Bmi-1基因在调节牙本质及牙槽骨形成和发育中的作用及机制研究
2021-12-20黄元清李斌魏敏肖婵刘丽
黄元清 李斌 魏敏 肖婵 刘丽
中图分类号:R780.2 文献标志码:A doi:10.3969/j.issn.1001-3733.2021.06.003
牙齿的形成是各种分子从起始到终末分化的精确协调过程,牙齿发育还包括一系列有序的诱导信号,对细胞增殖、分化和器官发生发挥全局控制作用[1-3]。下颌骨的发育始于下颌突第一鳃弓的出现,然后形成梅克尔软骨。除髁突之外,其余下颌骨均经历膜内成骨,成骨的一系列因素依赖于不同因素[4-5]。牙槽骨增大也是通过膜内成骨过程发生的,在此过程中,骨软骨祖细胞依次经历上皮-间充质诱导、增殖和分化[6]。目前,对出生后小鼠牙齿和下颌骨生长发育的分子基础了解甚少。
B细胞淋巴瘤滤过性病毒插入位点1 (B cell-specific moloney leukemia virus insert site-1,Bmi-1) 基因属于聚硫蛋白复合体基因家族(polycomb group,PcG)的成员之一,具有转录抑制剂的作用,对生物体发生过程中基因的正确表达起到关键的调控作用[7]。研究证实,Bmi-1基因缺失小鼠表现出包括骨质疏松[8]在内的全身过早衰老表型。Bmi-1对下游通路的调节主要通过两条通路来实现,一是通过介导的INK4a/Arf位点的降低P53水平完成对细胞周期分子机制的调控;其次是通过减少线粒体ROS水平的产生,从而间接地参与DNA损伤修复的作用。抗氧化剂在很大程度上纠正了Bmi-1缺陷小鼠引起的胸腺细胞和肾脏发育缺陷[9-10]。考虑到牙釉质生成、颌骨膜内成骨与长骨软骨成骨之间的差异,尚不清楚Bmi-1缺乏是否会通过扰乱氧化还原稳态和诱导牙槽骨DNA损伤而导致发育过程中的牙本质和牙槽骨缺损。
为了探讨Bmi-1在牙本质和牙槽骨发育中的潜在作用,取 3周龄Bmi-1基因敲除(BKO)小鼠15 只给予普通饮食4 周,取同窝野生型小鼠(WT)15 只进行对照,通过影像学、组织病理学、细胞培养和分子生物学的方法比较分析两组动物之间牙齿及牙槽骨相关指标之间差异。
1 材料与方法
1.1 实验动物
1.1.1 实验动物的繁殖及基因鉴定 将Bmi-1杂合子(Bmi-1+/-,BKO)小鼠(129Ola/FVB/N杂交背景)与C57BL/6J背景进行10~12 次回交,交配产生Bmi-1纯合子(Bmi-1-/-, BKO)及其野生型(WT)。小鼠饲养于南京医科大学实验动物中心。本研究严格按照南京医科大学实验动物研究所的指导方针进行。本研究的所有动物实验都经过了南京医科大学动物实验伦理委员会的批准(批准号: BK2006576)。
