伊犁马群体MSTN 速度基因多样性研究
2021-12-17阿拉达尔胡建伟李蒙蒙高雪丽郑文祥姚新奎
齐·阿拉达尔,胡建伟,李蒙蒙,高雪丽,郑文祥,姚新奎,3
(1.新疆农业大学动物科学学院,新疆乌鲁木齐 830052;2.新疆维吾尔自治区地方国营伊犁种马场,新疆昭苏县 835612;3.新疆马繁育与运动生理重点实验室,新疆乌鲁木齐 830052)
环境和训练是影响马速度能力的重要因素,近年的研究也揭示了遗传因素对马运动能力的影响。英纯血(Thoroughbred)也称为纯血马,起源于英国,是专门为速度赛而培育的马品种,经历了三百多年的封闭式人工选育,目前每200 米奔跑用时达10~12 s,可称为世界上速度最快的马品种。美国的夸特马(Quarter horse)、标准品种(Standardbred)以及欧洲的一些温血马品种的培育过程中都有纯血马血统导入。研究表明,纯血马的短距离爆发力和最适比赛距离与马的抑肌素基因(MSTN)有一定的相关性[1-3]。MSTN基因第一内含子中的g.66493737 T>C 突变可以判断纯血马最适比赛距离,携带CC 型纯合突变的马更适合短距离爆发力的比赛,而携带TT 型的纯合子个体更容易在中长距离比赛中获胜[4]。因此,MSTN基因的g.66493737 T>C 突变被认为是“速度基因”标记[5]。最近研究发现,在纯血马MSTN启动子上的一个227 bp 的短散在核原件(Short Interspersed Nuclear Elements,SINE)的插入影响了MSTN基因的转录效率和肌纤维组成,是真正引起“速度基因”效应的变异[5-6]。在纯血马群体中SINE 插入与C 变异呈高度连锁,但在一些非速度型品种中C 突变频率不高,也并不与SINE插入连锁[11]。
伊犁马分布于新疆伊犁州直属的昭苏、特克斯、尼勒克、新源、巩留等县境内,是从20 世纪初开始以哈萨克马为母本,与引进的前苏联的奥尔洛夫、顿河、布琼尼、阿哈尔捷金马等轻型种公马进行杂交改良而培育的骑乘型马品种[7]。随着近年新疆马产业的转型发展,人们再次尝试用不同形式的杂交改良和选育将伊犁马向速度型、速步型、耐力型、乳用型、肉用型和骑乘型等多个方向培育。21 世纪初,新疆伊犁种马场采取了伊犁母马与英纯血种公马杂交的形式,将伊犁马的培育方向定位为速度方向的培育,群体目前以级进杂交二代为主。伊犁种马场的速度型伊犁马经过二十年的培育,体尺和速度能力都得到了进一步改善。本研究旨在通过比较原始伊犁马群体和培育的速度型伊犁马群体中“速度基因”标记——MSTNg.66493737T>C 突变和SINE 插入突变的分布频率和连锁关系,为速度型伊犁马的进一步选育工作提供分子遗传学依据。
1 材料与方法
1.1 实验材料 本实验研究的两组群体分别是原始伊犁马和速度型伊犁马。原始伊犁马是指上个世纪以哈萨克马为母本,与引进的前苏联马品种进行杂交并经过横交固定的伊犁马品种。速度型伊犁马是指伊犁母马与英纯血公马的杂交群体,在本研究中称为英纯-伊犁马。原始伊犁马血样30 份,采集于2011 年的伊犁自治州昭苏县喀拉苏乡牧民家。英纯-伊犁马血样76 份,来自新疆维吾尔族自治区地方国营伊犁种马场。
