山茶属连蕊茶组和毛蕊茶组资源及其种质创新研究进展
2021-12-14张亚利胡永红
张亚利,杜 诚,胡永红
(1.上海植物园/上海城市植物资源开发应用工程技术研究中心,上海 200231;2.上海辰山植物园/上海资源植物功能基因组学重点实验室,上海 201602)
连蕊茶组(Camellia Sect. Theopsis)和毛蕊茶组(Camellia Sect. Eriandria)隶属于山茶科(Theaceae)山茶属(Camellia),两组资源因其芳香[1—4]、抗花腐病[5]以及密花等特性,在观赏茶花的创新与应用领域日渐受到关注,尤其是在束花茶花品种创新领域[6],成为该品种群创新的重要资源并迅速走进育种者和应用者的视野。结合作者近年来对两组资源野外调查、迁地栽培及其在育种中应用的研究探索,对两组植物资源及其种质创新研究进展综述如下,旨在为两组资源的开发应用提供借鉴。
1 资源分类
1826年,山茶属连蕊茶组第一个物种柃叶连蕊茶(Camellia euryoides Lindl.)发表,但发现者并没有注意到雄蕊花丝部分联合这一典型性状。1887年,Pierre首次对山茶属(Thea L.= Camellia L.)进行属下次级划分,将山茶属划分为6个组。依据花萼宿存的特征建立拟茶组(Thea sect. Camelliopsis Pierre),尽管他同样没有注意到花丝部分联合的特征,但该组成员基本都具有花丝部分联合的特征[7]。1916年,Cohen Stuart第一次注意到花丝部分联合的特征,在Pierre的基础上将山茶属划分为 5个组,将花萼宿存,雄蕊花丝基部联合、内部浓密有毛,花柱被毛的类群建立毛蕊茶组(Camellia sect. Eriandria Cohen-Stuart);将花萼宿存,雄蕊花丝基部联合、内部无毛,花柱无毛的类群建立连蕊茶组(Camellia sect. Theopsis Cohen-Stuart)[8]。1940 年,Nakai以毛蕊茶组为基础建立尾叶山茶属(Camelliastrum Nakai),以连蕊茶组为基础建立连蕊茶属(Theopsis Nakai),分别收录了中日两国产的 6种毛蕊茶组和14种连蕊茶组植物[9—10]。1958年,Sealy[11]仍将连蕊茶组和毛蕊茶组视为山茶属的属下次级划分,而没有接受Nakai提升为两个独立属的概念。Sealy没有注意到Cohen Stuart的处理,在Pierre和中井的基础上将连蕊茶组和毛蕊茶组分别命名为 Camellia sect. Theopsis Sealy和Camellia sect. Camelliopsis(Pierre) Sealy,分别收录了连蕊茶组29种、2变种,毛蕊茶组6种,共计35种、2变种[11]。在此期间,两组资源的新种发表及分类研究主要集中在国外。随后,国内山茶属研究逐渐深入,先后出现了张宏达和闵天禄的分类系统[12—14]。2000年后,Orel等[15]开展了大量的越南野生山茶属资源调查与研究,并发表新组大叻组(Camellia sect. Dalatia Orel),对山茶属的分类系统有了新的观点。
