黄藤Dof家族的全基因组鉴定及系统进化分析
2021-12-11张丹丹杨瑶君
张丹丹 杨瑶君 江 纳 付 春
(1. 乐山师范学院竹类病虫防控与资源开发四川省重点实验室,四川 乐山,614000;2. 乐山师范学院生命科学学院,四川 乐山,614000;3. 乐山师范学院数学与信息科学学院,四川 乐山,614000)
黄藤(Daemonorops jenkinsiana)是常绿攀援棕榈藤的代表,自然生长在海拔1 000 m以下的低地雨林,黄藤丛生生长,茎可达50 m,节间长40 cm,直径30 mm[1]。黄藤的果实可萃取药品“麒麟血竭”,具有止血、活血、化瘀、止痛等疗效,嫩茎梢17种氨基酸的总含量高达13.14%,营养丰富,可供食用,具有重要的开发价值[2-3],是热带森林重要的再生非木材资源,但长期的森林采伐和药材采集,使黄藤资源量大幅下降[1]。为了缓解及保护现有的资源状况,一些国家建立了藤种基因收集圃、藤种园等迁地保存基地;同时人们开始了对现有资源的研究利用,特别是在棕榈藤的良种选育方面[4],目前国内外对黄藤育种的系统研究还比较少。
Dof(DNA binding with one finger)蛋白是一类植物特异性转录因子。Dof基因家族属于单锌指蛋白超家族,一般由200~400个氨基酸构成[5]。Dof蛋白具有明显的结构特征,由N−末端高度保守的Dof结合结构域、寡聚化位点、引导Dof蛋白进入细胞核的核定位区段以及负责C−末端转录调控的激活结构域4个部分组成[6]。其中,Dof结合结构域是由52个保守的氨基酸残基构成的C2−C2型单锌指结构域,在该结构域中有4个绝对保守的Cys残基[7]。Dof蛋白由于其C−末端转录调控域的氨基酸序列较为多变,不具有保守性,因此在植物中的功能具有多样性[8]。目前对玉米(Zea mays)[9]、拟南芥(Arabidopsis thaliana)、番茄(Solanum lycopersicum)[10-11]、水稻(Oryza sativa)[12]、桃(Amygdalus persica)[13]、草莓(Fragaria ananassa)[14]、香蕉(Musa nana)[15]、毛竹(Phyllostachys heterocycla)[16]、大豆(Glycine max)[17]、甘蓝型油菜(Brassica napus)[18]、小麦(Triticum aestivum)[19]的Dof基因家族成员研究表明,Dof基因家族成员参与到植物体的多种生命活动中,对植物生长发育、光调控、防御机制以及激素调控等过程中的相关基因有重要的调控作用[2],值得注意的是,关于Dof蛋白的克隆和功能研究主要集中在拟南芥、番茄、水稻等模式植物中,在棕榈藤物种中尤其在黄藤中鲜有报道。
