80份棉花种质资源的育种应用价值评价
2021-12-12李兴河王海涛刘存敬唐丽媛张素君熊永斌张香云
李兴河,王海涛,刘存敬,唐丽媛,张素君,蔡 肖,熊永斌,张香云
(1河北省农林科学院棉花研究所/农业农村部黄淮海半干旱区棉花生物学与遗传育种重点实验室/国家棉花改良中心河北分中心,石家庄 050051;2河北省农林科学院,石家庄 050051)
0 引言
中国是世界上的产棉和纺织大国,棉花产业发展影响国民经济和外汇储备[1],主要植棉区农业人口达5 亿,劳动力人口2 亿,植棉收入是一些地区的主要经济来源[2]。广泛收集优异种质资源并进行生物学鉴定及遗传分析是棉花分子育种的任务之一[3],而且,种质资源的引进、评价与利用是遗传改良和不断培育新品种的重要基础,种质资源的遗传多样性对现有资源的改良[4]、维护物种稳定[5]以及亲本选配[6]非常重要。在中国棉花育种的历史进程中,一直注重种质资源的引进,最初引自美国等国家的几个重要品种如‘金字棉’、‘斯字棉’、‘岱字棉15’和‘乌干达棉’等,经育种家加以利用和改造,逐步发展成中国主栽陆地棉品种[7]。
了解和掌握棉花种质资源特征特性是选择亲本的重要前提[8],前人对国内外棉花种质资源做了大量遗传评价。王沛政等[9]从生产上选取20个陆地棉品种进行主成分分析和聚类分析,受样本数量和来源的局限,并未说明亲缘关系和地域之间的联系,故而不能在陆地棉中引伸;董承光等[10]基于表型性状对429 份陆地棉种质资源进行综合评价,通过聚类分析发现类群间差异明显,但并未发现聚类结果与地理来源的关系;董灵艳等[11]对山东省审定的37 个中早熟棉花品种的12个数量性状进行遗传多样性分析、主成分分析与聚类分析,发现供试品种亲缘关系密切,遗传基础狭窄,不适合同时作为育种亲本;徐敏等[12]分析了144 份种质资源的产量、品质和抗病性情况,指出产量性状、纤维品质和抗病性相关性不密切;戴茂华等[13]对黄河流域棉区的30份棉花品种的12个代表性状进行变异性、相关性和聚类分析,发现冀中南地区影响棉花皮棉产量的主要因素为单株铃数和衣分,并且指出产量与纤维品质间的负相关性在逐渐减弱。董承光等[14]对153份陆地棉种质资源的主要农艺性状进行综合评价,并通过聚类分析筛选出33份可作为育种亲本的种质资源;陈光等[15]分析了43 份陆地棉基础种质资源的遗传多样性,发现基础种质间在产量、品质、农艺性状上差异显著或极显著,提出环境适应、自然选择和人工选育造成了较为丰富的表型多样性的可能。