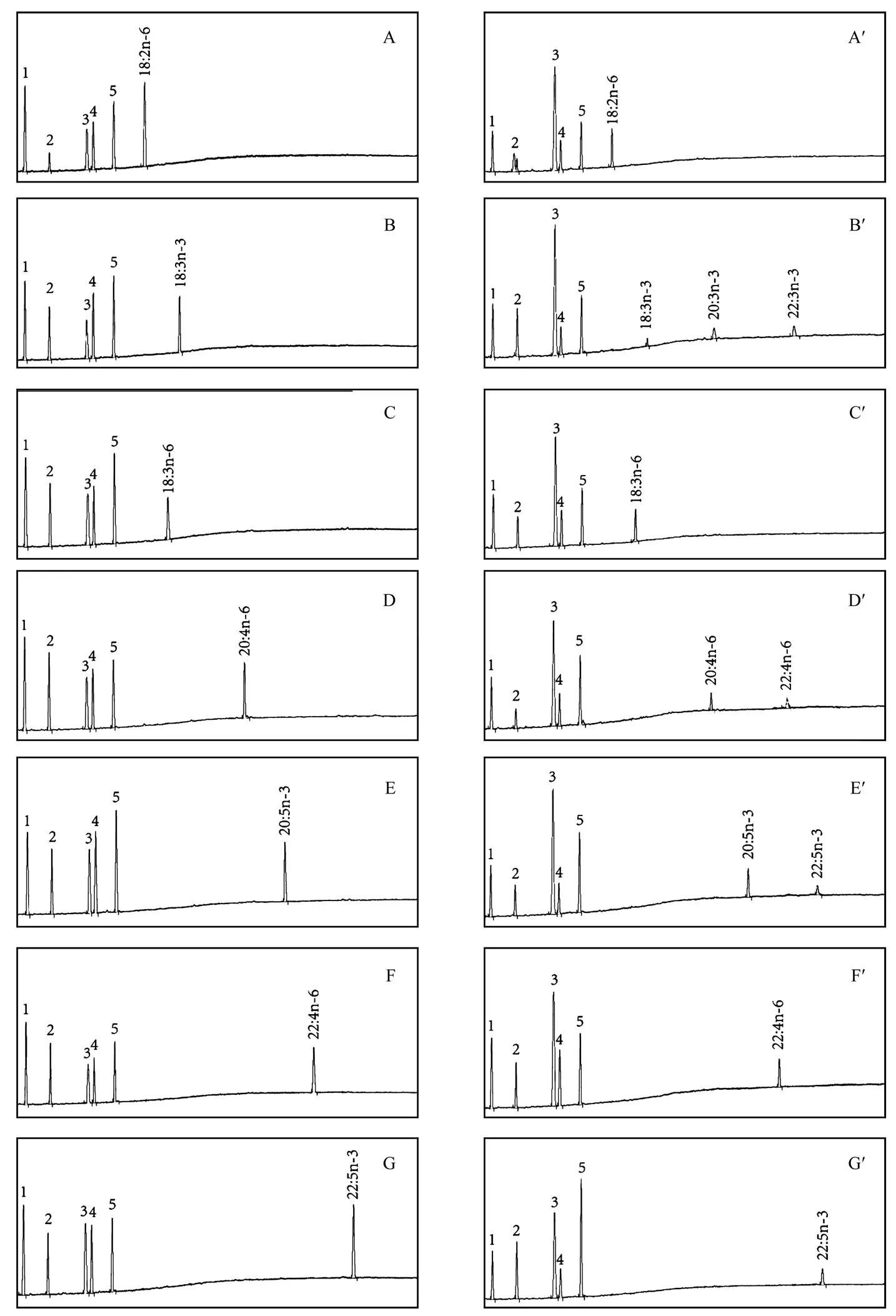
Fig.6 Functional characterization of Elovl4a in Saccharomyces cerevisiae. The numbers 1–5 are the major endogenous fatty acids in the yeast C16:0, C16:1, C18:0, C18:1T (TRANS-9), C18:1 (CIS-9).