目前,有关黄藤的研究还比较缺乏,仅在黄藤栽培、人工林培育与经营和种子分子标记等方面有过报道[20-22],但关于黄藤Dof基因家族的理化性质、氨基酸组成、亲疏水性、亚细胞定位、二三级结构等方面几乎没有报道。本研究利用生物信息学方法对黄藤全基因组中的Dof基因家族成员进行鉴定和分析,同时与拟南芥等其他9种物种基因组中的Dof家族成员进行同源比对并构建系统进化树,拟探讨藤本植物(黄藤)基因组中Dof基因家族与其他物种的进化起源关系,为今后进一步研究黄藤Dof基因家族的功能和黄藤育种提供参考依据。
1 材料与方法
1.1 数据获取
黄藤(Daemonorops jenkinsiana)全长基因组序列和其蛋白序列来自于国际竹藤中心公布的2种藤本植物基因组数据于GIGADB(http://gigadb.org/dataset/101053)[23],采用拟南芥数据库TAIR(https://www.arabidopsis.org/)下载拟南芥(Arabidopsis thaliana)Dof基因家族序列和蛋白序列,在植物转录因子数据库 PlantTFDB(http://planttfdb.cbi.pku.edu.cn/)下载毛竹(Phyllostachys heterocycla)、水稻(Oryza sativa)、二穗短柄草(Brachypodium distachyum)、小麦(Triticum aestivum)、玉米(Zea mays)、烟草(Nicotiana tabacum)、甘蓝型油菜(Brassica napus)和可可(Theobroma cacao)的Dof基因家族序列和蛋白序列。
1.2 黄藤Dof基因家族的鉴定
先检索下载pfam数据库(http://pfam.xfam.org/)中Dof基因家族的pfam模型号(PF02701),利用hmmsearch运行命令(hmmsearch --domtblout DjDof.txt --cut_tc zf-Dof.hmm(PF02701)Daemonorops jenkinsiana_protein.fasta)对黄藤蛋白质序列中的Dof基因家族进行预测分析,所得序列为黄藤Dof基因家族的候选蛋白质序列;再利用BlastP比对工具(参数:e值小于10−5)将黄藤中Dof基因家族的候选蛋白质序列与拟南芥36个Dof蛋白的氨基酸序列进行对比,剔除不相关的蛋白质序列后即为黄藤基因组中的Dof基因家族的蛋白质序列,共鉴定出25个黄藤Dof蛋白家族成员。
