miR-24家族的生成机制及生物学功能研究进展
2021-12-13沈曼曼侯启瑞王克华
沈曼曼,李 涛,侯启瑞,曲 亮,王克华,马 猛,吴 萍*
(1.江苏科技大学 生物技术学院,镇江 212100) (2.中国农业科学院 江苏省家禽科学研究所,扬州 225212)
miR-24是一种重要的miRNA,在多种生命过程中发挥了关键的功能作用.miR-24的表达受到一些重要转录因子、非编码RNA等调控影响,在癌症发生、心血管调控、炎症反应、脂质代谢、繁殖调控等方面发挥了重要作用.文中对miR-24的生成机制及miR-24在机体内的调控功能最新研究进展作一综述.
1 microRNA的作用机制及生物功能
microRNA(miRNA)是一类长度为18~25个核苷酸的非编码单链RNA分子,是基因表达的重要调控因子,在各种生物过程中发挥重要的作用.其生物合成和表观遗传机制形成一个调控网络,相互调节,共同调控一系列生物过程,广泛参与细胞的增殖、分化、凋亡以及激素的合成与分泌等过程.miRNA的生物合成受到转录因子、非编码RNA等诸多因素的调控.转录因子主要通过与miRNA启动子区域相互作用调控miRNA的表达,如SMAD可以和Drosha复合体相互作用,通过提高Drosha加工过程增加一些miRNA的表达.一个miRNA也可以同时受多个转录因子的调控影响癌症的发生及发展.非编码RNA通过miRNA反应元件(miRNA response element, MRE)吸附miRNA,以海绵体的角色影响miRNA的功能发挥.随着对miRNA的深入研究,发现其调控作用呈现出多样化、复杂化以及精准化等特点.动物miRNA的功能发挥主要是其种子序列与靶基因的3′非编码区(untranslated region, UTR)、5′UTR以及蛋白质编码区(coding sequence,CDS)结合发挥生物学功能.
2 miR-24的生成机制
2.1 miR-24的生成
miR-24首次在2001年被Lagos-Quintana等报道,目前相关研究报道已达751余篇(PubMed,截止至2020年12月),在各种组织中均有表达但表达水平不同,广泛参与了细胞增殖、分化、凋亡等生命事件,影响了细胞发育和代谢、癌症发展、炎症反应、病毒互作[1]等多个生命进程.人、小鼠、猪、牛、鸡等物种miR-24均由两条染色体剪切形成,人的miR-24分别来自于9号和19号染色体,小鼠的miR-24来自于8号和13号染色体,猪的miR-24来自于2号和10号染色体,牛的miR-24来自于7号和8号染色体,鸡的miR-24来自于30号染色体和Z染色体.由miR-24形成两种不同序列的剪接体:miR-24-3p和miR-24-5p,miR-24-3p发挥的生物学效应较为广泛,其序列为5’-UGGCUCAGUUCAGCAGGAA-3’,在人、小鼠、猪、牛、鸡等物种中成熟序列高度一致(表1).