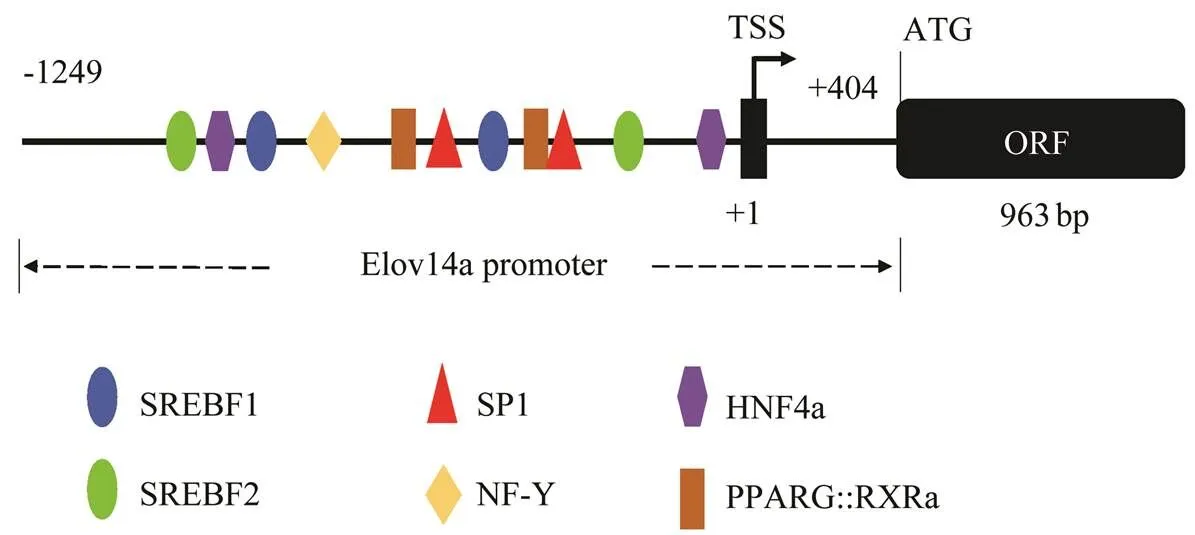
Fig.7 The diagram of transcription factors binding sites in the promoter of L. crocea elovl4a.
3.5 Dual-Luciferase Reporter Assays
Dual-luciferase assay showed that the transcription fac- tors Hnf4α, Lxrα and Pparα all had significantly positive effect onreporter activity (<0.05), but the degree of regulation was different (Fig.8). Compared with the control group, Hnf4α had the highest activation abi- lity, which is about 1.5 times of the control group. Mean- while,reporter activity was increased to 1.31-fold, 1.39-fold by over-expression of Lxrα and Pparα, respectively. It is worth mentioning that there was no sig- nificant difference among the three transcription factors.

Fig.8 The results of dual-luciferase assay.
4 Discussion
Fatty acid elongases (ELO) are able to add 2 carbon atoms to the FA substrates, Cn-CoA, through a series of conden- sation, reduction and dehydration steps (Osei., 1989;Cinti., 1992;Moon, 2003; Leonard., 2004; Vla- dimir and Jonathan, 2007; Ikeda., 2008). Elongases can be broadly divided into two categories. One class spe- cifically elongates saturated and monounsaturated fatty acids, such as,,,.The other class, like,, and, plays a crucial role in the prolongation of PUFAs and is a hot research topic in the field presently (Guillou., 2010).preferentially elongates C18 and C20 PUFAs, with residual conversiontoward C22 substrates (Agaba., 2004,2005; Monroig., 2012).It is commonly believed thatpresents in all marine fishes with high enzyme activity., an enzyme mainly elongates C20 and C22 PUFA, appears to be absent in most of marine fishes and has only been iso- lated from zebrafish (Tan and Zon, 2011), rainbow trout(Gregory and James, 2014) and Atlantic salmon (Morais., 2009) to date.is close toin function and can participate in the biosynthesis of C20, C22 PUFA and especially C24,a very long-chain PUFA (Carmona-Antoñanzas., 2011;Monroig., 2011;Castro., 2016). Such an ability ofhas made it as an essential component which can contribute to DHA biosynthesis through the Sprecher pathway in species that lacks.
In the present study, we isolated a newfamily gene–from thelarge yellow croaker. Analysis of the deduced polypeptide sequence showed that it possesses all the features of the Elovl family including one histidine box, four typical conserved motifs and lysine and arginine residues at the carboxyl terminus (Meyer, 2004;Jakobs- son., 2006), indicating that Elovl superfamily is very conserved in the evolutionary process (Lagali., 2003), andpossibly its function in LC-PUFA biosynthesis is also conserved. Additionally,Elovl4ashares a high ho- mology with Elovl4a in other fishes. Phylogenetic ana- lysis showed that the aminoacidsequence ofElovl4a is clustered in the Elovl4a group and is most close- ly toElovl4a, and is distant with Elovl4 and Elovl4b in other fish species. The clustering pattern ob- served for fish Elovl4 suggested that the function of the two types of Elovl genes might be different.The functions of Elovl4 andElovl4b are similar in fish, whereas Elovl4a is different from them.