1.1 Sealy分类系统的连蕊茶组和毛蕊茶组资源
Sealy将山茶属植物分成12组82种,其中连蕊茶组29种,毛蕊茶组6种,两组共计35种[11],但在连蕊茶组的分类检索表中仅列出连蕊茶组 28种及 2变种。Sealy分类系统中,除了 Camellia transnokoensis Hayata未列出之外,主要的两个存疑问题在张宏达及闵天禄的分类系统中得到一致的解决方案:(1) 樱花短柱茶(Camellia maliflora Lindl.)和中越山茶(Camellia indochinensis Merr.)均不属于连蕊茶组和毛蕊茶组;(2) 张宏达及闵天禄均将大萼毛蕊茶(Camellia assimiloides Sealy)、小果毛蕊茶(Camellia villicarpa S. S. Chien)、斑枝毛蕊茶(Camellia punctata Cohen-Stuart)、四川毛蕊茶(Camellia lawii Sealy)等4种归到毛蕊茶组。基于此,截至1958年,山茶属连蕊茶组资源为23种、毛蕊茶组资源为10种(表1)。
1.2 张宏达分类系统的连蕊茶组和毛蕊茶组资源
随着我国野生资源普查及调查工作的不断深入,我国山茶属植物系统研究也不断深入。1981年张宏达先生发表《山茶属植物的系统研究》,建立新亚属后生山茶亚属(Camellia subgen. Metacamellia Hung T. Chang),其中连蕊茶组44种(检索表中列出42种),毛蕊茶组17种(检索表中列出14种)[12];1984年,Camellias中的后生山茶亚属收录57种,其中连蕊茶组43种、6变种,毛蕊茶组14种[13];1998年出版的《中国植物志》山茶科基本依照张宏达的分类系统,收录连蕊茶组资源 49种(列出原产我国的46种),毛蕊茶组14种(全产我国)[16]。该分类系统除了将 Sealy系统中川滇连蕊茶(Camellia synaptica Sealy)处理为云南连蕊茶的变种[Camellia tsaii Hu var. synaptica (Sealy) Hung T. Chang],将尖萼连蕊茶(Camellia acutisepala H. T. Tsai & K. M.Feng)处理为蒙自连蕊茶的变种尖萼蒙自连蕊茶[Camellia forrestii (Diels) Cohen-Stuart var.acutisepala (H. T. Tsai & K. M. Feng) Hung T. Chang]之外,保留了 Sealy系统中毛蕊茶组和连蕊茶组其他种,并在此基础上发表29个新种、3个新变种。