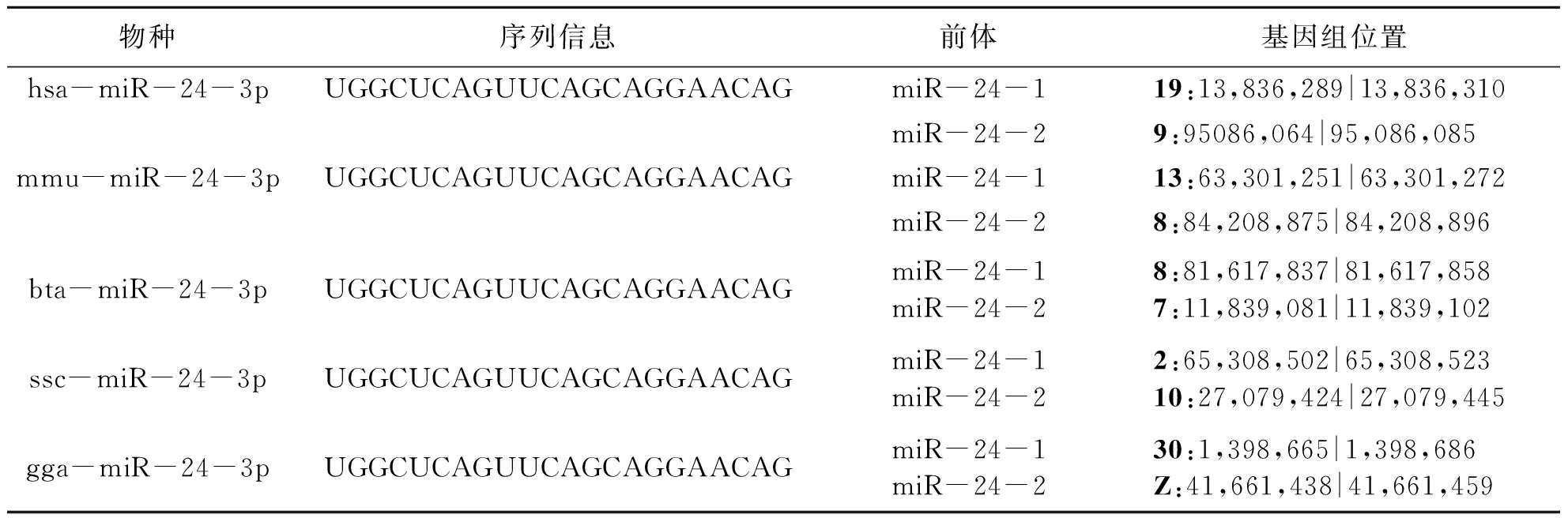
表1 各物种miR-24-3p序列信息及其染色体位置
2.2 转录因子对miR-24的调控作用
转录因子通过调控miR-24上游区域影响其表达从而调控相关靶基因在细胞中的功能作用.miR-24-2上游含有RUX2转录因子结合位点,RUX2能够降低miR-24的表达,从而解除miR-24对靶基因SATB2的抑制作用,影响骨细胞的分化[2].SMAD5作为BMPR4的下游转录因子,磷酸化后能够与miR-24-3p上游启动子区的结合位点作用,阻碍由BMP4诱导的胚胎干细胞凋亡过程[3].在结肠癌的研究中发现HIF-1α能够与miR-24-3p启动子区结合,促使miR-24-3p的靶基因PDHB、PDHA1、DLD等发挥功能作用,促进细胞存活[4].
2.3 非编码RNA对miR-24的表达调控作用
基于竞争性内源RNA(competing endogenous RNAs, ceRNA)理论,研究发现生物体内miR-24的表达也受非编码RNA的调控.非编码RNA包括了长链非编码RNA(long non-coding RNA, lncRNA)、环状RNA(cicular RNA,circRNA)、转运RNA(transfer RNA,tRNA)和核糖体RNA(ribosomal RNA,rRNA)等,其中以lncRNA调控miR-24的研究最多,广泛影响癌症的增殖、凋亡、纤维化等过程,参与癌症的进展.lncRNA ANRIL通过下调miR-24表达,解除miR-24对靶基因的抑制从而促进视网膜母细胞瘤细胞的存活、迁移和侵袭[5].lncRNA MIR22HG高表达可抑制miR-24-3p在甲状腺乳头状癌的高表达,从而参与癌症发展过程[6].LncRNA IGBP1-AS1通过竞争性吸附miR-24-1影响ZIC3基因的表达,从而抑制乳腺癌的侵袭和增殖[7].
miR-24不仅介导lncRNA对癌症的发生进展过程,也广泛参与了其他疾病的发生过程.lncRNA Gm5091可通过海绵吸附miR-27b~23b~24降低小鼠酒精性肝纤维化[8].linc00472启动子甲基化异常导致linc00472表达水平降低,从而降低linc00472对miR-24的吸附作用,使miR-24表达水平升高,参与调节心房颤动[9].文献[10]对小鼠心肌损伤模型研究发现,非编码lncRNA CYTOR通过与miR-24种子序列2-8位置结合调控其表达影响靶基因XIAP mRNA和蛋白的作用,从而调控小鼠脓毒病症状.