Previous studiessuggest that fishes generally possess twoElovl4 enzymes(Castro., 2016), Elovl4a and Elovl4b, and only Elovl4btakes PUFA as preferred sub- strates. Nevertheless, in zebrafish, both of the two genes involved in the synthesis of PUFA. Elovl4a may effective- ly elongate C20 and C22, generating products up to C24 in length, while Elovl4b can catalyze PUFA substrates pro-ducing polyenes up to C36 and thus participates in the biosynthesis of VLC-PUFA (Oscar., 2010).Moreover, Elovl4 in Atlantic salmon, rabbitfish, cobia, orange-spot- ted grouper, Atlantic bluefin tuna and African catfish can produce C36 VLC-PUFA as well (Carmona-Antoñanzas., 2011; Monroig., 2011; Monroig., 2012;Li., 2017;Oboh., 2017; Betancor., 2020).(Kabeya., 2015),Elovl4 (Xie., 2016), andElovl4a are simi- lar to zebrafish Elovl4a, and can only extend up to C24 PUFA. However, previously it was reported thatElovl4 is able to elongate all PUFA substrates up to C32 or C34, except for DHA that is only elongated to 24:6n-3 (Li., 2017). Thus, it is reasonable to believe that Elovl4 from a certain teleost species is sub-functionalized during evolution. Interestingly, the capability of theElovl4a for elongation of 18:2n-6 and 18:3n-6 was notobserved. Possibly there is a competitive inhibition of en- zymes between n-3 and n-6 PUFA, and there is a higher affinity for n-3 PUFA in fish (Tocher., 2002; Belland Dick, 2005; Stubhaug., 2005). We suspected that elon- gation products could have been produced in yeasts at an amount below the detection level. The specific mechanism still needs to be further verified.
The expression levels ofhave been extensively studied,whichexhibited different expres- sion patterns in different tissues in various species.and cobia (Monroig., 2011) and orange- spotted grouper (Li., 2017)are expressed wide- ly in various tissues,and are notably abundant in eye and brain, indicating that these tissues are important metabolicsites for the biosynthesis of LC-PUFA (Agbaga., 2010).The explanation might be related to a large amount of LC- PUFA needed in the brain and eye development, agreeing with the prominent role of LC-PUFA in promoting brain development and retinal formation as well as essential phy- siology (Fernandez-Palacios., 1997; Lauritzen., 2001;Cook and Mcmaster,2002; Jaya-Ram., 2008;Zakeri., 2011).Additionally, unlike(Xie., 2016)and(Kabeya., 2015)whoseis dominantly expressed in the eye, the ex- pression ofis substantially higher in brain than in other tissues. This result is consistent with the fact that the expression levels of fatty acid elongases are ge- nerally high in the brain of various animals (Zheng., 2005,2009). To further investigate whether the synthesis of LC-PUFA in the large yellow croaker is mainly regu- lated by a certain region of the brain, we detected the ex- pressions in cerebrum, cerebellum, thalamus, optic lobe and medulla oblongata, and no significant difference was observed.
It is known that upstream promoters and some transcrip- tion factors, such as peroxisome proliferator-activated re- ceptors (Ppars), hepatocyte nuclearfactor (Hnf), sterol re- gulatory element binding proteins (Srebps) and liver X re- ceptor (Lxr) can regulate lipid metabolism and the synthe-sis of LC-PUFA independently or cooperatively. A 1730bp (−1249bp–+11bp) candidategene promoter re- gion was clonedRACE in the large yellow croaker.Among them, −534–−516 and −405–−396 is the predict- ed specificity protein 1 (Sp1) binding site, a sign of strong promoter activity, which was only found in Atlantic sal-mon Δ6 Fad andFad1promoter (Zheng., 2009;Dong., 2018).Thepromoter might be very active, but whether the site is actually present remains to be examined by subsequent experiments.