目前,将分子标记应用于种质资源遗传评价的报道屡见不鲜,徐秋华等[16]利用RAPD 标记从分子水平上对51 个抗枯萎病陆地棉品种进行了遗传多样性分析,相似系数小于0.500 的仅占16%,总体相似性较高;刘文欣等[17]将建国以来中国代表性的166个棉花主栽品种进行分子标记分析,表明不同时期、不同种植区域和不同来源的棉花品种遗传差异明显,但整体遗传基础狭窄;温小杰等[18]利用分子标记对中国不同年代培育的54 个棉花品种进行遗传差异分析,指出中国抗枯、黄萎病品种主要来源于岱字棉和斯字棉血统,阐明了国内不同棉花品种来源的局限性;卫泽等[19]通过表型聚类和SSR 分子标记聚类研究了国内外57份棉花种质,指出同一国家遗传差异小、国家间遗传差异大的地域差异。宋宪亮等[20]从系谱分析、形态特征、生化及DNA 分子水平等方面分析了棉花遗传多样性研究进展,阐述了陆地棉品种间遗传多态性水平低并指出加强现有品种的研究利用和种质资源的交流是提高育成品种多样性的重要途径。
虽然前人在棉花种质资源评价方面做了大量工作,但大多以本土尤其是当地棉花品种(系)为主,造成陆地棉品种间遗传多样性不够丰富,遗传基础狭窄;同时,对种质资源的优缺点分析不够,难以真正筛选出优异的种质资源。针对以上问题,本研究选取80份来源广泛的棉花种质资源,针对4 个产量性状(单株铃数、铃重、衣分、子棉产量)、5 个纤维品质性状(上半部平均长度、断裂比强度、马克隆值、伸长率、整齐度)、1个抗病性状(黄萎病病指)进行遗传多样性分析、相关性分析、主成分分析以及系统聚类分析,分析种质资源的优势和不足,筛选适于棉花育种工作的优异资源,以期扩充育种资源圃,为种质资源创新、品种遗传改良提供资源基础。
1 材料与方法
1.1 材料来源
供试的80 份棉花种质资源均引自山东棉花研究中心资源库。资源名称及来源见表1。
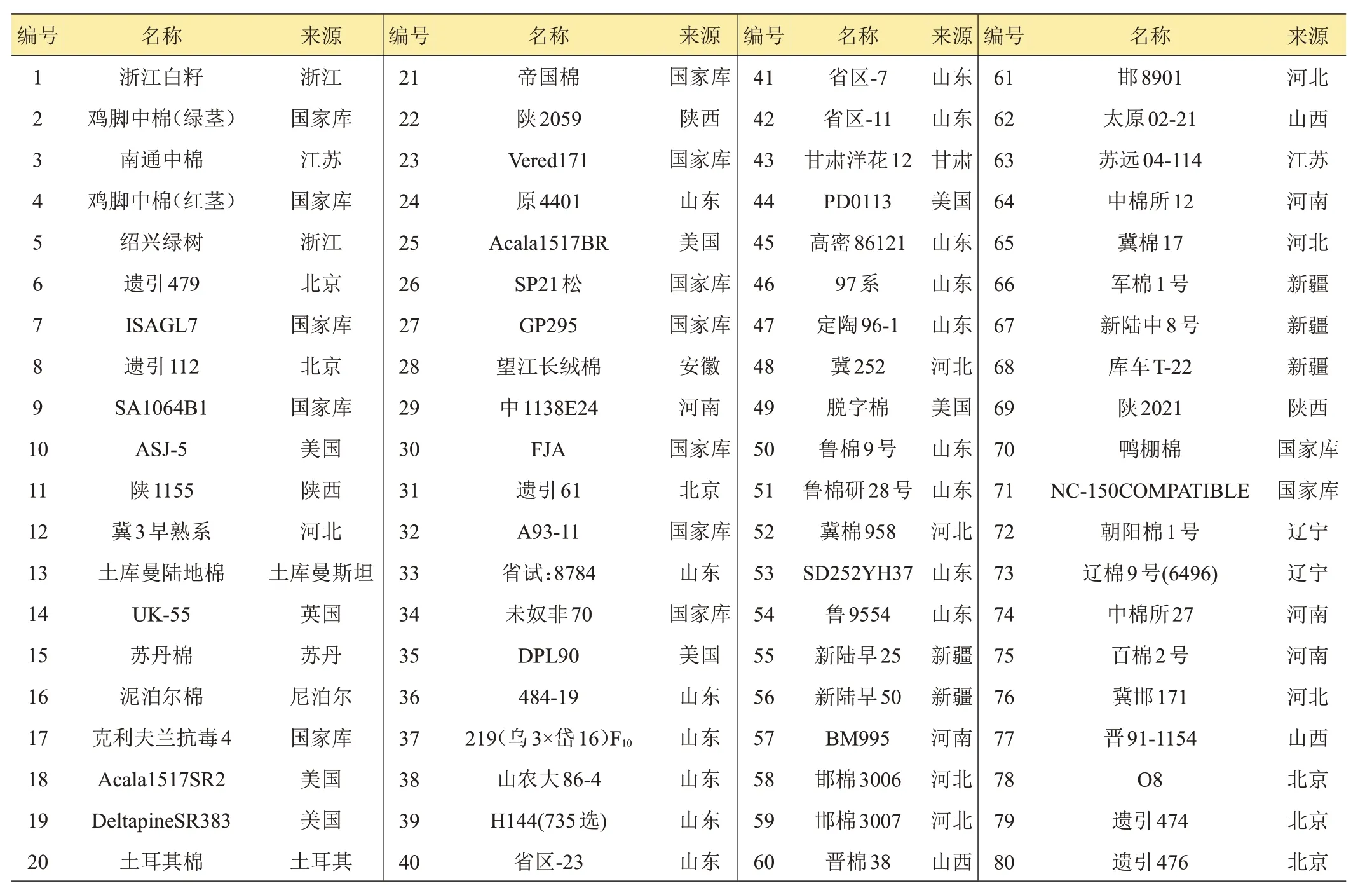
表1 供试种质资源编号及来源
1.2 试验方法
1.2.1 种植方式2019年,供试材料种植于河北省农林科学院棉花研究所石家庄市小安舍试验站黄萎病病圃。试验采取随机区组设计,3次重复,每个小区种植3 行,行长7.5 m,行距0.8 m,株距0.3 m,常规试验管理。
1.2.2 性状调查 针对目前棉花生产中较为关注的10个性状进行考查:8—9 月份,调查株高、果枝数、单株铃数和黄萎病发病情况,并采用相对病情指数(relative disease index,RDI)表示各个材料的抗病性强弱[21];10月份,每个小区选择正常吐絮单株收取中部棉铃30 个,用于考查衣分、铃重,纤维上半部平均长度、断裂比强度、马克隆值、伸长率和整齐度委托国家棉花纤维检验重点实验室(河南安阳)进行检测;11月份全部收获后测产。