1.2 基因组DNA 提取 部分马血液样品的DNA 提取使用常规酚-氯仿抽提法[8],部分样品使用全式金EasyPure®Blood Genomic DNA Kit(EE121-02)进 行提取。2 种方法对后续实验结果无影响。将基因组DNA溶解于TE 缓冲液,在-20℃冰箱保存备用。用1.5%的琼脂糖凝胶电泳对基因组DNA 进行质量检测。
1.3 PCR 扩增和测序 本实验对MSTN第一内含子区域(含SNP g.66493737T>C)和MSTN启动子附近227 bp的SINE 插入区域进行PCR 扩增和电泳检测。PCR 扩增体系为25 μL:2×Master Mix Buffer 12.5 μL,l0 μM 正向和反向引物分别0.5 μL,DNA(50 ng/μL)模板0.5 μL,加ddH2O 至25 μL。为了保证得到特异性产物,MSTN第一内含子片段扩增使用Touchdown 扩增方法:60℃退火2 个循环,59℃退火2 个循环,最后58℃退火28 个循环,每循环的延伸时间为45 s,其他步骤按Taq PCR Master Mix 说明书要求设置。SINE 插入PCR 反应使用59℃退火温度,延伸时间为90 s,进行30 个循环,其他步骤根据Taq PCR Master Mix 说明书要求设置。扩增试剂为Taq PCR Master Mix(生工生物工程(上海)股份有限公司,B639295)及相关产品。引物设计分别参考Dall'Olio[9]和Rooney[5](表1),引物合成和PCR 产物扩增由生工生物工程(上海)股份有限公司完成。
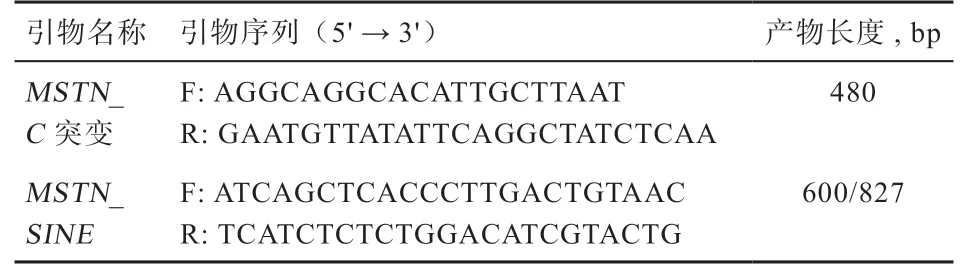
表1 MSTN 基因和SINE 插入引物序列和产物长度
1.4 数据分析 计算基因杂合度(Heterozygosity,He)使用公式、有效等位基因数(Effective Number of Alleles,Ne)和多态信息含量(Polymorphism Information Content,PIC)分别见公式(1)、(2)、(3)。

式中,n=等位基因数量,Pi=第i 个等位基因频率,Pj=第j 个等位基因频率。
连锁不平衡(LD)的r2值使用PLINK(v1.07,http://pngu.mgh.harvard.edu/purcell/plink/)软件的ld命令计算,其中ped 文件中个体的父母本和性别信息全部设为0。
2 结果与分析
2.1 伊犁马中MSTNg.66493737T>C SNP 多态性 对原始伊犁马和英纯-伊犁马MSTN第一内含子区域进行扩增,产物长度为480 bp(图1)。在MSTN基因g.2115位点处存在A 和G 多态性(图1)。由于本研究测得的序列与文献中的速度基因多态位点SNP g.66493737 T>C 为互补关系[10],因此G 突变即代表C 突变,按照文献惯例将本文中测得的G 突变称为C 突变。结果显示,原始伊犁马群体MSTN基因T/C 突变的有效等位基因数Ne 为1.142,主要以T 等位基因形式存在,C 等位基因频率只占0.07。原始伊犁马中没有CC 型纯合个体,CT 型杂合个体占13%。而在英纯-伊犁马中,C 等位基因频率为0.47,有效等位基因数Ne 为1.994,说明C 和T 2 个等位基因都是有效等位基因。在英纯-伊犁马中CC 型纯合个体占13%、CT 型杂合个体占68%、TT 型纯合个体占18%,基因杂合度He 为0.499,说明群体中的C 基因主要以杂合形式存在。