Dual-Luciferase assay showed that, like other enzymes,is regulated by transcription factors, such as Hnf4α, Lxrα and Pparα. Ppar has been extensively stu- died in fish, and three subtypes of Ppar have been success- fully cloned from zebrafish, large yellow croaker, Euro- pean sea bass, rabbitfish, Rainbow trout,, Atlantic salmon,. (Andersen., 2000; Ibabe.,2002;Leaver., 2005;Kondo., 2007; Leaver.,2007;Dong., 2017; You., 2017). Pparα and Pparβ indirectly mediate the synthesis of LC-PUFA through re- gulating the expression of genes related to β-oxidation, whilePparγ mainly participates in the lipogenesis. It has been demonstrated that the over-expression of rainbow troutPparα2 andPparα1 and 2 can elevate the Δ6 Fad promoter activity (Dong., 2017). In addi- tion, rabbitfish Pparγ can inhibit the synthesis of LC-PUFAby inhibiting Δ6/Δ5 expression (You., 2017).It is also worthnoting that studies have shown that Pparα has no activation effects on the promoter of fatty acid desatu- rase, whilereporter activity is induced by the over- expression of Pparα. The contributions of Pparα to the pro-moter activities of fish desaturase and elongase are diverse, and we speculated that the activity of fish LC-PUFA syn- thesis-related enzymes were modulated by different tran- scription factors. So far, there are few studies about Hnf4α in fish. In rabbitfish, the changes ofcan affect the ex- pressions of Δ6/Δ5, Δ4,and(Wang., 2018). In this experiment,Elovl4a reporter ac- tivity was 1.31-fold elevated by over-expression of Hnf4α, manifesting that Hnf4α is a key factor in the regulation of LC-PUFA synthesis. With respect to Srebps, existing data indicated that only in the presence of sterol response ele- ments (SREs), Srebp-1 can regulate the promoter activity (Horton., 2002). Srebp-1 stimulates the expression of Δ6 Fad in pink perch and rainbow trout (Dong., 2017); stimulates the expressions of Δ6 Fad, Elovl4, Elovl5 in the large yellow croaker (Li., 2017); and the expres- sions of Δ6/Δ5 Fad, Elovl5 in rabbitfish (Zhang., 2016)binding SREs in the promoter region. Lxrα can direct- ly or indirectly affect the synthesis of LC-PUFAbind- ing to liver X responsive elements (Lxres) in the promoter region or through regulating the expression of some specific transcription factors like Srebp-1 (Cha and Repa, 2007; Postic., 2007).It has been documented in studies in Atlantic salmon (Carmona-Anto., 2014), large yel- low croaker (Li., 2017) and rabbitfish (Zhang., 2016).
In conclusion, we report the cloning, functional charac- terization and transcriptional regulation ofgene in the large yellow croaker. The deduced protein exhibits all the structural features of Elovl family and shares high ho- mology with Elovl4a of other fishes. Functional characteri- zation reveals thatElovl4acan effectively elon- gate both C18 and C20 PUFA substrates. In addition, tis- sue distribution indicates that it is expressed predominant- ly in brain. Moreover, the transcription factors Hnf4α, Lxrα and Pparα all have significant positive effect onreporter activity. These results enrich the know- ledge on biosynthesis pathway of LC-PUFA in the large yellow croaker and lay a foundation for further understand- ing the anabolism mechanism of LC-PUFA in fish.
Acknowledgements
This work was supported by the Natural Science Foun- dation of Fujian Province (No. 2018J01450), the National Natural Science Foundation of China (No. U1705231), the National Marine Fisheries Industrial Technology System Post Scientist Project (No. CARS-47-G04), the Scientific Research Foundation of Jimei University (No. ZQ2019046), the Central Public-Interest Scientific Institution Basal Re- search & Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture, P. R. China, CAFS(No. 2017HY-XKQ01). The authors declare that they have no competing financial interests or personal relationships that can influence this study.
Agaba, M., Tocher, D. R., Dickson, C. A., Dick, J. R., and Teale, A. J., 2004. Zebrafish cDNA encoding multifunctional fatty acid elongase involved in production of eicosapentaenoic (20: 5n-3) and docosahexaenoic (22:6n-3) acids.,6 (3): 251-261.
Agaba, M. K., Tocher, D. R., Zheng, X., Dickson, C. A., Dick, J. R., and Teale, A. J., 2005. Cloning and functional characterisation of polyunsaturated fatty acid elongases of marine and freshwater teleost fish., 142 (3): 342-352.
Agbaga, M. P., Mandal, M. N. A., and Anderson, R. E., 2010. Retinal very long chain polyunsaturated fatty acids: New insights from studies on ELOVL4 protein., 51 (7): 1624-1642.
Andersen, I., Eijsink, V. G. H., and Thomassen, M., 2000. Mul- tiple variants of the peroxisome proliferator-activated recep- tor (PPAR) γ are expressed in the liver of Atlantic salmon ()., 255 (2): 411-418.