以上所有性状全部采用3个重复的平均值。
1.2.3 数据整理及软件分析 通过Microsoft Excel 2007整理试验数据,采用李晶等[22]的计算方式计算遗传多样性指数,利用DPS 15.10进行性状相关分析、性状主成分分析和系统聚类分析。
2 结果与分析
2.1 资源材料的遗传多样性分析
表2 为80 份资源材料的遗传多样性分析结果,可以看出,不同性状的变异系数范围为2.59%~30.97%。其中,子棉产量变异系数最大,为30.97%,变异幅度为401.60~6185.00 kg/hm2,整齐度变异系数最小,为2.59%。性状变异系数由大到小顺序为子棉产量>黄萎病病指>单株铃数>铃重>断裂比强度>马克隆值>衣分>上半部平均长度>伸长率>整齐度。从遗传多样性指数来看,单株铃数的遗传多样性指数最高,为2.05,整齐度的遗传多样性指数最低,为1.68,遗传多样性指数由大到小顺序为单株铃数>伸长率>衣分>断裂比强度>子棉产量>黄萎病病指>马克隆值>铃重>上半部平均长度>整齐度。

表2 供试种质资源性状遗传多样性分析结果
2.2 性状之间的相关性分析
表3 为80 份资源材料10 个表型性状的相关性分析结果,可以看出,各性状之间存在不同程度的相关性,部分为显著或极显著相关。相关性达到显著水平的性状组合:单株铃数与衣分、子棉产量呈极显著正相关;衣分与上半部平均长度呈显著正相关,与子棉产量、整齐度呈极显著正相关;铃重与断裂比强度呈显著正相关,与子棉产量、上半部平均长度、整齐度呈极显著正相关,与马克隆值呈显著负相关;子棉产量与黄萎病病指呈显著正相关,与上半部平均长度、整齐度呈极显著正相关,与马克隆值呈显著负相关;上半部平均长度与整齐度、断裂比强度呈极显著正相关,与马克隆值呈极显著负相关;整齐度与断裂比强度呈极显著正相关,与马克隆值呈极显著负相关;断裂比强度与马克隆值呈极显著负相关;马克隆值与黄萎病病指呈极显著负相关。相关性分析体现了性状之间的复杂相互影响,在育种利用价值评价时应综合分析。

表3 10个性状之间的相关性分析结果
2.3 资源材料的主成分分析
对80份资源材料进行主成分分析,提取特征值大于1 的4 个主成分,累积贡献率为76.893%,包含了调查性状的绝大部分信息(表4)。从表中可以看出,第1主成分特征值为3.369,贡献率为33.693%,其中上半部平均长度特征向量值正值最大(0.488),说明上半部平均长度对第1主成分影响最大,整齐度(0.465)、子棉产量(0.341)、断裂比强度(0.333)和铃重(0.297)的特征向量也相对较高,马克隆值特征向量为负值最大(-0.376)。