1.1.2 实验动物分组及饲养 取子代小鼠WT和BKO小鼠将分别给予以下不同处理:正常饮食(WT)组(15 只):子代小鼠3 周龄断奶后,普通饮食(含1%钙、0.67%磷)饲养WT小鼠4 周。正常饮食(BKO)组(15 只):子代小鼠3 周龄断奶后,普通饮食(含1%钙、0.67%磷)饲养BKO小鼠4 周。
1.2 实验方法
1.2.1 取材 取7 周龄WT、BKO小鼠,麻醉后立即行颈椎脱臼处死,取同侧牙及下颌骨,使用能更好保存酶活性和组织抗原性的甲醛混合物(2%副醛,75 mmol/L赖氨酸,10 mmol/L过碘酸钠,Paraldehyde,Lysine,Sodium periodate,PLP) 固定液固定,经常规脱水、石蜡包埋、切片,用于HE染色、组织化学染色和免疫组织化学染色。
1.2.2 X线摄影观察 固定后的牙及下颌骨使用Faxitron Model 805 radiographic检测系统(Faxitron Contact, Germany; 22 kV and 4-minute exposure time)进行X线摄影,观察骨骼形态和骨密度的改变。
1.2.3 Micro-CT扫描和三维重建 取固定后的牙及下颌骨,进行Micro-CT扫描(电压100 kV,电流98 mA,SKY SCAN 1072,Bruker公司,德国)和三维重建,选择经中切牙、第一磨牙、第二磨牙和第三磨牙断面进行比较分析观察。
1.2.4 HE染色 石蜡切片常规脱蜡水化,苏木精(Sigma公司,美国)染色,1%盐酸酒精分化,流水冲洗返蓝,伊红(Sigma公司,美国)复染,常规脱水透明,中性树胶封片。
1.2.5 组织化学染色 取下颌骨组织的石蜡切片,作总胶原蛋白(total collagen,TCOL)、碱性磷酸酶(akaline phosphatase,ALP)和抗酒石酸酸性磷酸酶(tartrate-resistant acid phosphates,TRAP)等组织化学染色。
1.2.6 免疫组织化学染色 取牙及下颌骨组织进行固定、脱水、石蜡包埋切片,作免疫组织化学染色,检测指标包括反映成骨和成牙本质细胞相关的指标:骨钙素(OCN),I型胶原蛋白(Col I),双糖链蛋白多糖(Biglycan),牙本质涎磷蛋白(dentin sialophosphoprotein,DSP)。
1.2.7 牙槽骨骨髓间充质干细胞的培养和细胞化学染色 取3 周龄下颌牙槽骨骨髓间充质干细胞(bone marrow stem cells,BMSCs)进行培养后,用PBS洗涤,加入冷乙醇固定。固定后,按顺序对碱性磷酸酶(ALP)阳性菌落、钙化菌落、胶原阳性菌落和总菌落进行染色。每个染色步骤结束后,对培养皿拍照、去渍、按要求留置。总成纤维细胞菌落(CFU-f)菌落的数量是用人工计算的。ALP染色阳性的菌落被认为是来自具有表达ALP能力的CFU-f,并被称为CFU-f ALP。