Bell, M. V., and Dick, J. R., 2005. Distribution of 22:6n-3 new- ly synthesized from 18:3n-3 into glycerolipid classes from tis- sues of rainbow trout ()., 40 (7): 703-708.
Bell, M. V., and Tocher, D. R., 2009. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In:. Arts, M. T.,., eds., Springer, New York, 377pp.
Betancor, M. B., Oboh, A., Ortega, A., Mourente, G., Navarro, J. C., de la Gándara, F,, 2020. Molecular and functional characterization of a putative elovl4 gene and its expression in response to dietary fatty acid profile in Atlantic bluefin tuna()., 240: 110372- 110383.
Calder, P. C., 2012. Mechanisms of action of (n-3) fatty acids., 142 (3): 592S-599S.
Carmona-Antoñanzas, G., Monroig, Ó., Dick, J. R., Davie, A., and Tocher, D. R., 2011. Biosynthesis of very long-chain fat- ty acids (C ≥ 26) in Atlantic salmon: Cloning, functional cha- racterisation, and tissue distribution of an Elovl4., 159 (2): 122-129.
Carmona-Antoñanzas, G., Tocher, D. R., Martinez-Rubio, L., and Leaver, M. J., 2014. Conservation of lipid metabolic gene trans-criptional regulatory networks in fish and mammals.,534 (1): 1-9.
Castro, L. F. C., Tocher, D. R., and Monroig, Ó., 2016. Long- chain polyunsaturated fatty acid biosynthesis in chordates: In- sights into the evolution of Fads and Elovl gene repertoire., 62: 25-40.
Cha, J. Y., and Repa, J. J., 2007. The liver X receptor (LXR) and hepatic lipogenesis: The carbohydrate-response element-bind- ing protein is a target gene of LXR., 282 (1): 743-751.
Cinti, D. L., Cook, L., Nagi, M. N., and Suneja, S. K., 1992. The fatty acid chain elongation system of mammalian endoplas- mic reticulum., 31 (1): 1-51.
Cook, H. W., and Mcmaster, C. R., 2002. Fatty acid desatura- tion and chain elongation in eukaryotes. In:,.Elsevier B.V., 181-204.
Das, U. N., 2003. Long-chain polyunsaturated fatty acids in the growth and development of the brain and memory., 19 (1): 62-65.
Dong, X., Tan, P., Cai, Z., Xu, H., Li, J., Ren, W.,., 2017. Regulation of FADS2 transcription by SREBP-1 and PPAR-α influences LC-PUFA biosynthesis in fish., 7: 40019-40024.
Dong, Y., Zhao, J., Chen, J., Wang, S., and Li, Y., 2018. Cloning and characterization of ∆6/∆5 fatty acyl desaturase (Fad) gene promoter in the marine teleost., 647: 174-180.
Fernandezpalacios, H., Izquierdo, M., Robaina, L., Valencia, A., Salhi, M., and Montero, D., 1997. The effect of dietary protein and lipid from squid and fish meals on egg quality of broodstock for gilthead seabream ()., 148: 233-246.
Folch, J. M. S., Lees, M. M., and Stanley, G. H. S., 1957. A simple method for the isolation and purification of total lipids from animal tissues., 22 (1): 24-36.
Gatlin III, D. M., 2003. Nutrition and fish health. In:. Halver, J. E., and Hardy, R. W., eds., Academic Press, San Diego, California, 671-702.
Ghioni, C., 1999. Low C18 to C20 fatty acid elongase activity and limited conversion of stearidonic acid, 18:4n-3, to eico- sapentaenoic acid, 20:5n-3, in a cell line from the turbot,.–, 1437 (2): 170-181.
Gregory, M. K., and James, M. J., 2014. Rainbow trout () Elovl5 and Elovl2 differ in selectivity for elongation of omega-3 docosapentaenoic acid.–, 1841 (12): 1656-1660.
Guillou, H., Zadravec, D., Martin, P. G. P., and Jacobsson, A., 2010. The key roles of elongases and desaturases in mamma- lian fatty acid metabolism: Insights from transgenic mice., 49 (2): 186-199.
Halver, J. E., and Hardy, R. W., 2003. Nutrient flow and reten- tion. In:. Academic Press, San Diego, Califor- nia, 755-770.
Hastings, N., Agaba, M., Tocher, D. R., Leaver, M. J., Dick, J. R., Sargent, J. R.,., 2002. A vertebrate fatty acid desatu- rase with Δ5 and Δ6 activities., 98 (25): 14304-14309.