因此,第1主成分主要与纤维品质有关,产量因素也起到较大作用。第2主成分特征值为1.932,贡献率为19.317%,其中单株铃数特征向量值正值最大(0.604),说明单株铃数对第2主成分影响最大,其次是子棉产量(0.508),因此,第2主成分主要与产量性状有关。第1主成分和第2主成分可合并作为纤维品质和产量因子。第3 主成分特征值为1.344,贡献率为13.444%,其中黄萎病病指特征向量值负值最大(-0.703),说明黄萎病病指对第3主成分影响最大,第3主成分可作为棉花抗病性因子。第4主成分的特征值为1.044,贡献率10.439%,其中伸长率特征向量值为正值最大(0.928),第4主成分可作为伸长率因子。
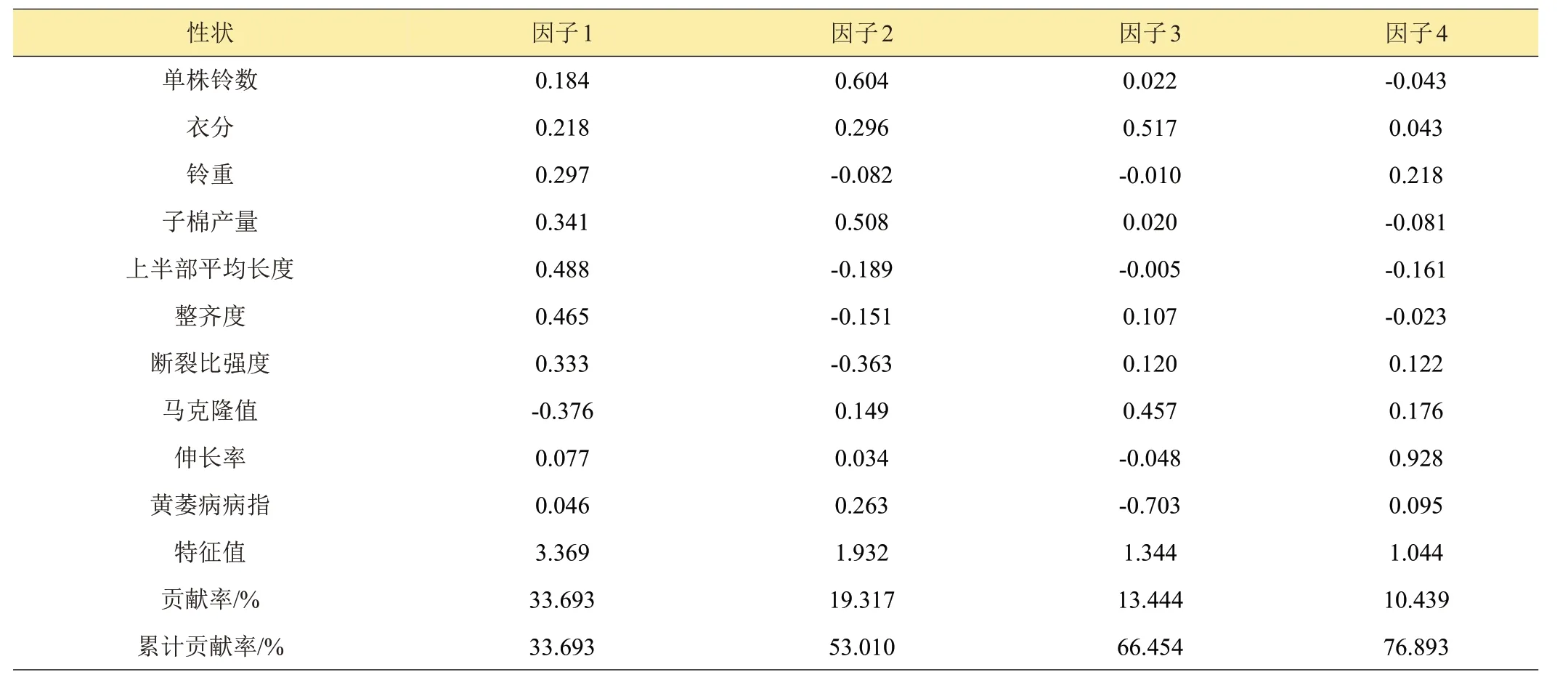
表4 供试材料主成分分析前4个主成分的因子载荷矩阵及特征值描述
参照周绍松[23]的分类方法,将供试棉花资源材料分为4类(图1):第Ⅰ类,以‘484-19’为代表,位于散点图的右上角,第1主成分和第2主成分得分均较高,表明产量性状和纤维品质性状表现协调且相对优异;第Ⅱ类,以‘FJA’为代表,位于散点图的右下角,第1主成分得分高且第2 主成分得分低,主要表现为纤维品质相对优异,产量性状有待改良;第Ⅲ类,以‘遗引479’为代表,位于散点图的左上角,第1主成分得分低且第2主成分得分高,普遍表现为产量性状良好,纤维品质性状有待改良;第Ⅳ类,以‘亚洲棉鸡脚中棉’(红茎)为代表,位于散点图的左下角,第1主成分和第2主成分得分均低,纤维产量和品质性状均表现较差。

图1 供试材料主成分分析散点图
2.4 资源材料的系统聚类分析
本研究采用卡方距离-可变类平均法进行系统聚类分析。依据10个表型性状在卡方距离D=2.15时,各性状p值均小于0.05,将80 份种质资源划分为6 个类群。聚类情况见图2,各类群的性状均值见表5。

表5 不同类群10个性状的平均值

图2 80份种质资源的聚类结果
各类群具体分析如下。第Ⅰ类群:包括22份种质资源,包括‘土库曼陆地棉’、‘尼泊尔棉’、‘陕2059’、‘原4401’、‘省试8784’、‘484-19’、‘省区-23’、‘省区-7’、‘省 区-11’、‘97 系’、‘鲁 棉研28 号’、‘冀棉958’、‘BM995’、‘邯棉3006’、‘邯棉3007’、‘晋棉38’、‘中棉所12’、‘冀棉17’、‘库车T-22’、‘陕2021’、‘中棉所27’、‘冀邯171’。该类群产量性状优异,单株铃数17.3,衣分41.24%,子棉产量5316.53 kg/hm2。其中,‘97 系’、‘484-19’、‘省区-23’、‘冀棉958’、‘晋棉38’的子棉产量均超过5825.09 kg/hm2,其他性状没有明显缺陷,可以直接用于改良现有品种的产量性状。