图1 MSTN 第一内含子PCR 扩增产物条带及多态位点基因型

表2 伊犁马中MSTN 的g.66493737C>T 基因型分布
2.2 伊犁马中SINE 插入多态性 通过PCR 产物等位基因多态性分析,对MSTN中有无SINE 插入进行检测。PCR 产物电泳检测结果显示,3 种类型的带型分别命名为II 型(2 条同源染色体都发生插入,双插入)、IN 型(一条同源染色体有插入而另一条没有,单插入)和NN 型(2条同源染色体都没有插入,无插入)(图2)。为了得到精确的产物长度和序列信息,将II 型和NN 型PCR产物连接至克隆载体后进行测序,得出有插入的片段长度为827 bp,无插入的片段长度为600 bp,插入序列长度为227 bp,与Rooney 等[5]报道的纯血马SINE 插入序列和长度都相一致。
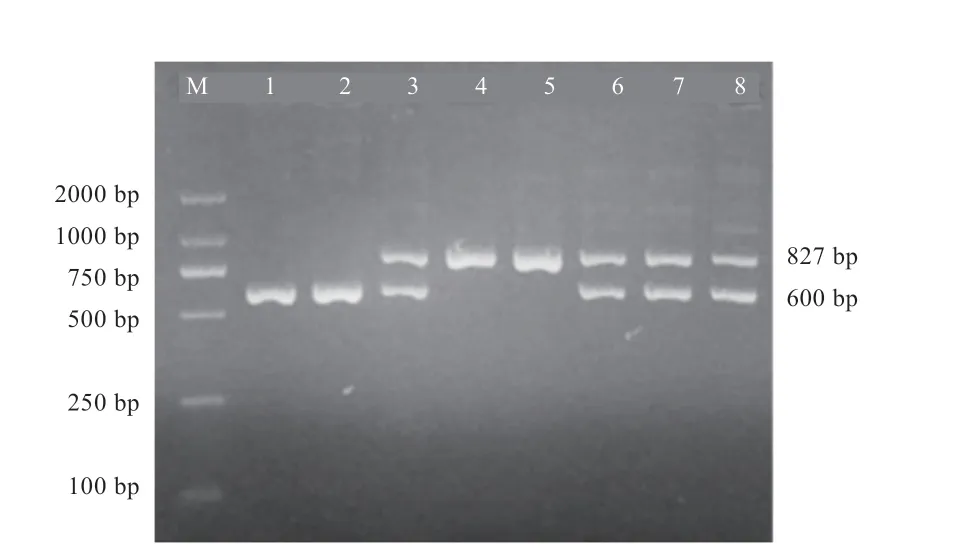
图2 SINE 插入PCR 产物电泳图
在原始伊犁马中未检测到SINE 插入多态性,均为NN 型个体。英纯-伊犁马中检测到SINE 插入(I),频率为0.43,有效等位基因数Ne 为1.959,说明2 个等位基因I(有插入)和N(无插入)均为有效等位基因。群体中II 型纯合个体占7%,IN 型杂合个体占72%,NN 型纯合个体占21%,基因杂合度He 为0.490,说明I 等位基因主要以杂合型分布。

表3 伊犁马中MSTN 的SINE 插入基因型分布
2.3 伊犁马中C 突变与SINE 插入的连锁关系分析 C突变与I 突变组合在一起理论上会存在4 种单倍型:I-C、N-C、I-T 和N-T(图3)。在原始伊犁马中,C 突变频率为0.07,没有I 突变。个体的基因型只有NNCT(4)和NNTT(26)2 种形式,群体中只有N-C 和N-T 2 种单倍型,N-C 频率仅为0.07(图3)。英纯-伊犁马中检测到IICC(5)、INCC(5)、INCT(50)、NNCT(2)和NNTT(14)5 种基因型。群体中有N-T(0.53)、I-C(0.43)和N-C(0.05)3 种单倍型,无I-T(图3)。根据PLINK(v1.07)软件的LD 分析,在杂交群体中SINE 插入与C 突变呈现较高连锁(r2=0.83)。
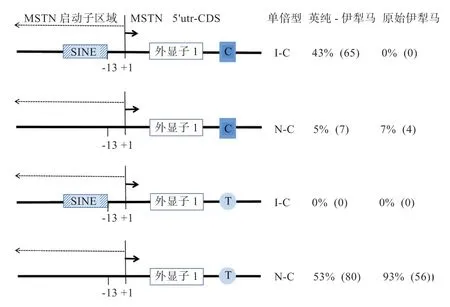
图3 MSTN 基因g.66493737T>C 和SINE 插入组合模式图及单倍型分布频率
纯血马以及有纯血马血统的夸特马中C 突变和SINE 插入呈现较高关联,而在其他的步态品种或地方品种,如美洲的密苏里狐快步马、田纳西走马,欧洲的比利时马、夏特兰马,亚洲的蒙古马中,C 等位基因分布较低或无,且与SINE 插入并无连锁关系[11](表4)。

表4 不同马群体中MSTN C 等位基因和SINE 插入频率
3 讨 论