随着非编码RNA研究的深入进行,发现circRNA也广泛地参与到了生命过程中,且部分circRNA的发挥也需要miR-24的参与.文献[11]对成肌细胞分化过程中进行circRNA和miRNA测序分析,结果发现在成肌细胞分化过程中miR-24与circRNA表达呈负相关关系,表明了circRNA可能通过调控miR-24影响成肌细胞分化过程.
3 miR-24的调控功能
miRNA主要通过其成熟体的第2~8个碱基与靶基因的UTR或CDS区域完全或部分互补配对,诱导靶mRNA的降解或抑制靶基因的蛋白合成,从而在转录水平上调控基因表达.miR-24主要与靶基因3′UTR、5′UTR和CDS区域结合发挥生物学功能(表2).
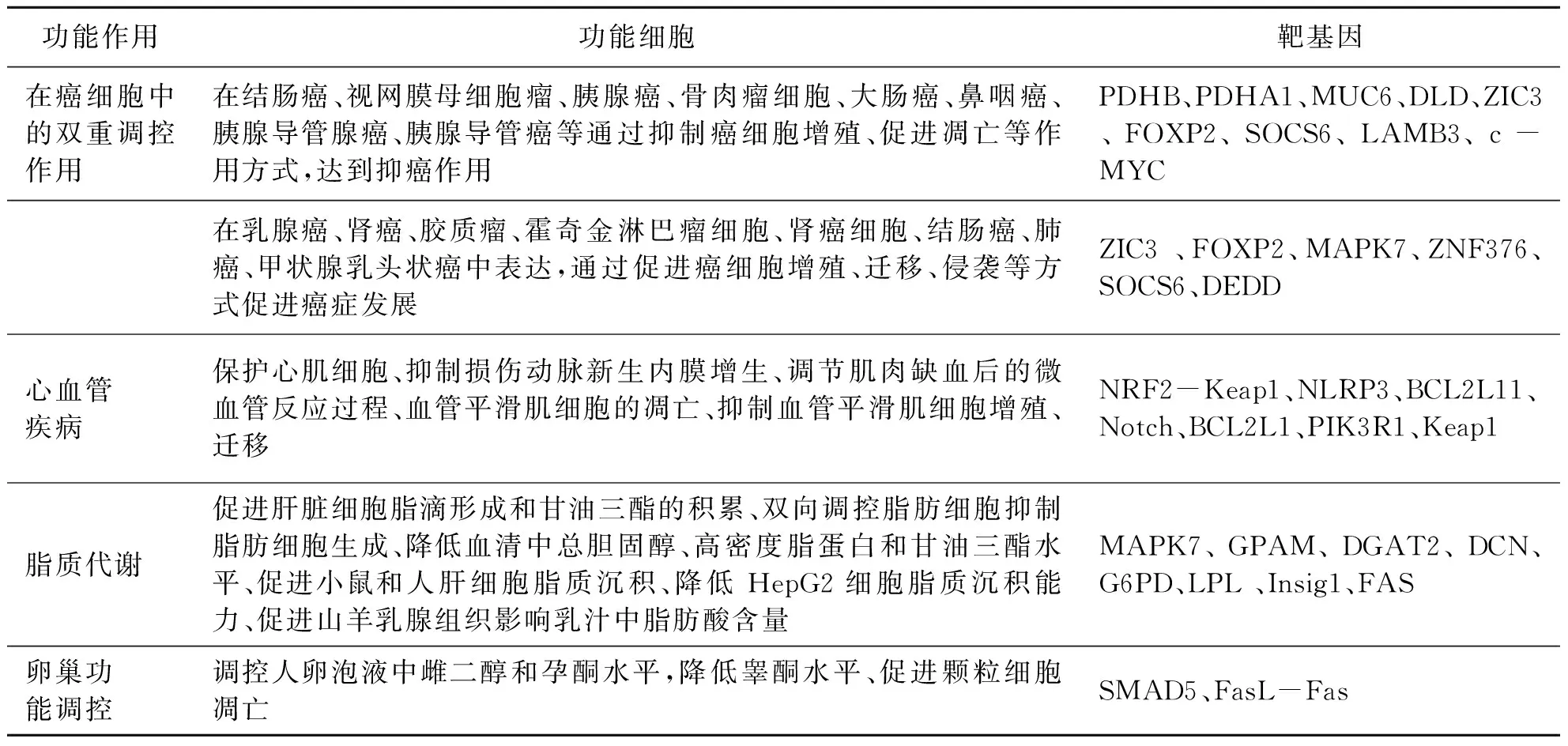
表2 miR-24在癌症、心血管疾病、脂代谢及卵巢发育中的功能作用及其靶基因
以miR-24-3p为搜索关键词,在miRWalk数据中已验证的hsa-miR-24-3p靶基因有2000多个,包括BCL2L11、CYP11B2、SP1等(http:∥mirwalk.umm.uni-heidelberg.de/interactions/).miR-24对靶基因的调控也表现了复杂性特点,不同的miR-24成熟体会靶向不同的基因达到共同调控作用.同一个成熟体对部分基因可以单独靶向3′UTR、5′UTR或CDS区域发挥调控作用,也会同时靶向基因的3′UTR和5′UTR区域发挥复杂的调控作用.文献[12]对小鼠的胚胎干细胞(ESC)研究发现miR-24-3p和miR-24-2-5p能够共同靶向KLF4和C-MYC的3′UTR区域,并负反馈调控PRMT7的表达,从而诱导ESC的全能分化特性.文献[13]研究发现miR-24-3p能够同时靶向JAB1 3′UTR和5′UTR影响鼻咽癌的进展过程.miR-24通过多种复杂的调控网络参与了诸多的生物过程.