1.3 分析方法
1)黄藤Dof基因家族的理化特性分析。ExPASy-ProtParamtool(https://web.expasy.org/protparam/)[24]程序分析预测黄藤Dof基因家族编码所有蛋白质的分子量、等电点、亲疏水性、氨基酸组成等理化特性。
2)氨基酸的跨膜结构预测分析。利用TMHMMSeverv.2.0(http://www.cbs.dtu.dk/services/TMHMM/)在线程序对黄藤Dof蛋白成员进行跨膜结构域预测分析[25]。
3)亚细胞定位分析。利用在线软件CELLOv2.5[26](http://cello.life.nctu.edu.tw/)对25条黄藤Dof蛋白序列进行亚细胞定位分析。
4)蛋白的二、三级结构预测分析。利用SOPMA[27]网站对黄藤基因组中的Dof基因家族编码的所有蛋白质的二级结构进行预测分析。利用 SWISS−MOOEL[28]在线网站对其蛋白三级结构进行预测分析。
5)蛋白质的保守基序分析。利用 MEME 7.0.26[29](http://meme-suite.org/tools/meme)在 线工具对黄藤基因组中的Dof基因家族编码的氨基酸序列的保守基序进行预测分析。参数设置:保守基序个数设置为10,其他参数设置为默认。
6)基因结构分析与染色体定位分析。利用基因结构显示系统GSDS(Gene Structure Display Server)(http://gsds.cbi.pku.edu.cn/)[6]进行基因结构分析。根据黄藤Dof基因在染色体上的具体位置,使用MapChart软件绘制黄藤基因组中Dof基因家族成员的染色体物理图谱。
1.4 系统进化树的构建
利用 MEGA7.0.26[30]软件对黄藤Dof基因家族编码的蛋白质序列进行多重比对和系统进化树构建分析,构建黄藤与拟南芥基因组中的Dof基因家族的系统进化树以及构建黄藤Dof基因家族与拟南芥等其他9个物种的系统进化树。构建方法采用邻接法,校验参数设置为 Bootstrap 重复1 000次。
2 结果与分析
2.1 黄藤全基因组Dof基因家族的鉴定及其基本信息分析
