运动神经元的模型研究方法
2021-12-18徐伟航
徐伟航
前言
行进运动(Locomotion)是指简单重复的机械运动并产生位移,是脊椎动物运动的基本形式[1],因此研究行进运动对于运动康复、智能行走等领域的研究有着非常重要的意义[2]。刺激位于脑干中的中脑运动区域(Midbrain Locomotor Region,MLR)可以产生神经信号,经过脊髓中的中枢模式发生器(Central Pattern Generator,CPG)调节,可引发肢体产生左右交替的节律性行进运动,其中运动神经元作为直接控制肌肉的最终神经元在脊椎动物行进运动中扮演着重要的角色。对于运动神经元的研究一直在进行,1992年,Brownstone等人在去除大脑的猫中通过刺激脑干MLR区域产生神经信号来控制猫在跑步机上产生行进运动,同时借助膜片钳技术来研究行进运动中猫运动神经元的兴奋性变化[3];2004年,Gazula等人研究了运动对于脊髓损伤(Spinal Cord Injury,SCI)大鼠运动神经元位置以及树突形态的影响,发现了运动可以使脊髓损伤大鼠运动神经元树突变得更加丰富[4]。通过实验研究运动神经元得出了许多非常重要的结论,但受限于生理实验的高成本且操作难度大等局限性,很多生理实验无法进行,因此,模型研究运动神经元受到越来越多科研人员的青睐。2009年,Kurian等人使用Matlab软件建立了大鼠运动神经元的双室(胞体、树突)模型研究了细胞形态对于运动神经元放电模式的影响,为脊髓损伤的病理研究提供理论支持[5];2018年,戴跃等人通过Genisis软件建立了猫的运动神经元模型,给出了离子通道主导的主动膜特性与运动神经元兴奋性之间的数学关系[6]。本文综述了前人的研究成果,总结了当前运动神经元的模型研究方法,希望为相关的研究提供参考依据。
1 运动神经元生理学背景
1.1 运动神经元简介
脊椎动物的运动神经元多集中分布在脊髓前角,轴突伸出直接与骨骼肌相连来支配肌肉纤维[7],这样的结构决定了运动神经元是联系中枢神经系统、外周神经系统以及骨骼肌之间的桥梁[1][7]。运动神经元由胞体、轴突以及树突构成,表面为磷脂双分子层及少量蛋白质,内部为细胞核与细胞质等物质。根据支配肌肉类型的不同,运动神经元可分为α运动神经元和γ运动神经元两种类型,其中α运动神经元控制梭外肌,γ运动神经元控制梭内肌。在行进运动过程中,上游神经信号经过脊髓中间神经元传导进而引发运动神经元产生兴奋,再由神经元轴突末梢释放神经递质引起肌肉收缩,过程涉及到电信号到化学信号的转变。
1.2 离子通道