胡先骕[17]发表了包括疏叶毛蕊茶(Camelliastrum laxiflorum Hu nom. nud.)、麻栗坡毛蕊茶(Camelliastrum marlipoense Hu nom. nud.)在内一批山茶科类群,但不属于合格发表。1965年,又在连蕊茶属(Theopsis Nakai)发表了金花茶(Theopsis chrysantha Hu)、卫矛叶连蕊茶(Theopsis euonymifolia Hu)、长梗连蕊茶(Theopsis longipedicellata Hu)和龙岩山茶(Theopsis lungyaiensis Hu)[18],但这些类群最终都分别被作为古茶组(Camellia sect. Archecamellia Sealy)的金花茶(Camellia petelotii (Merr.) Sealy)、油茶组(Camellia sect. Paracamellia Sealy)的落瓣油茶(Camellia kissii Wall.)、长梗茶组(Camellia sect.Longipedicellatae Hung T. Chang)的长梗茶[C.longipedicellata (Hu) Hung T. Chang & D. Fang]和油茶组的短柱油茶[Camellia brevistyla (Hayata)Cohen-Stuart]的异名[16]。
结合《中国植物志》中列出分布于我国的连蕊茶组原种46种、毛蕊茶组14种,加上日本分布的琉 球 连 蕊 茶 (Camellia lutchuensis T. Itô &Matsum.)[19]、1991年发表的假长管连蕊茶(Camellia pseudoelongata Hung T. Chang & S. X. Ren)[20]以及1992年发表的长管毛蕊茶(Camellia tubiformis Hung T. Chang & S. X. Ren)[21]。截止2000年,按照张宏达的分类系统,连蕊茶组资源应为48种,毛蕊茶组为15种(表1)。
1.3 闵天禄分类系统的连蕊茶组和毛蕊茶组资源