黄藤Dof蛋白家族的理化特性分析结果表明,黄藤Dof蛋白质成员中氨基酸数目最多的是Dj-Dof20蛋白,有586个氨基酸,最少的为DjDof22蛋白,有184个氨基酸;脂肪族氨基酸指数在49.47~74.51之间;10个成员带负电荷的残基总数多于带正电荷的残基总数,有2个成员正负残基总数相等(DjDof24,DjDof22),13个成员正电荷残基总数多于负电荷总数(表1)。
蛋白质的分子量在20.12~62.42 kDa之间,既含有酸性蛋白又含有碱性蛋白,且碱性蛋白比酸性蛋白多,有11个成员的等电点大于8.0,最大等电点为9.57(DjDof17),最小等电点为4.71(DjDof18);不稳定系数显示,所有的蛋白均为不稳定蛋白(不稳定系数<40时稳定);黄藤Dof蛋白均为亲水性蛋白;亚细胞定位结果显示所有黄藤Dof基因家族成员都定位在细胞核上,(表1);跨膜结构预测结果显示黄藤Dof蛋白都不存在跨膜结构域,即都是非跨膜蛋白。
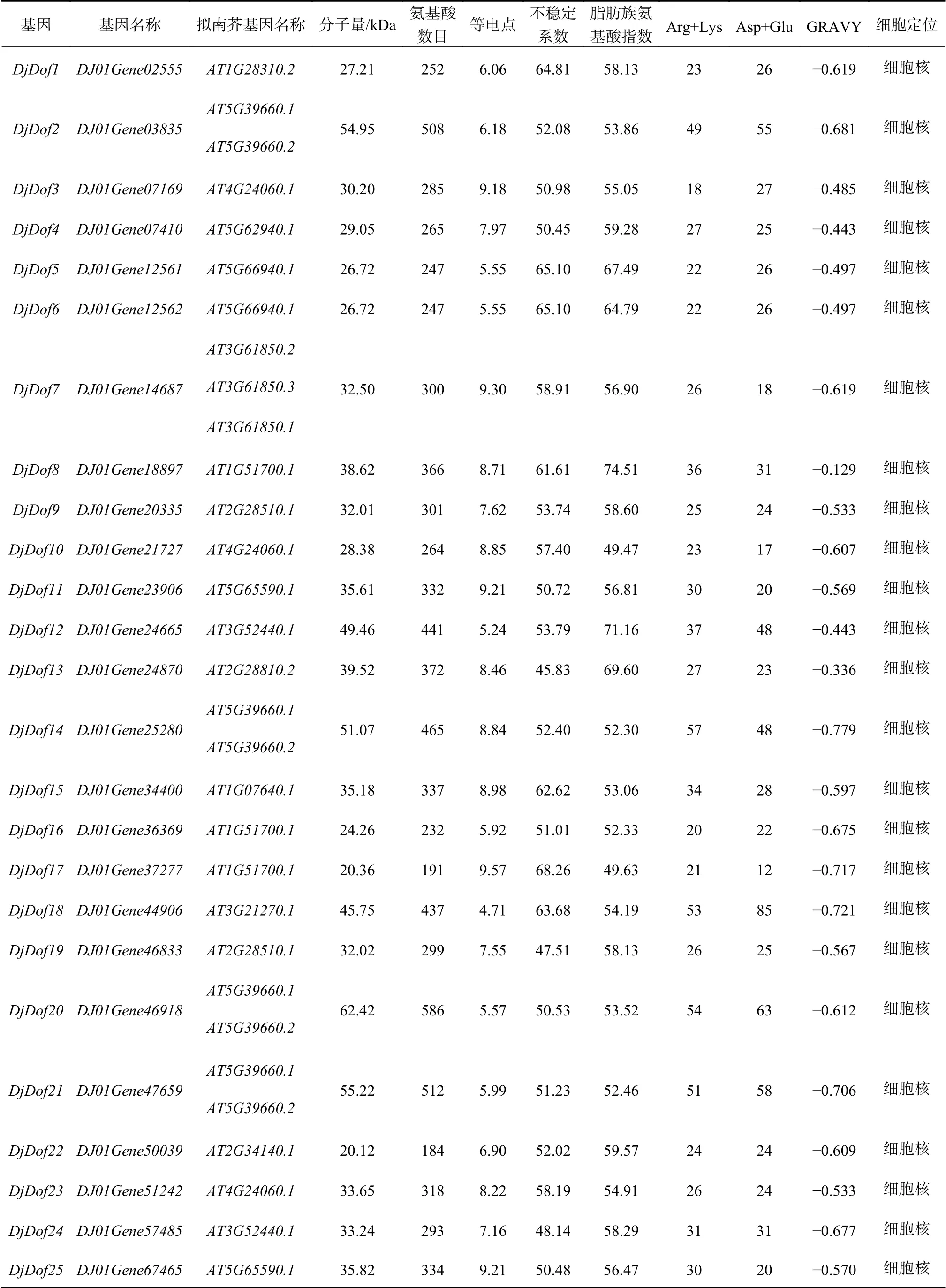
表1 黄藤Dof蛋白基本信息Table 1 Basic information of the Dof protein in D. jenkinsiana
2.2 编码蛋白的结构分析
2.2.1 编码蛋白的二级结构分析
蛋白质的二级结构主要是指蛋白质的多肽链中有规则重复的构象,从黄藤Dof基因家族成员的蛋白二级结构分析可以看出,其基因家族的二级结构主要为β转角、α螺旋、无规则卷曲、延伸链组成,4种蛋白质的二级结构通常被主要研究[6]。通过预测和分析将有助于认识蛋白的空间结构,其中无规则卷曲主要起连接其他二级结构元件。
同时蛋白二级结构预测结果显示:多数成员以α−螺旋和无规则卷曲为主要二级结构方式,多数成员二级结构的占比高低为 α−螺旋>无规则卷曲>延伸链>β−转角,α−螺旋在成员中占比范围为6.67%(DjDof7)~48.28% (DjDof18);无规则卷曲占比为39.36%(DjDof18)~83.33% (Dj-Dof7);延伸链占比为6.05%(DjDof21)~17.74%(DjDof4);β−转角占比为1.8%(DjDof25)~7.51%(DjDof24)(图1),综上所述,β−转角占比几乎是最少的,延伸链其次,说明β−转角和延伸链分散在整个氨基酸中。

图1 黄藤Dof蛋白二级结构分布Fig. 1 The secondary structure distribution of the Dof protein in D. jenkinsiana
2.2.2 编码蛋白的三级结构分析