3.1 miR-24对癌症发生的作用
miR-24广泛参与多种癌症的进展过程,在生命体内通过不同的调控途径参与癌细胞的增殖、凋亡、迁移和侵袭等过程,然miR-24受多种因素的影响以及调控多个靶基因对癌症的发生过程发挥双重作用.诸多研究通过构建小鼠疾病模型或体外培养人癌细胞,在细胞和个体水平上验证miR-24对肿瘤发展的作用影响,利用增殖、凋亡、靶点互作等研究技术验证其相关功能.研究发现miR-24在乳腺癌、胶质瘤、霍奇金淋巴瘤细胞、肾癌细胞中高表达,通过促进癌细胞增殖、迁移、侵袭的能力促进癌症发展.其作用的发挥也主要通过与靶基因的UTR区域结合,如miRNA-23b~27b~24 主要通过靶向转录因子FOXP2基因的启动子区域,从而降低FOXP2基因的mRNA和蛋白的表达水平,促进结肠癌细胞的迁移[14].miR-24通过靶向MAPK7、ZNF376[15]促进肺癌细胞增殖、迁移和侵袭.miR-23a~27a~24-2基因家族通过靶向SOCS6基因抑制其表达,从而促进胃癌细胞的增殖和迁移作用[16].
此外,miR-24在一些癌症进展过程中起到抑制的作用,如胰腺癌、骨肉瘤细胞、大肠癌、鼻咽癌等.β-catenin基因突变广泛参与各种癌症的发生过程,miR-24通过抑制β-catenin表达从而抑制癌症发生,是β-catenin基因有效调节因子.在胶质瘤中研究发现miR-24能够调节β-catenin基因从而影响胶质瘤细胞的存活和自噬功能[17].在结肠癌中,miR-24-1-5p可以靶向β-catenin基因从而抑制β-catenin的表达,达到抑制癌细胞增殖、迁移和存活的能力[18].在其他各类癌症中,miR-24通过靶向DND1抑制胃癌细胞的增殖、迁移和侵袭的作用,靶向LAMB3抑制胰腺导管腺癌的增殖,调控SW48细胞的凋亡抑制大肠癌[19-20]等多种癌症进展过程,在上述抑癌作用中可将miR-24作为治疗癌症的靶点,从而提高化疗的敏感性.