1.3 统计学处理
统计工具应用SPSS 14.0版软件包,计数资料采用χ2检验,计量资料采用t检验进行统计学分析(取双侧),P<0.05为差异有统计学意义。
2 结 果
2.1 Bmi-1对下颌骨牙槽骨矿化的影响
为了确定BKO小鼠是否会导致牙齿和下颌生长迟缓,通过X线摄影和Micro-CT断层摄影对两组小鼠牙齿和下颌生长的相关参数进行比较分析。结果显示,与WT小鼠相比,BKO小鼠牙齿和下颌骨的骨密度(BMD)较低(图1A)。中切牙、第一、第二、第三磨牙显微CT扫描切片显示,与WT小鼠相比,BKO小鼠的切牙和磨牙矿化程度以及下颌骨皮质骨和牙槽骨体积均减小(图1B)。
2.2 Bmi-1对牙齿、皮质骨和牙槽骨容量的影响
为确定Bmi-1缺失是否会导致牙齿、皮质骨和牙槽骨容量的减少,通过HE和TCOL染色分析BKO和同窝WT小鼠下颌骨的表型差异。如图2所示,与WT小鼠相比,BKO小鼠的牙本质(矿化和非矿化)厚度和牙槽骨体积均减小(图1C~D);BKO小鼠第一磨牙的牙本质厚度(图1E)、皮质骨厚度(图1F)和牙槽骨容量(图1G)均较WT组显著降低。
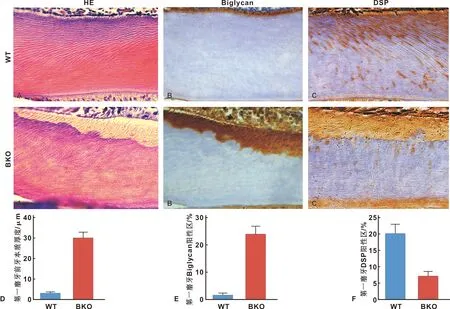
图2 Bmi-1对前期牙本质成熟和牙本质形成的影响(×400)
2.3 Bmi-1对前期牙本质成熟和牙本质形成的影响
HE染色切片显示,与同WT小鼠相比,BKO小鼠第一磨牙的前期牙本质面积占比明显增加(图2A、2D)。在前期牙本质区域检测到biglycan阳性免疫反应(图2B)。与WT小鼠相比,BKO小鼠第一磨牙中biglycan阳性区域占牙本质的比例明显增加(图2E)。在第一磨牙的前期牙本质和牙本质中检测到DSP阳性免疫反应(图2C)。与WT小鼠相比,BKO小鼠第一磨牙中DSP阳性区域显著减少(图2F)。
2.4 Bmi-1对牙槽骨成骨细胞骨形成的影响
为检测Bmi-1对牙槽骨成骨细胞骨形成的影响,采用HE染色、ALP染色、Col I和OCN免疫组化染色分析牙槽骨成骨细胞数量。如图3结果显示,与BKO小鼠相比,WT小鼠牙槽骨中成骨细胞、成骨细胞数量区域、ALP、Col I和OCN阳性区域显著增加。
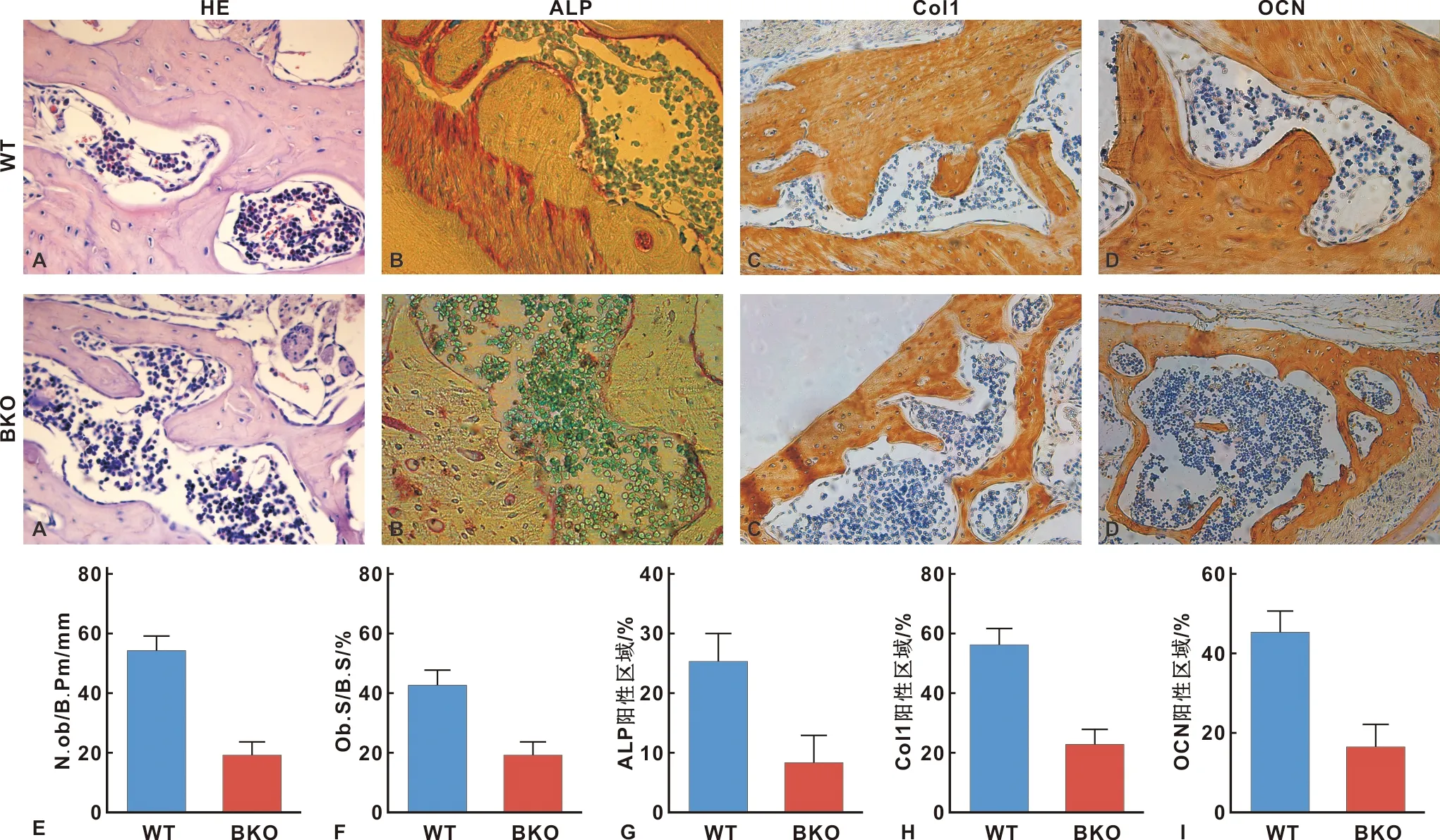
图3 Bmi-1对成骨细胞骨形成的影响(×400)
2.5 Bmi-1对牙槽骨破骨细胞骨吸收的影响
为了测定Bmi-1对牙槽骨破骨细胞吸收的影响,采用TRAP染色法分析牙槽骨破骨细胞的数量。如图4结果显示,与BKO小鼠相比,WT小鼠牙槽骨中破骨细胞阳性细胞数量、破骨细胞阳性细胞区明显减少。上述结果表明,Bmi-1降低了破骨细胞骨吸收能力。
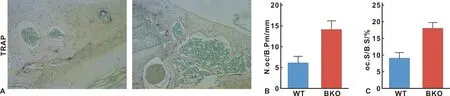