利用 SWISS−MOOEL 在线网站分析了黄藤Dof蛋白的三级结构,再根据其构象进行分组。黄藤Dof蛋白可以分为11个组,以A−K表示组别,A组(DjDof7、DjDof8、DjDof12、DjDof13、DjDof17、DjDof22)成员最多有6个;其次为B组(DjDof15、DjDof16、DjDof18、DjDof19),有4个;C组(DjDof1、DjDof4、DjDof10)和D组(DjDof2、DjDof9、DjDof25)有3个;E组(DjDof5、DjDof6)和F组(DjDof3、DjDof23)有2个;其余G(DjDof14)、H(DjDof20)、I(DjDof11)、J(DjDof21)、K(DjDof24)组都有1个,其中组C、组D、组E和组F主要以α−螺旋、β−转角和β−折叠为主,组B、组G和组I主要以α−螺旋为主,组A主要以α−螺旋和β−折叠为主,组H主要以α−螺旋和β−转角为主,组J主要以β−转角为主(图2),该结果说明黄藤Dof蛋白存在保守性和差异性。
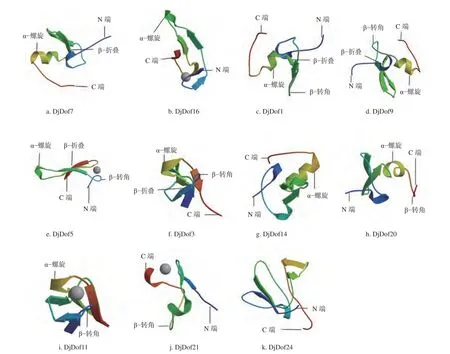
图2 黄藤Dof 蛋白3级结构Fig. 2 Tertiary structure of the Dof protein in D. jenkinsiana
2.3 氨基酸序列的保守基序分析
通过MEME对黄藤Dof蛋白保守基序进行了详细的 分析。结果表明,10个独立的保守基序被鉴定,命名为motif1~10,其中motif1为C2−C2单锌指结构,存在于所有的黄藤Dof蛋白中(图3)。该结果表明,在相同组中含有相同保守基序的Dof蛋白也许具有相似的功能。
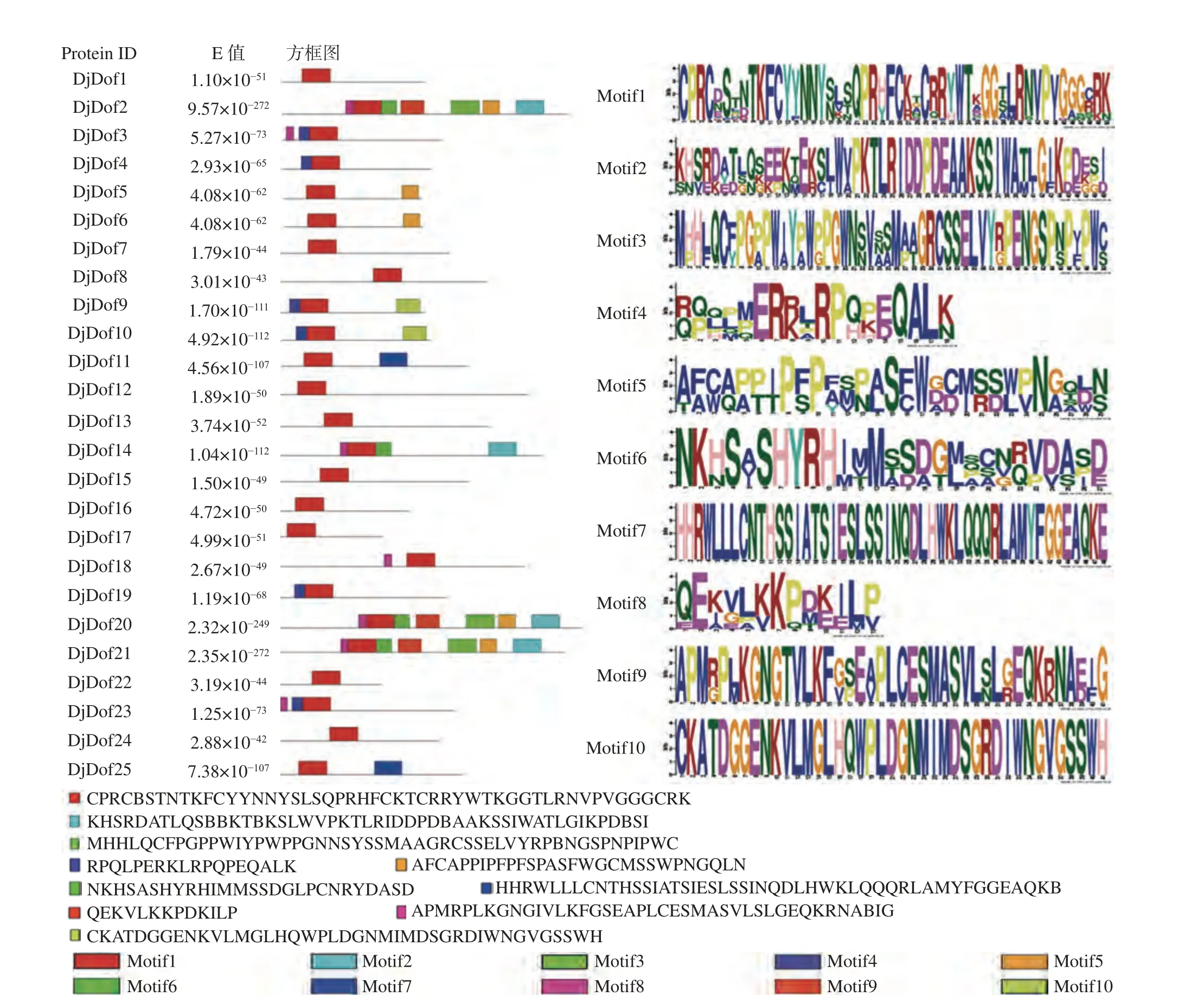
图3 黄藤Dof 蛋白保守基序分析Fig. 3 The conservative motif analysis of the Dof protein in D. jenkinsiana
2.4 黄藤Dof蛋白家族的进化关系分析
通过MEGA软件运用邻接法对来自黄藤的25个Dof蛋白家族进行进化树分析。根据树的分支可将其分为4组(ClassⅠ−ClassⅣ),分别是ClassⅠ−ClassⅣ,Ⅰ组内有5个成员,成员DjDof25和DjDof11进化速率最快,成员DjDof13的进化速率最慢;Ⅱ组内有8个成员,成员DjDof10和DjDof9的进化速率最快,其次是DjDof19,成员DjDof12和DjDof24的进化速率最慢;Ⅲ组内有5个成员,成员DjDof5和DjDof6的进化速率最快,其次是DjDof1,成员DjDof15的进化速率最慢;Ⅳ组内有7个成员,其中成员DjDof2和Dj-Dof21的进化速率最快,其次是DjDof20,成员DjDof22的进化速率最慢,其中ClassⅡ成员分化速度是最快的,与原始祖先的亲缘关系相对较远。