在癌症发生过程中miR-24在血清中的表达水平可以作为癌症的临床诊断标记.文献[21]研究发现miR-24-3p在口腔鳞状细胞癌肿瘤组织表达水平较高,且外源性的miR-24-3p增加了恶性肿瘤细胞的增殖作用,揭示唾液中的miR-24-3p可以作为口腔鳞状细胞癌筛查的潜在检测标志物.文献[22]研究发现口腔鳞状细胞癌患者血清中miR-24的表达水平较正常人要高,进一步说明miR-24可以作为一种口腔癌诊断工具.除了在口腔癌中,在对胃癌的研究发现miR-24在早期胃癌的表达水平显著低于晚期胃癌血清中,上述研究提示miR-24可以作为癌症进展的生物标记物.
3.2 miR-24对心血管疾病的影响
miR-24在心血管系统中广泛存在,能够抑制发育期和心肌梗死的血管生成,从而影响心血管的功能作用.miR-24是一种对缺氧敏感的miRNA,能够通过调节血管功能参与心血管疾病的发展.miR-24通过与Notch基因3′UTR结合靶向调节Notch以及其他一些血管形态因子参与调节肢体肌肉缺血后的微血管反应过程[23].miR-24-3p通过靶向BCL2L11,使冠心病患者外周血BCL2L11表达下调,抑制血管平滑肌细胞的凋亡,可作为治疗冠心病的潜在靶点[24].miR-24的上调表达通过PI3K/Akt/mTOR轴的PIK3R1,抑制血管平滑肌细胞增殖、迁移,显著减轻糖尿病大鼠血管重构[25].miR-24-3p可通过激活NRF2-Keap1信号通路,降低Keap1蛋白表达水平,达到保护心肌细胞免受缺血/再灌注损伤过程[26].miR-24参与糖尿病的血管重构过程,miR-24通过参与NLRP3相关炎症信号通路,抑制损伤动脉膜增生过程,从而减轻糖尿病大鼠的血管重塑过程[19].miR-24在血清中的不同表达水平也与心血管疾病进展不同时期相关,如miR-24、miR-23a和miR-145a在急性心肌梗死后的急性肾损伤患者的血清中表达水平显著下调,提示这3种miRNA可以作为早期发现急性心肌梗死的标记基因.
3.3 miR-24在脂质代谢中的功能作用
miRNA影响脂质代谢过程, 主要通过参与调控前脂肪细胞分化、肝脏脂质和血浆中脂类的合成等过程.在影响脂肪分化形成上通过不同的调节作用起到促进或抑制的作用,文献[27]研究发现过表达miR-24可以抑制MAPK7,增加PPAR-γ、SREBP1基因的表达,使脂滴形成和甘油三酯的积累增加,从而显著促进3T3-L1的脂肪生成.而miR-23a~27a~24基因家族通过同时靶向促脂肪(GPAM、DGAT2)和抑脂肪(DCN、G6PD、LPL)相关基因调控牛脂肪细胞生成过程,最终基因家族的平衡调节作用抑制脂肪的生成.miR-24的过表达下调与脂肪生成有关的某些基因(FASN、ACLY和SCD1)表达,但增加胆固醇合成、脂质合成相关基因(HMGCR、DHCR24和SREBP2)的表达,其功能的发挥主要是通过靶向基因SR-B1实现[28].miR-24同时影响脂质代谢相关产物的生成,在脑梗死大鼠中miR-24能够降低血清中总胆固醇、高密度脂蛋白以及甘油三酯水平,进而抑制脑组织的凋亡[29].miR-24在高脂小鼠的肝脏和离体的人肝细胞中表达水平显著增加,其作用机制主要是通过抑制Insig1促进肝脏脂质沉积和高脂血症,miR-24被敲除后将导致肝脏脂质积聚受损,血浆甘油三酯降低,提示miR-24可作为动脉粥样硬化的潜在治疗剂[30].在动物生产上的研究发现miR-24参与了兔肝细胞中多不饱和脂肪酸的生物合成过程[31].在山羊乳腺组织泌乳期和干乳期的表达水平显著不同,体外实验表明miR-24通过靶向FAS影响甘油三酯含量、调控SREBP1等基因的表达从而影响乳汁中脂肪酸含量[32].