兴奋在运动神经元上的表现形式是动作电位,由离子通道调节,离子通道是内嵌在磷脂双分子层中的蛋白质结构,主要负责运输细胞内外的带电离子,其中钠(Na+)、钾(K+)、氯(Cl-)三种离子对动作电位的产生起主要作用。以大鼠的运动神经元为例,当细胞膜处于静息状态时,细胞外液中的Na+、Cl-浓度大于细胞内液,而K+的浓度小于细胞内液,造成了静息状态下细胞膜的电势为-55mV左右,当离子通道开启时,Na+内流,K+外流导致膜电压去极化形成动作电位,而当动作电位结束后,离子通道关闭,钠钾泵将多余的Na+和K+反向搬运,膜电位恢复至静息状态。离子通道分为电压门控型和配体门控型两种类型,电压门控型离子通道的开启和关闭主要受膜内外电压差的影响,比如钠离子通道和钾离子通道,而配体门控型需要特定类型的离子与受体结合才能开启,比如钙依赖性的钾离子通道。
2 运动神经元建模方法
2.1 舱体模型
在电流钳实验中,向神经元胞体上施加一个适当大小的电流刺激,可以观察到动作电位的产生,而建模的本质就是拟合出神经元电流与电压之间的数学表达式来模拟这一过程。运动神经元建模的第一步是细胞被动膜特性的数学模型化,包括膜电容、膜电阻、轴向电阻等神经元形态参数。细胞膜的磷脂双分子层结构在电流经过时类似于一个电容,被称为膜电容,造成的阻力被称为膜电阻,而细胞内部物质造成的阻力被称为轴向电阻,舱体模型使用Cm表示膜电容,Rm表示膜电阻,这样就构成了一个RC回路,类似于一个“舱体”,相邻舱体之间用一个耦合电阻Ra连接,用来代表轴向电阻,图一A表示一个原始的神经元,图一B是将神经元简化为多段圆柱体组成的“电缆”,再将每段电缆用舱体模型代替,最终模型化结果如图一C所示。

图一 舱体模型化过程
2.2 3/2等效定理
舱体模型给出了神经元被动膜特性的模型化方法,但考虑到运动神经元树突的数量庞大且拓扑结构复杂,如果直接使用舱体模型拟合运动神经元会产生数量庞大的RC回路,导致计算的时间复杂度过大,因此需要对模型进行简化,Rall等人在1959年提出了3/2等效定理,定理指出:将树突视为柱体,只要两个树突分支的直径d1和d2满足方程(1),就可以用一个直径为dp的树突代替它。通过3/2等效定理,可以对运动神经元的树突结构进行简化,图二A为简化前的运动神经元,图二B为使用3/2等效定理简化后的运动神经元,利用等效定理甚至可以将整个运动神经元用单个“柱体”表示。

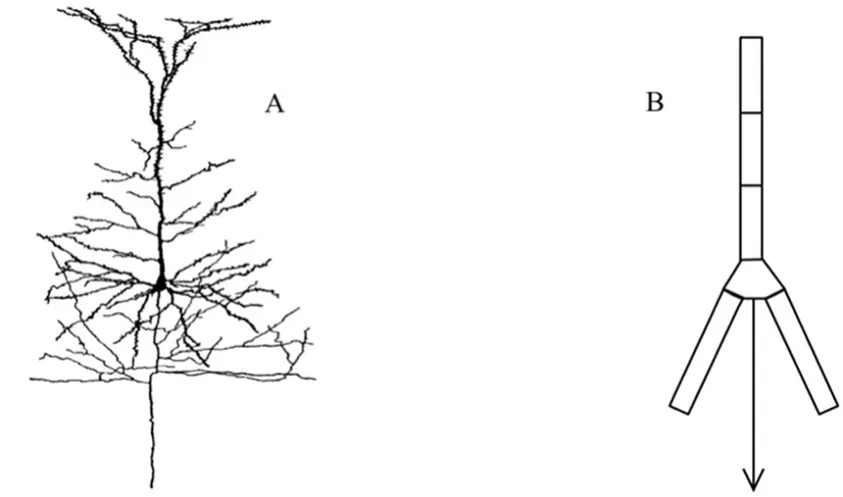
图二 3/2等效定理简化运动神经元
2.3 Hudgkin-Huxley模型