从组别上来看,ClassⅡ成员分化速度是最快的,因而亲缘关系相对较远。从组中来看,ClassⅡ中成员DjDof9和DjDof10分化最快,亲缘关系相对较远,DjDof12和DjDof24分化较慢;ClassⅢ中成员DjDof5和DjDof6分化最快,亲缘关系相对较远,DjDof15分化较慢(图4)。此外,通过MEME软件中的ClustalW功能进行蛋白保守序列比对(图5),结果表明,黄藤Dof蛋白的结构域是高度保守的,且所有Dof蛋白的结构域均含有motif1,进而构成C2−C2单锌指结构,该结果进一步证明了所鉴定的蛋白为Dof家族的成员。

图4 黄藤Dof基因家族的系统发育树Fig. 4 The phylogenetic trees of the Dof gene family in D. jenkinsiana

图5 氨基酸序列比对Fig. 5 Amino acid sequence alignment
通过MEGA软件运用邻接法对来自黄藤的25个Dof和拟南芥的47个Dof进行进化树构建并分析。拟南芥ATDof2.3是复制特异性转录因子,缩短了细胞周期;过表达ATDof5.4通过促进内循环的早期发生,抑制细胞的扩增;ATDof2.4和ATDof5.8在拟南芥不同生长发育过程中发挥不同作用;ATDof6对种子萌发起负调控的作用。该结果表明,黄藤和拟南芥Dof基因家族的系统发育树分为5组,分别是ClassⅠ−ClassⅣ,Ⅰ组内包括黄藤Dof成员8个,拟南芥Dof成员13个;Ⅱ组内只有6个拟南芥Dof成员,不含有黄藤Dof成员,这表明黄藤Dof家族基因的形成迟于拟南芥的分化;Ⅲ组内包括黄藤Dof成员7个,拟南芥Dof成员11个;Ⅳ组内包括黄藤Dof成员和拟南芥Dof成员各4个(图6)。其中DjDof15在2个进化树中分布在2个不同的组中可能是因为在进化过程中发生了蛋白序列的改变。
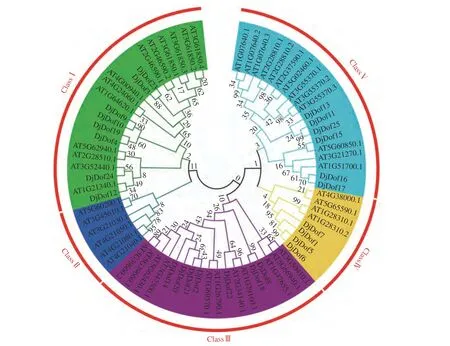
图6 黄藤和拟南芥Dof基因家族的系统发育树Fig. 6 The phylogenetic trees of the Dof gene family in D. jenkinsiana and A. thaliana