3.4 miRNA-24对卵巢及卵泡生长的调控
miRNA参与卵巢发育的整个生物学功能,包括卵泡生长、闭锁以及排卵等一系列过程中的因子调控功能,卵泡生长各个阶段受特异性表达的miRNA调控.miR-24也参与繁殖过程中的各个途径,人卵泡液中的miR-24-3p能够调控雌二醇和孕酮的水平,降低睾酮水平[33].miR-24-3p能够靶向SMAD5通过FasL-Fas信号通路调控人卵泡颗粒细胞凋亡水平[34].miR-24的表达水平与牛卵泡颗粒细胞发育相关,影响受精率,与山羊的妊娠状态相关等.miR-24-3p还能够降低HepG2细胞脂质沉积能力,而脂质沉积影响了鸡卵泡生长过程.miR-24-3p在30周龄的鸡的肝脏中表达水平下降[35],表明其有可能参与了卵黄脂质合成及代谢,影响卵黄沉积等过程,从而影响卵泡生长.
3.5 miR-24在其他生物过程中的作用
miR-24除了影响细胞增殖、凋亡等过程,也影响如少突胶质前体细胞、神经元、胚胎干细胞等分化过程.文献[36]研究发现miR-24在损伤的脊髓细胞中表达水平增加,并通过靶向调节ADM抑制少突胶质细胞前体细胞的分化.文献[37]研究发现miR-24-3p通过控制海马钙蛋白的表达来调节神经元的分化进展.文献[3]研究发现BMP4通过募集SMAD5调控miR-23a/b~27a/b~24启动子序列,使miR-23a/b~27a/b~24影响胚胎干细胞的分化功能.
miR-24也参与机体内炎症反应和氧化应激等生物过程,并通过靶向基因的功能作用作为疾病治疗靶点.文献[38]将外源性miR-24-3p转染进缺血再灌注小鼠的肝脏中,发现miR-24-3p可显著抑制肝脏内STING和p-IRF3蛋白的表达,从而达到减轻炎性细胞因子的释放,并降低肝细胞功能障碍和凋亡过程.文献[39]研究发现抑制类风湿关节炎的GSK-3β抑制剂TDZD-8能够通过下调miR-24表达,降低类风湿关节炎的炎症反应过程,说明miR-24促进类风湿关节炎的发生.miR-24在溃疡性结肠炎患者的结肠活检和血液样本中比健康对照组的表达水平要高,进一步研究提示miR-24是肠道免疫屏障的重要调节因子[40].在氧化应激上研究显示miR-24在人体外胸主动脉平滑肌细胞的上调,可以通过NRF2/HO-1信号通路抑制细胞内ROS、MDA等的活性水平,从而抑制细胞的氧化应激反应[41].
4 展望
综上所述,miR-24的调控是一个复杂的系统,其表达水平受到转录因子和非编码RNA等多方面调控,其功能发挥主要通过与不同靶基因结合广泛地参与细胞增殖、分化、凋亡等过程.研究表明miRNA的生成机制和作用靶基因在各物种或各组织中表现差异性,因此进一步研究miR-24在不同物种或组织中的生物合成的分子机制及其调控作用,将为miR-24家族成员的生物学功能提供新的见解.然目前的研究多集中于在人、小鼠物种,相关组织也多与肿瘤相关,大部分的miR-24的靶基因及其相关功能仍未被解析验证,仍存在许多科学问题有待于深入研究,全面剖析miR-24的功能作用仍有很长的路要走.