第Ⅱ类群:包括30 份种质资源,包括‘ISAGL7’、‘遗引112’、‘SA1064B1’、‘ASJ-5’、‘苏丹棉’、‘Vered171’、‘Acala1517BR’、‘GP295’、‘望江长绒棉’、‘中1138E24’、‘FJA’、‘A93-11’、‘DPL90’、‘山农大86-4’、‘H144(735 选)’、‘PD0113’、‘定陶96-1’、‘冀252’、‘鲁棉9号’、‘SD252YH37’、‘鲁9554’、‘新陆早25’、‘新陆早50’、‘邯8901’、‘苏远’‘04-114’、‘军棉1号’、‘晋91-1154’、‘O8’、‘遗引474’、‘遗引476’。该类群产量、纤维品质性状优异,子棉产量超过整体平均值17.87%,断裂比强度为各类群最高(32.43 cN/tex)。其中,‘山农大86-4’、‘邯8901’、‘苏远04-114’的子棉产量均超过5103.69 kg/hm2,超平均值45.88%以上,纤维品质均达到“Ⅱ型”,可以直接用于改良现有品种的产量和品质。
第Ⅲ类群:包括9份种质资源,包括‘陕1155’、‘冀3早熟系’、‘UK-55’、‘克利夫兰抗毒4’、‘土耳其棉’、‘帝国棉’、‘甘肃洋花12’、‘脱字棉’、‘新陆中8 号’。该类群优点是铃大、纤维伸长率高;缺点是抗黄萎病能力差,可用于棉花铃重、纤维伸长率等方面的改良。
第Ⅳ类群:包括12 份种质资源,包括‘遗引479’、‘Acala1517SR2’、‘Deltapine SR383’、‘SP21 松’、‘遗引61’、‘219(乌3×岱16)F10’、‘高密86121’、‘太原02-21’、‘NC-150 COMPATIBLE’、‘朝阳棉1 号’、‘辽棉9号(6496)’、‘百棉2 号’。该类群龄大、衣分高。其中,‘Acala1517SR2’铃重达到8.0 g,是典型的大铃材料,可以用于改良棉花品种的铃重,间接提高棉花产量;‘遗引61’衣分达到45.88%,可以用于棉花高衣分品种培育。
第V 类群:包括2 份种质资源,包括‘未奴非70’、‘鸭棚棉’。其中,‘未奴非70’的纤维品种优异(上半部平均长度34.24 mm,断裂比强度34.70 cN/tex,马克隆值4.81),黄萎病病指为0,但是子棉产量仅为1875.2 kg/hm2,难以直接利用,可通过构建回交群体、RIL 群体等方式进行资源创新、优异性状QTL 定位等研究。
第Ⅵ类群:包括5 份种质资源,包括‘浙江白籽’、‘鸡脚中棉(绿茎)’、‘南通中棉’、‘鸡脚中棉(红茎)’、‘绍兴绿树’,全部为亚洲棉,产量性状和纤维品质性状均最差,难以直接应用于棉花育种。