2000年,中国科学院昆明植物研究所闵天禄研究员出版《世界山茶属的研究》一书[9],山茶属订正为2亚属14组119种。其中,连蕊茶组19种,毛蕊茶组9种,隶属于茶亚属[subg. Thea (L.) Hung T.Chang]。Flora of China沿用了闵天禄的分类系统[22]。与前述两个分类系统相比,该分类系统对已有发表种进行了大量归并,从而大大缩减了两组物种的种类,归并情况具体见表 1。除了归并之外,闵天禄分类系统与上述两个分类系统的不同还包括:(1) 玫瑰连蕊茶(Camellia rosiflora Hook.)被认为是园艺杂交种,不再被收入该分类系统;(2) 厚瓣短蕊茶(Camellia crassipetala Hung T. Chang)自短蕊茶组(sect. Brachyandra Hung T. Chang)[8]转入连蕊茶组,并作为窄叶连蕊茶(Camellia tsaii Hu)的异名;(3) 增加1新种秃肋连蕊茶(C. glabricostata T. L. Ming)和1新变种线叶连蕊茶(Camellia viridicalyx Hung T.Chang & S. Y. Liang var. linearifolia T. L. Ming)。
结合叶创兴等[23]2001年发表毛蕊茶组新种毛药山茶(Camellia renshanxiangiae C. X. Ye & X. Q.Zheng),Dung等[24]在2016年发表越南连蕊茶组新种(Camellia quangcuongii L. V. Dung, Son & Nhan),截至2020年,按闵天禄系统连蕊茶组应为20种,毛蕊茶组为10种(表1),较张宏达分类系统种类数有大幅减少。
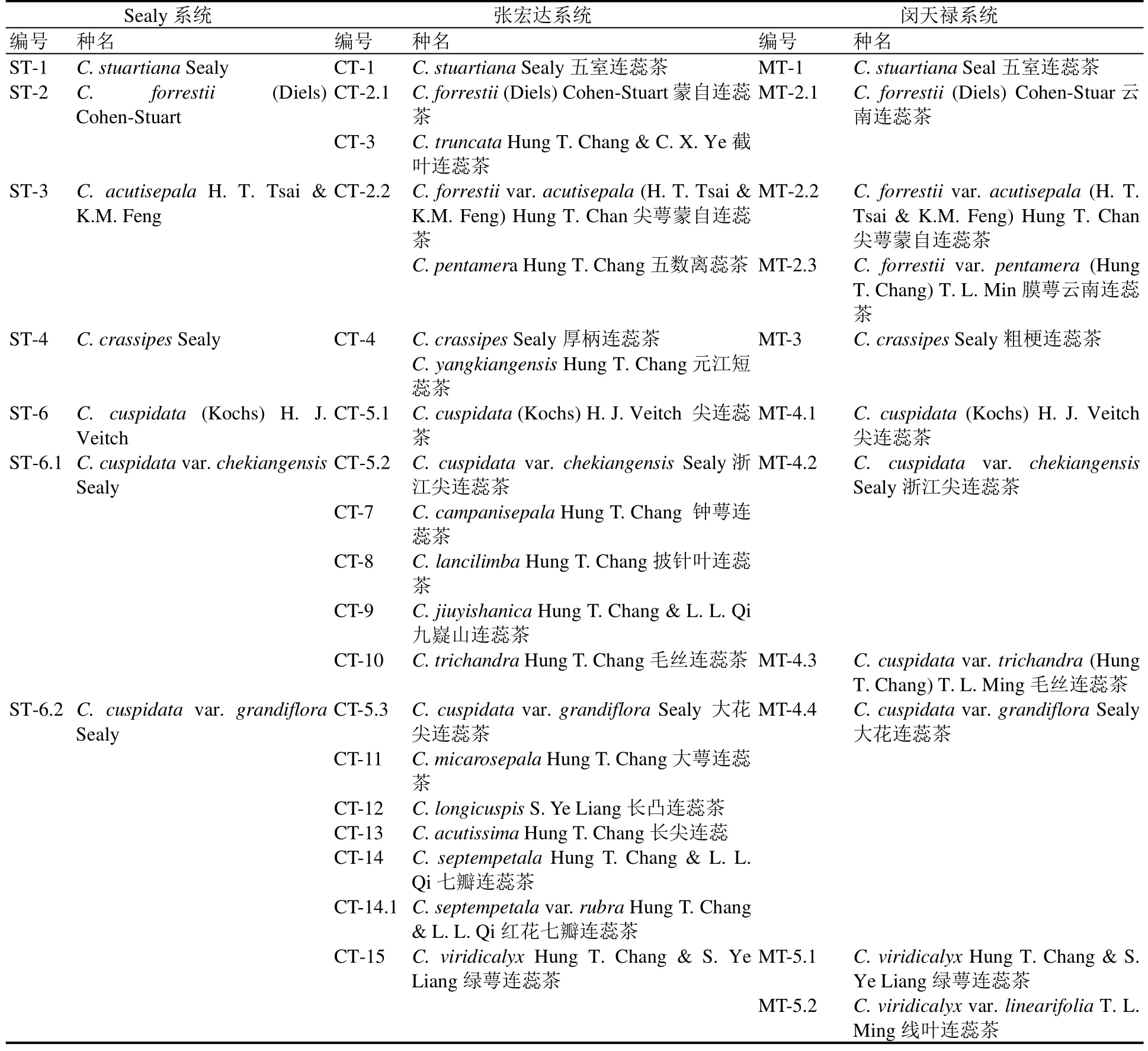
表1 不同分类系统中连蕊茶组和毛蕊茶组植物资源对照Table 1 The development of taxonomic study in Camellia Sect. Theopsis and Sect. Eriandria
1.4 近年来关于连蕊茶组和毛蕊茶组的讨论
20世纪,Sealy、张宏达、闵天禄三者对山茶属的种类、系统演化、起源及扩散存在一致或不同或相反的观点。在其系统发育上,张宏达及其团队[25]认为,花无梗,苞被不分化为苞片和萼片,代表了山茶属的原始特征;而花梗伸长,引起苞萼分化,苞片在花梗上疏离,宿存,脱落或减少,演化出更加进步的类群,而连蕊茶组和毛蕊茶组则为山茶属中较为进化的类群。闵天禄等[26]认为山茶属系统发育的演化趋势包括明显具花梗,小苞片散生在花梗上至花梗缩短至无梗或近无梗,小苞片与萼片密集排列组成由小到大的连续系列等17个。其中,两者对山茶属系统发育的认识主要区别在于花梗长短及苞被分化上的演化趋势。