Horton, J. D., Goldstein, J. L., and Brown, M. S., 2002. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver., 109: 1125-1131.
Huang, S. S. Y., Oo, A. N., Higgs, D. A., Brauner, C. J., and Sa- toh, S., 2007. Effect of dietary canola oil level on the growth performance and fatty acid composition of juvenile red sea bream,., 271 (1-4): 420-431.
Ibabe, A., Grabenbauer, M., and Baumgart, E., 2002. Expressionof peroxisome proliferator-activated receptors in zebrafish ()., 118 (3): 231-239.
Ikeda, M., Kanao, Y., Yamanaka, M., Sakuraba, H., Mizutani, Y., Igarashi, Y.,., 2008. Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis., 582 (16): 2435-2440.
Jakobsson, A., Westerberg, R., and Jacobsson, A., 2006. Fatty acid elongases in mammals: Their regulation and roles in metabo- lism., 45 (3): 237-249.
Jaya-Ram, A., Kuah, M., Lim, P., Kolkovski, S., and Shu-Chien, A. C., 2008. Influence of dietary HUFA levels on reproduc- tive performance, tissue fatty acid profile and desaturase and elongase mRNAs expression in female zebrafish., 277 (3): 275-281.
Kabeya, N., Yamamoto, Y., Cummins, S. F., Elizur, A., Yazawa, R., Takeuchi, Y.,., 2015. Polyunsaturated fatty acid metabolism in a marine teleost, Nibe croaker: Functional characterization of Fads2 desaturase and Elovl5 and Elovl4 elongases., 188: 37- 45.
Kang, J. X., and Leaf, A., 1996. The cardiac antiarrhythmic ef- fects of polyunsaturated fatty acid., 31: 41-44.
Kondo, H., Misaki, R., Gelman, L., and Watabe, S., 2007. Li- gand-dependent transcriptional activities of four torafugu puf- ferfishperoxisome proliferator-activated re- ceptors., 154 (1-3): 120- 127.
Lagali, P. S., Liu, J., Ambasudhan, R., Kakuk, L. E., and Ayya- gari, R., 2003. Evolutionarily conserved ELOVL4 gene ex- pression in the vertebrate retina., 44 (7): 2841-2850.
Lauritzen, L., Hansen, H. S., Jorgensen, M. H., and Michaelsen, K. F., 2001. The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina., 40 (1-2): 1-94.
Leaver, M. J., Evridiki, B., Efthimia, A., Amalia, D., Laurence, F. K., Tariq, E. M.,., 2005. Three peroxisome proliferator- activated receptor isotypes from each of two species of ma- rine fish., 146 (7): 3150-3162.
Leaver, M. J., Ezaz, M. T., Fontagne, S., Tocher, D. R., Boukou-vala, E., and Krey, G., 2007. Multiple peroxisome proliferator- activated receptor beta subtypes from Atlantic salmon ()., 38 (3): 391-400.
Lee, S. M., and Cho, S. H., 2008. Influences of dietary fatty acid profile on growth, body composition and blood chemistry in juvenile fat cod (Jordan et Starks)., 15 (1): 19-28.
Leonard, A. E., Pereira, S. L., Sprecher, H., and Huang, Y. S., 2004. Elongation of long-chain fatty acids., 43 (1): 36-54.
Li, S., Monroig, Ó., Navarro, J. C., Yuan, Y., Xu, W., Mai, K.,., 2017. Molecular cloning and functional characterization of a putative Elovl4 gene and its expression in response to dietary fatty acid profiles in orange-spotted grouper., 48 (2): 537-552.
Li, S., Monroig, Ó., Wang, T., Yuan, Y., Carlos Navarro, J., Hon-toria, F.,., 2017. Functional characterization and differen- tial nutritional regulation of putative Elovl5 and Elovl4 elongases in large yellow croaker ()., 7 (1): 2289-2303.
Li, Y., Monroig, Ó., Zhang, L., Wang, S., Zheng, X., Dick, J. R.,., 2010. Vertebrate fatty acyl desaturase with Δ4 activity., 107 (39): 16840-16845.
Livak, K. J., and Schmittgen, T. D., 2002. Analysis of relative gene expression data using real-time quantitative PCR., 25 (4): 402-408.
Marsh, D., 2008. Protein modulation of lipids, and vice-versa, in membranes.–, 1778 (7): 1545-1575.
Meyer, A., 2004. Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis., 45 (10): 1899-1909.