3 讨论与结论
本研究考查了80份种质资源的10个性状,包含产量、纤维品质、抗病性3 个方面。遗传多样性分析显示,4 个产量性状变异系数均大于10%,其中单株铃数、铃重、子棉产量的变异系数均大于20%,而5 个纤维品质性状变异系数最大仅为10.83%,其中上半部平均长度、整齐度、伸长率变异系数均小于10%,变异程度小,说明纤维品质性状的变异程度低于产量性状,这与代攀虹等[24]研究结果一致,不同来源的种质资源具有相同的性状变异规律,说明纤维品质性状相对稳定,产量性状可能更易受环境、气候等因素的影响。
性状相关性分析中,单株铃数与衣分、子棉产量,衣分与整齐度,铃重与子棉产量、上半部平均长度、整齐度,上半部平均长度与断裂比强度,整齐度与断裂比强度均呈极显著正相关,表明单株铃数越多,衣分越高,子棉产量越高;衣分越高,整齐度越高;铃重越重,子棉产量越高,上半部平均长度越高,整齐度越高;上半部平均长度越长,断裂比强度越大;整齐度越高,断裂比强度越大。上半部平均长度与马克隆值,整齐度与马克隆值,断裂比强度与马克隆值;马克隆值与黄萎病病指均呈极显著负相关,表明上半部平均长度越长,马克隆值越小;整齐度越高,马克隆值越小;断裂比强度越大,马克隆值越小;马克隆值越大,黄萎病病指越低。以上结果与李武等[8]、董承光等[10,14]、徐敏等[12]、戴茂华等[13]、代攀虹等[24]、李慧琴等[25]的部分研究结果一致。分析表明,4个产量性状间、4个纤维品质性状(除伸长率)间以及部分产量性状与部分纤维品质性状间均表现为极显著的相互促进关系,说明产量和纤维品质存在同步改良的可能;马克隆值与黄萎病病指的相关关系表明,纤维品质与黄萎病抗性的改良存在相互制约的可能。
主成分分析是多元统计分析的重要统计方法。主成分因子之间相关性不显著且容易直观分析[26]。提取贡献率达到76.893%的4个主成分因子,贡献率由大到小依次是:棉花纤维品质因子>棉花产量因子>棉花抗病性因子>伸长率因子。图1中,‘484-19’、‘冀棉958’、‘中1138E24’、‘省区-11’、‘山农大86-4’等右上角的资源材料,棉花纤维品质和产量因子得分都比较高,可作为首选种质资源进行改良;同时,前三个因子贡献率累积达到66.454%,该结果体现了引种时的主要关注点:首先是纤维品质(贡献率33.693%),其次是产量(贡献率19.317%)和抗病性(贡献率13.444%),这与目前棉花育种的主要关注点[3]是一致的。