进入21世纪,随着分子生物学等技术的发展,关于山茶属系统发育的研究逐渐增多。皮二旭[27]开展云理论在山茶属植物数值分类学中的应用研究,杨俊波等[28]对4个DNA 片段在山茶属分子系统学研究中的应用进行初步探索,基于tsnL-F、rpL16和waxy三组数据所建分子系统树支持山茶属为一单系,但属下系统由于取样的原因有待进一步研究。Vijayan等[29]利用ITS片段对112个山茶属物种进行系统学研究,初步确定连蕊茶组和毛蕊茶组共同构成一个单系分支。吕林玲通过对该亚属16种1变种植物的 3个叶绿体基因片段(psbA-trnH、trnL-F和rpl32-trnL)的分子系统发育分析,结果显示连蕊茶组和毛蕊茶组是一自然单系类群,建议将两组合并处理[30],江正栋通过叶绿体 DNA构建的系统树主要分为 A~M 共 13个分支,其中 C 和 I 分支基本由连蕊茶组组成,毛蕊茶组物种也分布在这两个分支中。此外,其花梗的祖征重建结果显示山茶属原始类群是具长花梗的,并在分化过程中逐渐形成了短花梗及无花梗的类型,这一结果与闵天禄认为山茶属向花梗缩短的方向演化的理论相契合,与张宏达系统认为的山茶属原始类群花无梗相反[31]。以上研究均呈现出连蕊茶组和毛蕊茶组为单系类群的初步结论,为进一步开展两组资源的研究奠定了基础。

(续表)

(续表)
2 观赏性及适应性
近年来,随着连蕊茶组和毛蕊茶组资源在芳香等观赏特性及抗花腐病等适应性方面的研究深入,两组资源逐渐受到广泛关注[32—33],如川鄂连蕊茶(Camellia rosthornianaHand.-Mazz.)等逐渐在生产和园林中应用[34]。
2.1 观赏性研究
从观赏特征来看,两组资源具有多枚花蕾顶生和腋生的特征,从而形成串花或簇花的密花效果,如岳麓连蕊茶(Camellia handeliiSealy)、毛枝连蕊茶[Camellia trichoclada(Rehder) S. S. Chien]单枝上多枚花蕾腋生,花似成串开放,毛药山茶等顶生花蕾繁多,形成簇花效果(图1: A~C)。在花色上,两组资源以白色为主,但部分种存在花蕾粉红色,花白粉色的单株,如岳麓连蕊茶、毛花连蕊茶(Camellia fraternaHance)等。此外,连蕊茶组和毛蕊茶组叶片大小主要集中在2~11 cm,其中部分种在2~4 cm,如黄杨叶连蕊茶(Camellia buxifoliaHung T. Chang)、毛枝连蕊茶等,部分野生资源具有嫩叶红色,或是在不同的栽培环境下呈现黄绿色或红色的嫩叶效果,极具观赏性(图1: D~F)。

图1 部分迁地栽培连蕊茶组和毛蕊茶组植物的花、叶观赏性Fig. 1 Ornamental characters of some ex-situ species in Camellia Sect. Theopsis and Sect. Eriandria