舱体模型对运动神经元的被动膜特性进行了数学建模,3/2等效定理对模型进行了简化,但动作电位的产生还依赖于离子通道所决定的主动膜特性,而主动膜特性建模则由两位诺贝尔奖获得者Hudgkin和Huxley在1952年提出,即著名的Hudgkin-Huxley模型(H-H模型)[2]。Hudgkin和Huxley在简陋的条件下利用枪乌贼实验拟合出了三种重要离子通道Na+、K+、Cl-的欧姆定律等效方程,如表1所示,其中Vm表示膜电压,iNa、iK、iL表示离子通道的电流,ENa、EK、EL为离子通道的平衡电位,GNa、GK、GL则表示离子通道的可变电导,而对于三个可变电导,又可分别用gNam3h、gKn4、gL表示,其中m表示Na+的激活概率,h表示Na+的失活概率,n表示K+的激活概率,取值范围都在0到1之间,gNa,gK,gL是定值,表示离子通道的最大电导,图三是根据等效方程并结合舱体模型建立的等效电路,公式(2)是根据基尔霍夫电流定律得出的等效电路的最终表达式,基于此数学表达式可以计算出神经元在电流刺激下膜电压的变化曲线,进而模拟动作电位的产生。

表1 Hodgkin-Huxley等效方程
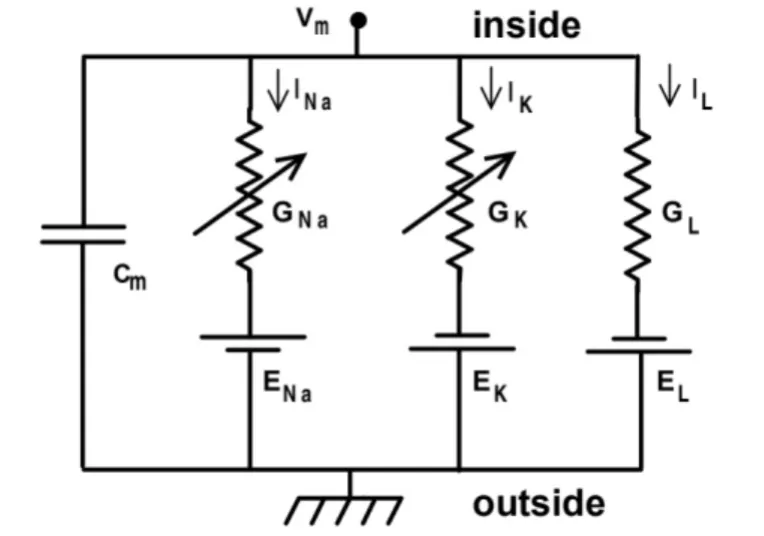
图三 Hudgkin-Huxley模型
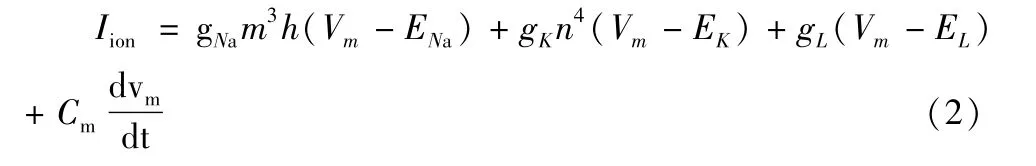
2.4 模型可用性分析
基于以上理论,可以建立运动神经元的数学模型,但在进行建模任务时,还需要考虑模型是否具有生理学意义,不同类型动物的运动神经元参数之间差异巨大,比如猫的运动神经元树突长度和膜电阻分别为2000~3000um和7000~10000Ωcm2,而大鼠的多为600~1500um和500~1000Ωcm2,这就要求研究人员在搭建模型的时候尽可能地拟合实验对象的生理参数,既包括运动神经元各部分的长度、直径、膜电容、膜电阻、轴向电阻等被动膜特性,还需要调整离子通道的主动膜特性参数使动作电位的各项指标(电压阈值、动作电位幅度和宽度、后超极化电位深度和宽度等)处于合理的范围之内。
3 结语
文章主要综述运动神经元的模型研究方法,首先介绍了运动神经元的生理学背景,包括运动神经元的结构与功能、离子通道的作用,接着从被动膜特性和主动膜特性两个方面介绍了运动神经元模型化的方式,包括舱体模型、3/2等效定理以及Hudgkin-Huxley模型,最后分析了实际建模过程中需要注意的细节。模型研究运动神经元克服了生理实验研究的很多障碍,既可以用于验证实验研究的结论,又可以在无法进行生理实验时,通过模型研究给出预测。