Miyazaki, M., and Ntambi, J. M., 2008. Fatty acid desaturation and chain elongation in mammals. In:5th edition, Vance, D. E., and Vance, J. E., eds.,Elsevier Science, 191-211.
Monroig, Ó., Wang, S., Zhang, L., You, C., Tocher, D. R., and Li, Y., 2012. Elongation of long-chain fatty acids in rabbitfish: Cloning, functional characterisation and tissue distribution of Elovl5- and Elovl4-like elongases., 350-353: 63-70.
Monroig, Ó., Webb, K., Ibarracastro, L., Holt, G. J., and Tocher, D. R., 2011. Biosynthesis of long-chain polyunsaturated fatty acids in marine fish: Characterization of an Elovl4-like elongase from cobiaand activation of the path- way during early life stages., 312: 145-153.
Moon, A. Y., 2003. Identification of two mammalian reductases involved in the two-carbon fatty acyl elongation cascade., 278 (9): 7335-7343.
Morais, S., Monroig, Ó., Zheng, X., Leaver, M. J., and Tocher, D. R., 2009. Highly unsaturated fatty acid synthesis in Atlantic salmon: Characterization of Elovl5- and Elovl2-like elongases., 11 (5): 627-639.
Mourente, G., Díaz-Salvago, E., Tocher, D. R., and Bell, J. G., 2000. Effects of dietary polyunsaturated fatty acid/vitamin E (PUFA/ tocopherol) ratio on antioxidant defence mechanisms of juvenile gilthead sea bream (L., Osteichth- yes, Sparidae)., 23 (4): 337- 351.
Navarro-Guillén, C., Engrola, S., Castanheira, F., Bandarra, N., Hachero-Cruzado, I.,., 2013. Interaction between dietary levels of LC-PUFA and vegetable oil sources in Senegalese sole () post-larvae: Puzzling results suggesting complete biosynthesis pathway from C18 PUFA to DHA., 78 (4): 300-303.
Nord, Y. A., Marchioli, R., Arnesen, H., and Videb, K. J. R., 2001. n-3 Polyunsaturated fatty acids and cardiovascular diseases: To whom, how much, preparations., 36 (1 supplement): S127-S129.
Oboh, A., Navarro, J. C., Tocher, D. R., and Monroig, Ó., 2017. Elongation of very long-chain (>C(24)) fatty acids in: Cloning, functional characterization and tissue ex- pression of elovl4 elongases.,52 (10): 837-848.
Oscar, M., Josep, R., José, M. C., James, R. D., Antonio, F., and Douglas, R. T., 2010. Expression and role of Elovl4 elongases in biosynthesis of very long-chain fatty acids during zebrafishearly embryonic development.–, 1801 (10): 1145-1154.
Osei, P., Suneja, S. K., Laguna, J. C., Nagi, M. N., Cook, L., Prasad, M. R.,., 1989. Topography of rat hepatic microsomal enzymatic components of the fatty acid chain elongation system., 264 (12): 6844- 6849.
Park, W. J., Kothapalli, K. S. D., Lawrence, P., Tyburczy, C., and Brenna, J. T., 2009. An alternate pathway to long-chain polyunsaturates: The FADS2 gene product delta8-desaturates 20:2n-6 and 20:3n-3., 50 (6): 1195- 2020.
Postic, C., Dentin, R., Denechaud, P. D., and Girard, J., 2007. ChREBP, a transcriptional regulator of glucose and lipid metabolism., 27 (1): 179-192.
Sargent, J. R., and Tacon, A. G. J., 1999. Development of farm- ed fish: A nutritionally necessary alternative to meat., 58 (02): 377-383.
Sargent, J. R., Tocher, D. R., and Bell, J. G., 2003. The lipids. In:. Halver, J. E., and Hardy, R. W., eds., Academic Press, San Diego, California, 181-257.
Stubhaug, I., Tocher, D. R., Bell, J. G., Dick, J. R., and Torstensen, B. E., 2005. Fatty acid metabolism in Atlantic sal- mon (L.) hepatocytes and influence of dietary vegetable oil.–, 1734 (3): 277-288.
Stulnig, T. M., and Zeyda, M., 2004. Immunomodulation by poly- unsaturated fatty acids: Impact on T-cell signaling., 39 (12): 1171-1175.
Tan, J. L., and Zon, L. I., 2011. Chemical screening in zebrafish for novel biological and therapeutic discovery., 105: 493-516.