除了开花繁密、叶形精致、嫩叶色泽丰富等观赏特征之外,花芳香是两组资源的另一个重要特征。感官评价显示,连蕊茶组蒙自连蕊茶、毛花连蕊茶、秃梗连蕊茶(Camellia dubiaSealy)、岳麓连蕊茶、琉球连蕊茶、云南连蕊茶及毛蕊茶组广东毛蕊茶(Camellia mellianaHand.-Mazz)具花香[11]。随着检测方法的不断发展,两组植物基于挥发性成分分析的芳香资源陆续被报道(表2)。已报道10种连蕊茶组植物的花挥发性成分显示,芳香型主要包括以苯乙醇、芳樟醇、柏木醇等化合物为主的玫瑰香型、铃兰香型、柏木香型[2—4],2019年,Fan等[1]对22种连蕊茶组植物的花挥发性成分分析,鉴定出苯类、苯丙酸类、萜类等223种挥发性化合物,丰富了两组植物资源的芳香成分研究。
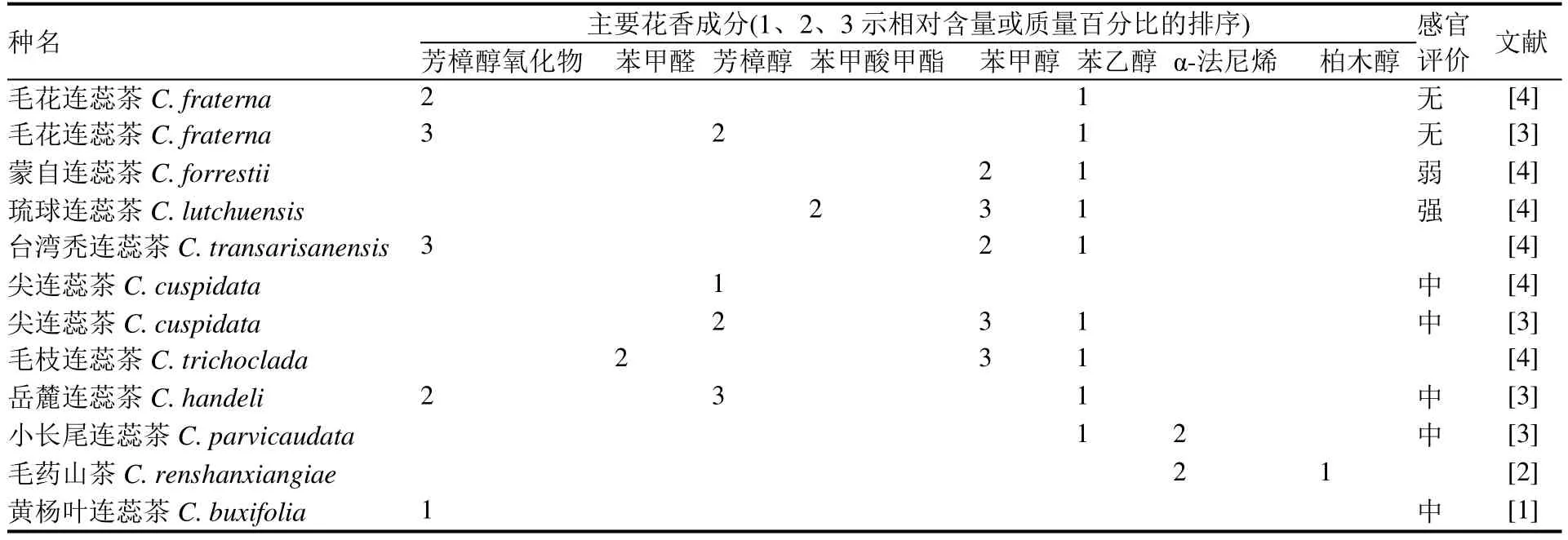
表2 连蕊茶组和毛蕊茶组部分植物的主要花香成分Table 2 Main aroma components of some species in Camellia Sect. Theopsis and Sect. Eriandria
2.2 适应性研究
在连蕊茶组和毛蕊茶组资源中,其抗逆性也越来越多地被发现和关注,主要集中在耐热性、抗花腐病和耐盐碱等方面。
在野生资源的耐热性研究中,李辛雷等[35]以山茶属17组130个物种为材料,通过相对电导率、丙二醛含量等生理指标的测定分析,认为耐热性较强的有离生雄蕊茶组(Sect.CorallinaSealy)、秃茶组(Sect.GlaberrimaChang)、毛蕊茶组、茶组[Sect.Thea(L.)Dyer]和连蕊茶组。彭绍峰等[36]对21个山茶种质进行耐热性评价,其中2个来自连蕊茶种源的半致死温度均在50 ℃以上,遗憾的是并未指出具体的种。在连蕊茶组培育后代的耐热性研究中,张亚利等[37]对5个束花茶花新品种的耐热性研究显示,‘垂枝粉玉’(Camellia‘Pink Cascade’)的耐热性最强,其半致死温度为64.90±2.99 ℃,45 ℃阶段高温胁迫(8:00~20:00,室温45 ℃;20:00~08:00,室温40 ℃)处理2 d后叶片损伤率为1.10%。此外,王江英等[38]对连蕊茶组24种植物在夏季极端高温干旱条件下的耐旱潜力评价显示,小卵叶连蕊茶(Camellia parvi-ovata)、七瓣连蕊茶(Camellia septempetalaHung T. Chang & L. L. Qi)、长尖连蕊茶(Camellia acutissimaHung T. Chang)、贵州连蕊茶(Camellia costeiH. Lév.)和大萼连蕊茶(Camellia micarosepalaHung T. Chang)的耐旱性最强。