图4 Bmi-1对成骨细胞骨形成和破骨细胞骨吸收的影响(TRAP, ×200)
2.6 Bmi-1对CFU-F形成效率及牙槽骨BMSC分化的影响
为了阐明所观察到的成骨细胞数量减少是否由于BKO小鼠成骨细胞功能缺陷,对3 周龄的BKO小鼠及其同窝WT小鼠进行骨髓培养。与WT小鼠相比,BKO培养产生CFU-Fs和ALP阳性CFU-fap菌落的能力明显受损(图5)。
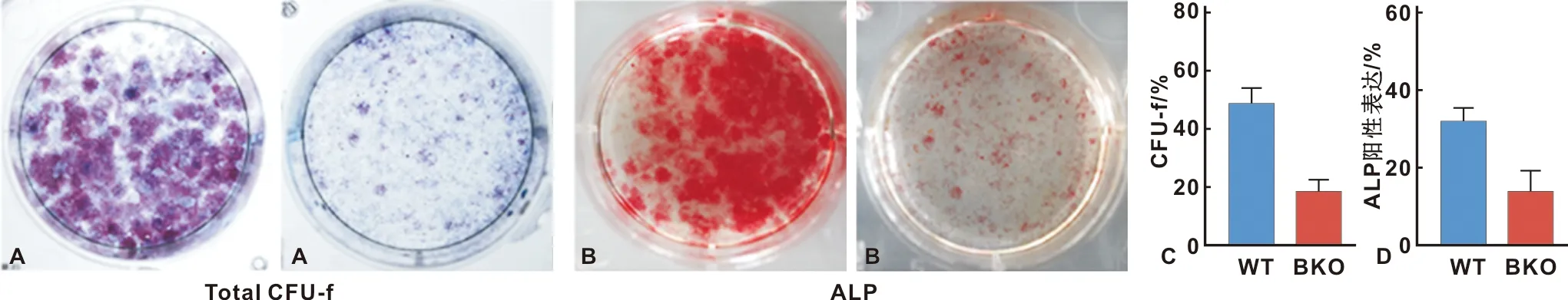
图5 Bmi-1对CFU-F形成效率及牙槽骨BMSC分化的影响
3 讨 论
Bmi-1来源于多疏基因家族,具有抑制Hox基因在生长发育中的空间特异性和时间特异性表达的作用。Bmi-1基因缺失导致寿命缩短,生长迟缓[11]。Zhang等[12]研究表明,Bmi-1基因敲除小鼠可诱导衰老相关骨质疏松症,主要是通过下调骨髓间充质干细胞的自我更新。本研究通过对Bmi-1缺陷小鼠牙及下颌骨的表型分析发现,牙及下颌骨的骨密度、矿化皮质骨和牙槽骨体积、成骨细胞数量、成骨细胞ALP活性的表达均降低。前期研究表明抗氧化剂吡咯喹啉醌(PQQ)可以显著改善Bmi-1缺失小鼠的长骨缺损[8]以及抵抗腮腺辐射损伤的作用[12]。因此,这些结果表明,Bmi-1基因的缺失干扰了长骨和下颌骨成骨细胞的骨形成作用。
牙齿和下颌骨的成骨方式属于膜内成骨,长骨的成骨方式属于软骨成骨。鉴于此,提出Bmi-1是否也在抵抗由Bmi-1缺失引起的牙齿和下颌骨缺损中发挥重要作用的假说。本研究结果显示,与同窝WT小鼠相比,Bmi-1缺失小鼠的第一磨牙前期牙本质以及biglycan占比均显著增加。DSP在形成牙本质的胶原基质中的分布表明该蛋白在调节矿物沉积方面发挥着重要作用。结果还显示,Bmi-1缺失小鼠的DSP免疫阳性区域减少,这与牙本质厚度减少一致。因此,上述研究结果表明,Bmi-1基因缺陷会损害牙本质的形成和矿化,因此,Bmi-1可以促进牙本质的形成和矿化。
本研究还发现BKO小鼠CFU-F形成效率明显降低,牙槽骨BMSC的增殖减少,传代的牙槽骨BMSCs在Bmi-1缺陷小鼠细胞培养中无法增殖。以上结果表明,Bmi-1在牙槽骨骨髓间充质干细胞高效自我更新中也起着重要作用。在之前的[13]研究中,在明确的体内外条件下,一定比例的CFU-F可形成多种间充质组织,包括骨、脂肪组织、软骨、骨髓支持间质、平滑肌、心肌细胞和肌腱。因此,BMSC自我更新的缺陷不仅可能导致骨质疏松表型,而且可能导致多组织生长迟缓。
本研究结果发现Bmi-1基因敲除小鼠诱导了下颌骨骨质疏松以及牙齿发育障碍,并证实了Bmi-1基因主要通过增加下颌骨成骨细胞骨形成、减少破骨细胞骨吸收、促进前期牙本质发育成熟等途径发挥保护作用。为下一步深入Bmi-1基因调节骨微环境机制研究提供一定的理论和实验依据。