Tocher,D. R., and Ghioni, C., 1999. Fatty acid metabolism in marine fish: Low activity of fatty acyl delta5 desaturation in gilthead sea bream () cells., 34 (5): 433- 440.
Tocher, D. R., Fonseca-Madrigal, J., Bell, J. G., Dick, J. R., and Sargent, J. R., 2002. Effects of diets containing linseed oil on fatty acid desaturation and oxidation in hepatocytes and intestinal enterocytes in Atlantic salmon ()., 26 (2): 157-170.
Tocher, D. R., Leaver, M. J., and Hodgson, P. A., 1998. Recent advances in the biochemistry and molecular biology of fatty acyl desaturases., 37 (2-3): 73- 117.
Vladimir, D., and Jonathan, S. W., 2007. A molecular caliper me-chanism for determining very long-chain fatty acid length.,130 (4): 663-677.
Wang, S., Chen, J., Jiang, D., Zhang, Q., You, C., Tocher, D. R.,., 2018. Hnf4α is involved in the regulation of vertebrate LC-PUFA biosynthesis: Insights into the regulatory role of Hnf4α on expression of liver fatty acyl desaturases in the marine teleost., 44: 805-815.
Xiao, Y. F., Ke, Q., and Wang, S. Y., 2001. Single point muta- tions affect fatty acid block of human myocardial sodium chan- nel α subunit Na+channels., 98 (6): 3606- 3611.
Xie, D., Chen, F., Lin, S., You, C., Wang, S., Zhang, Q.,., 2016. Long-chain polyunsaturated fatty acid biosynthesis in the euryhaline herbivorous teleost: Functional characterization, tissue expression and nutritional re- gulation of two fatty acyl elongases., 198: 37-45.
You, C., Jiang, D., Zhang, Q., Xie, D., Wang, S., Dong, Y.,., 2017. Cloning and expression characterization of peroxisome proliferator-activated receptors (PPARs) with their agonists, dietary lipids, and ambient salinity in rabbitfish., 206: 54-64.
Zakeri, M., Kochanian, P., Marammazi, J. G., Yavari, V., Savari, A., and Haghi, M., 2011. Effects of dietary n-3 HUFA concentrations on spawning performance and fatty acids composition of broodstock, eggs and larvae in yellowfin sea bream,., 310: 388-394.
Zhang, Q., You, C., Liu, F., Zhu, W., Wang, S., Xie, D.,., 2016. Cloning and characterization of Lxr and Srebp1, and their potential roles in regulation of LC-PUFA biosynthesis in rabbitfish., 51: 1051-1063.
Zheng, X., Ding, Z., Xu, Y., Monroig, Ó., Morais, S., and Tocher, D. R., 2009. Physiological roles of fatty acyl desaturases and elongases in marine fish: Characterisation of cDNAs of fatty acyl delta6 desaturase and elovl5 elongase of cobia ()., 290 (1-2): 122-131.
Zheng, X., Leaver, M. J., and Tocher, D. R., 2009. Long-chain polyunsaturated fatty acid synthesis in fish: Comparative ana- lysis of Atlantic salmon (L.) and Atlantic cod (L.) Δ6 fatty acyl desaturase gene promoters., 154 (3): 255-263.
Zheng, X., Torstensen, B. E., Tocher, D. R., Dick, J. R., and Bell, J. G., 2005. Environmental and dietary influences on highly unsaturated fatty acid biosynthesis and expression of fatty acyl desaturase and elongase genes in liver of Atlantic salmon ().–, 1734 (1): 13-24.
September 17, 2020;
February3, 2021;
April 20, 2021
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2021
#The two authors contributed equally to this work.
E-mail: li.wanbo@jmu.edu.cn
E-mail: zywang@jmu.edu.cn
(Edited by Qiu Yantao)
杂志排行

Journal of Ocean University of China的其它文章
- Meshless Method with Domain Decomposition for Submerged Porous Breakwaters in Waves
- Facial Features of an Air Gun Array Wavelet in the Time-Frequency Domain Based on Marine Vertical Cables
- Magma Evolution Processes in the Southern Okinawa Trough:Insights from Melt Inclusions
- Summery Intra-Tidal Variations of Suspended Sediment Transportation–Topographical Response and Dynamical Mechanism in the Aoshan Bay and Surrounding Area, Shandong Peninsula
- High-Resolution Geochemical Records in the Inner Shelf Mud Wedge of the East China Sea and Their Indication to the Holocene Monsoon Climatic Changes and Events
- Geological Guided Tomography Inversion Based on Fault Constraint and Its Application