连蕊茶组和毛蕊茶组植物的抗花腐病研究主要见于国外。Denton-Giles以(Camellia‘Nicky Crisp’)和(Camellia lutchuensis)作为亲和与不亲和寄主,显微分析显示出诱导植物抗性的几个特征,包括乳头形成、H2O2积累和局部细胞死亡;进一步对山茶属40个野生种及1个杂交种的分析显示,其中供试的连蕊茶组11个种均属于不亲和性寄主,但对花腐病的抗性存在差异[5]。Denton-Giles对18个连蕊茶组的后代进行抗花腐病研究显示,在琉球连蕊茶、台湾秃连蕊茶、毛花连蕊茶、蒙自连蕊茶及闽鄂山茶(Camellia grijsiiHance)的后代中,琉球连蕊茶后代对花腐病的抗性最强[39]。
在耐盐碱研究方面,张亚利等[40]对连蕊茶组野生资源的后代‘小粉玉’(Camellia‘Xiaofenyu’)在不同土壤中的适应性研究显示,‘小粉玉’可在pH 8.16 的土壤中正常生长。郭卫珍等[41]以连蕊茶组资源的后代‘玫玉’(Camellia‘Meiyu’)为材料,设置0、50、100、200、400 mmol·L-1NaCl梯度进行盐胁迫试验,其中50 mmol·L-1NaCl处理的植株在处理28 d后,相对生长量为65%,盐害指数为22.2%,植株生长状况良好,且慢慢适应了低盐环境。
3 资源种质创新
2007年,William[32]提出“Cluster-flowering Hybrids”,“Cluster-flowering”一词主要用来描述精致繁密的花及相应的小叶这一茶花品种特征,所提及的育种亲本主要为连蕊茶组的部分资源。2009年,沈荫椿[42]提到微型山茶花,突出了小花、芳香的特征,并再次提及毛花连蕊茶等连蕊茶组资源。2018年,张亚利等[6]结合对连蕊茶组和毛蕊茶组资源及其种质创新的研究,逐步形成和完善了束花茶花的概念,束花茶花主要指多枚花蕾顶生和腋生,花似成束(串)开放的茶花品种,主要以山茶属后生山茶亚属的种质资源为亲本。
3.1 育种进展
自 1939年前 Hiller 以原产中国的尖连蕊茶[Camellia cuspidata(Kochs) H. J. Veitch]为亲本培育出束花茶花品种‘Winton’至今,束花茶花品种已有300余种[6]。束花茶花育种源于英国,兴于日本、美国、澳大利亚和新西兰,以日本为例,利用本土分布的琉球连蕊茶就培育出60余个品种。
应用于种质创新的连蕊茶组资源有10余种,主要集中在尖连蕊茶、毛花连蕊茶、琉球连蕊茶、玫瑰连蕊茶、云南连蕊茶等种类,其中尤以毛花连蕊茶和琉球连蕊茶为多;毛蕊茶组有1~2种[6]。现有束花茶花品种的花色以白粉色及粉色为主,部分呈深红色,花型以单瓣、半重瓣居多;香花品种主要为琉球连蕊茶后代。在新种质抗逆性研究方面,新西兰梅西大学(Massey University)与美国加利福尼亚大学戴维斯分校(University of California, Davis, CA)合作,开展杂交种抗花腐病研究[5]。上海植物园针对耐盐碱土壤、耐全光照环境等抗性开展品种创新与研究[43—44]。