系统聚类分析是目前应用最广泛的聚类分析方法,具有数值特征的变量和样品即可进行系统聚类分析[27]。结合图2、表4 和表5,6 个类群主要从主成分分析中的第1主成分和第2主成分的差异性方面进行划分,同时也体现出了棉花种质资源的地域因素。第Ⅰ类群包含22 份材料,占供试材料的27.5%,其中19 份为黄河流域的已审定品种和参加黄河流域省区试优质材料,地域普遍较近;第Ⅱ类群包含30份材料,在地域上分布广泛;第Ⅰ、第Ⅱ类群都有突出的优点,没有明显的缺点,可以直接改良棉花品种。第Ⅲ、第Ⅳ、第V类群优缺点明显,难以直接改良棉花品种,可以通过杂交,轮回选择等育种方法,结合分子辅助选择技术,聚合优异性状进行新品种培育。第Ⅵ类群中,‘鸡脚中棉(红茎)’和‘鸡脚中棉(绿茎)’亲缘关系相近,但从田间表现来看,2个材料在茎秆颜色上存在明显差异(红与绿),可作为研究茎杆颜色的材料加以利用。
前人在种质资源评价中,普遍将主成分分析[23]和聚类分析[13-14]单独分析讨论,进而筛选出符合要求的亲本材料。董灵艳等[11]提出了主成分分析和聚类分析相结合的观点,但未见具体报道。本研究将主成分分析散点图中的第I 类群和聚类分析的第Ⅰ、第Ⅱ类群取交集,获得25份资源材料:‘陕2059’、‘原4401’、‘484-19’、‘省区-23’、‘省区-7’、‘省区-11’、‘97 系’、‘冀棉958’、‘BM995’、‘邯棉3006’、‘邯棉3007’、‘晋棉38’、‘中棉所12’、‘陕2021’、‘冀邯171’、‘中1138E24’、‘DPL90’、‘山农大86-4’、‘PD0113’、‘定陶96-1’、‘冀252’、‘鲁棉9号’、‘SD252YH37’、‘新陆早50’、‘军棉1号’,以上材料在2种分析方法中均被划分到高产、优质类群,同时结合遗传多样性分析和性状相关性分析,结果更加客观、准确。
主成分分析散点图和聚类分析相结合的分析方法并非在任何情况下都适用。首先,主成分因子与育种目标性状不一致时不可用;其次,聚类分析未将含有育种目标性状的种质资源划分在同一类群时不可用;第三,主成分分析散点图划分的优势类群和聚类分析的优势类群没有交集时不可用。此外,仅仅基于表型的资源评价容易受环境等外在因素的影响,难以做到精准评价。今后可将分子标记等现代生物技术手段与常规方法结合进行资源评价[28-30],可更加精准的评价种质资源的育种利用价值,为种质资源利用提供有益信息。
综上所述,本研究中80份资源具有丰富的遗传多样性,尤其在单株铃数、铃重、子棉产量等产量性状尤为突出,为棉花产量改良提供了丰富的资源;产量性状和纤维品质性状有望同步改良,纤维品质性状与黄萎病抗性的改良存在相互制约的可能;筛选出可用作棉花育种亲本的25份高产、优质资源材料。