马属动物不同物种以及家马不同品种的MSTN基因比较研究发现,g.66493737T>C SNP 位点的T 突变为野生型突变,且现在的大部分家马品种中都以野生型形式存在[11]。被称为“速度基因”[5,12-13]的C 突变在18 世纪至19 世纪的纯血马中还是很稀少的。大约从20世纪开始,人们对赛马的兴趣从耐力转向速度,之后便对纯血马的速度能力有了选择,因而MSTN的C 突变逐渐在群体中积累。C 突变的来源可被追溯到上世纪50 年代的一匹纯血种公马Nearctic,并由它的一匹优秀的种公马后代北方舞者(Northern Dancer)得以广泛传播[14]。新近研究表明,在MSTN上游启动子位置的一个SINE插入(I 突变)是真正引起“速度效应”的变异[5-6]。SINE 插入是广泛存在于高等真核生物基因组内的中等重复序列,占据了基因组的很大一部分,如在人类基因组中约占13.6%[15]。马的基因组中也分布着SINE 插入,其长度为227 bp[16]。由于SINE 插入发生在MSTN基因的转录启动子区域,导致MSTN基因表达水平显著下降,使纯血马肌肉中快肌纤维比例增加[5,11,16]。
本研究分析发现,原始伊犁马群体主要携带T 等位基因,C 等位基因分布频率较低,且均为N-C 单倍型,表明原始伊犁马的培育过程中没有I-C 单倍型渗入,与其当时的培育目标和引入的种公马品种有关。而在英纯-伊犁马群体中C 等位基因频率为0.47,SINE 插入频率为0.43,I-C 单倍型呈高度连锁(r2=0.83),与纯血马的研究结果(SINE 插入0.48,C 突变0.50)较为接近[11]。根据品种间的对比分析,多个携带C 突变的骑乘马品种中,并没有SINE 插入,只有在纯血马和夸特马2 个速度群体中发现了C 与I 的连锁。这一现象似乎说明与C突变相比,SINE 插入突变可能更适合作为速度基因标记。
多态信息含量是评估群体遗传多样性和衡量等位基因片段多态性的理想指标,多态信息含量越大,该位点的多态性就越丰富,群体内基因的变异性越高,选择潜力就越大[17]。当PIC>0.50 时,表示具有高度多态性;当0.25<PIC<0.50 时,表示具有中度多态性;当PIC<0.25 时,表示具有低度多态性[18]。本研究的英纯-伊犁马MSTN基因SINE 插入的多态信息含量:PIC=0.370,属于中度多态性,表明SINE 插入在英纯-伊犁马群体中的遗传性很稳定,群体内基因的变异程度为中等,发生遗传变异的可能性相对较小,具有较大的选择遗传潜力。
研究虽然已经证明了MSTN基因与纯血马最佳比赛距离的关联性,且目前已经广泛应用于预测纯血马的最适比赛距离[12-13]。但在英纯-伊犁马中,MSTN基因的C 突变和SINE 插入对速度能力的贡献,以及在肌纤维组成和蛋白表达层面的作用还有待于进一步深入研究。
4 结 论
以携带第一内含子C 突变为标记的MSTN速度基因,在英纯-伊犁马杂交群体中的分布频率达到47%,并与代表MSTN基因功能性变异的SINE 插入呈较高连锁,r2=0.83;而在伊犁马原始群体中C 突变频率仅为7%,且与SINE 插入为0 连锁。伊犁马中速度基因频率的提高为速度型伊犁马选育打下了良好的遗传多样性基础。