3.2 国内种质创新研究进展
如前所述,连蕊茶组和毛蕊茶组资源主要分布在我国,但我国对两组植物的种质创新工作起步较晚。随着近年来相关研究者及育种者对两组资源的关注度日渐提高,其种质创新工作也逐步发展,2010~2019年,在国家林业和草原局获批新品种保护的品种共9件(表3)。
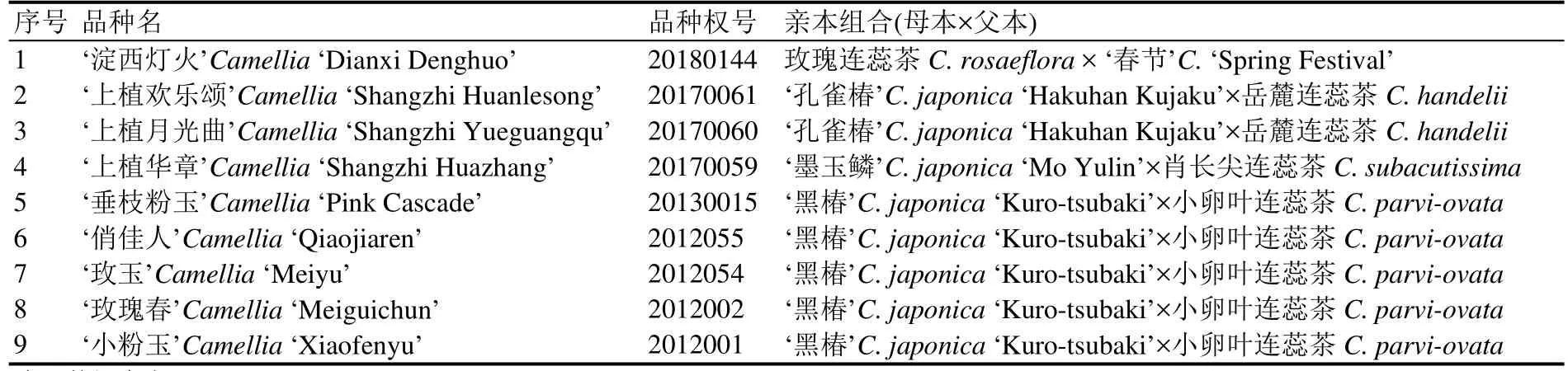
表3 以连蕊茶组资源为亲本培育的新品种(中国)Table 3 Varieties bred from Camellia Sect. Theopsis in China
在山茶属组间杂交研究中,连蕊茶组是最难在育种中获得成功的一个,部分资源只能作父本;即使可以作母本,成功率也较低[45]。从我国获批的新品种来看,除了‘淀西灯火’(Camellia‘Dianxi Denghuo’)以连蕊茶组资源为母本之外,其余8个品种均以连蕊茶组资源为父本。张亚利等[6]、胡禾丰等[46]对以连蕊茶组资源为亲本的205个品种进行分析显示,以连蕊茶组植物为母本的非机遇苗选育品种仅在10%左右,进一步采用提前授粉、延迟授粉等方法,都未能获得到杂交种子。由此可见,以两组资源为亲本的种质创新存在杂交低(不)亲和现象,从而制约了种质创新的迅速发展。此外,‘金叶粉玉’、‘鱼叶粉香’等以连蕊茶组和毛蕊茶组资源为父本的新品种及杂交实生苗也逐步选育出来[47],为两组山茶资源的应用不断提供丰富的素材。
此外,两组资源的挖掘与利用研究不断丰富,如园林应用[48]及作为茶油资源的开发[49]等。
4 展望
连蕊茶组和毛蕊茶组作为主要分布在我国的山茶属资源,以其在观赏性、芳香性及抗逆性等方面的潜质,为丰富园林景观的茶花种质等提供了丰富的素材。但是,在实际应用中,物种的识别为应用者带来了较大困扰,应用者按照分种检索表往往出现模棱两可、无所适从的困境,这也对两组资源的进一步开发利用形成制约。因此,在两组植物资源的分类及品种选育等方面仍需开展系统深入研究,如相对稳定的典型识别特征提取及连续(稳定)变异的判断标准等,不仅有助于分种检索表的科学发展,更便于非分类专业人员的实际应用,尤其是育种者。
如前所述,在两组植物资源的种质创新中还存在花色、花型较为单一的情况,耐热、耐盐碱及抗花腐病等抗性育种还处于起步阶段。与此同时,我国作为连蕊茶组和毛蕊茶组资源的主要分布地,在种质创新研究中所培育品种数占世界范围内已培育数量(300余品种)比例不足5%。因此,应结合我国特有连蕊茶组与毛蕊茶组资源,开展芳香、抗花腐病、耐盐碱等育种目标的种质创新,野生资源在园林景观中的应用开发等工作,从而不断提升我国野生资源的开发应用及自主品种创新研发能力。