黄藤Dof基因家族成员与来自双子叶植物纲,单子叶植物纲和不同科的代表性植物拟南芥等多个物种的Dof成员分子进化关系分析表明,黄藤和其他9种植物Dof基因家族的系统发育树分为5组,分别是ClassⅠ−ClassⅤ,黄藤Dof蛋白成员主要分布在ClassⅤ,其次是ClassⅢ,ClassⅠ和ClassⅡ中不含有黄藤Dof蛋白成员,可能是黄藤在这2组中的同源基因在进化过程中丢失。亲缘关系越近,说明植物间的共同特征越多;亲缘关系越远,说明植物间的共同特征越少。其中,黄藤的Dof蛋白成员DjDof4和甘蓝型油菜的Dof蛋白成员BRNA2T00148796001、DjDof11和GSBRNA2T00127875001、DjDof12和GSBRNA2 T00141516001、DjDof19和GSBRNA2T00027470-001的亲缘关系最近,与可可的亲缘关系最远。从科分类的角度来分析,黄藤与十字花科的甘蓝型油菜的亲缘关系最近,与可可的亲缘关系最远;从双子叶植物与单子叶植物分类分析,黄藤与小麦的亲缘关系最近,亦是与可可的亲缘关系最远,可能与他们分别属于单双子叶植物有关(图7)。
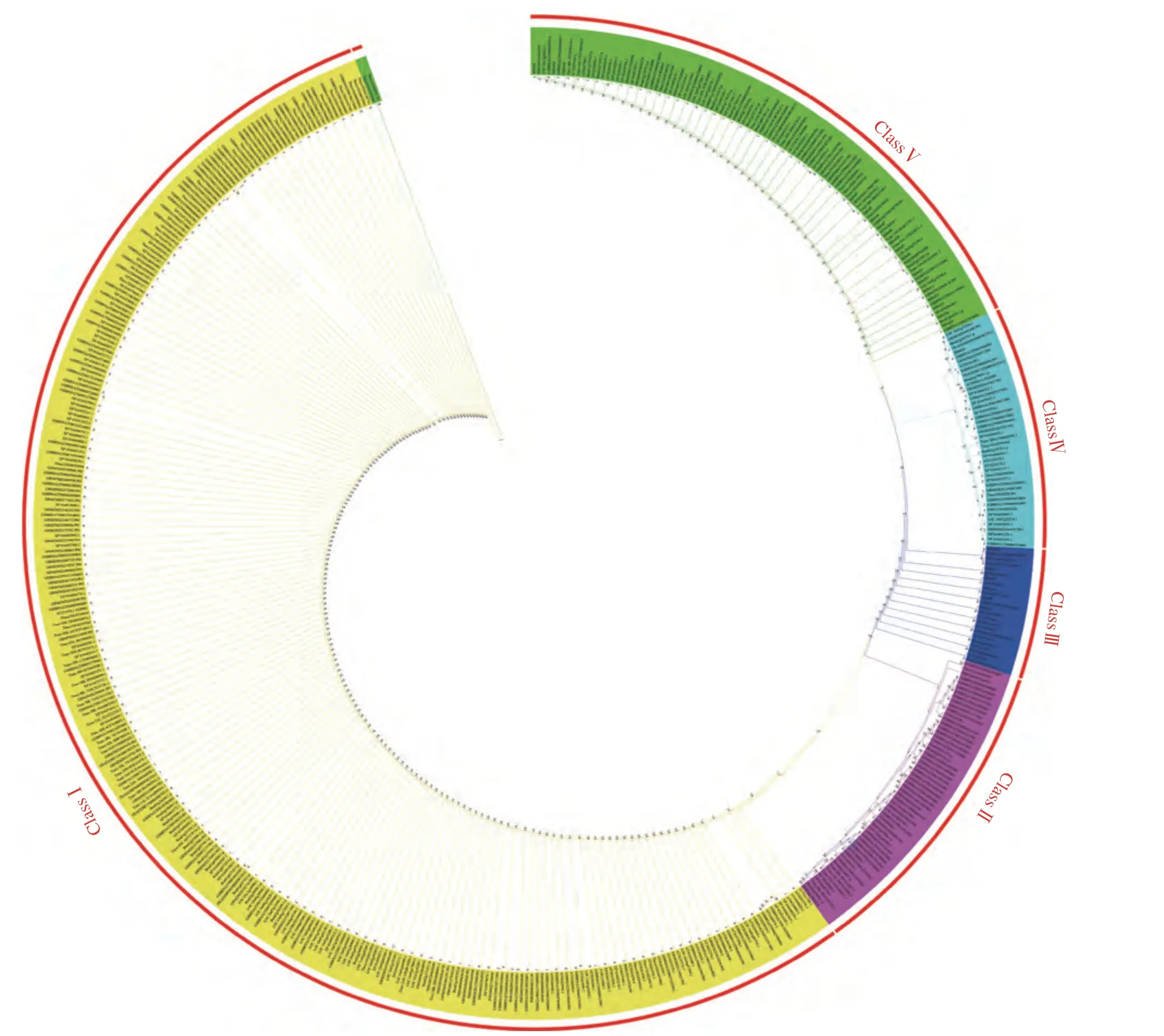
图7 黄藤和其他物种Dof基因家族的系统发育树Fig. 7 The phylogenetic trees of the Dof gene family between D. jenkinsiana and other species
2.5 黄藤Dof基因家族的结构及染色体定位分析
2.5.1 黄藤Dof基因家族的结构分析
通过在线网站GSDS2.0(gene strucyure display server)对黄藤Dof转录因子家族进行基因结构分析,结果显示黄藤Dof家族成员基因结构简单且相似性较高,均含有1~2个内含子,其中有20个DjDof基因只含有1个内含子,占总数的80%;其余的5条序列(DjDof4、DjDof10、Dj-Dof14、DjDof18、DjDof20)都含有2个内含子,占总数的20%。进1步分析发现,系统进化树属于同一分支的Dof基因,其基因结构也有明显差异,如图8 ClassⅣ中DjDof2、DjDof8、DjDof21、DjDof22只含有1个内含子,其余成员则包括2个内含子。可能是黄藤Dof基因家族在进化过程中相对保守。且各基因长度不等,相差较大。
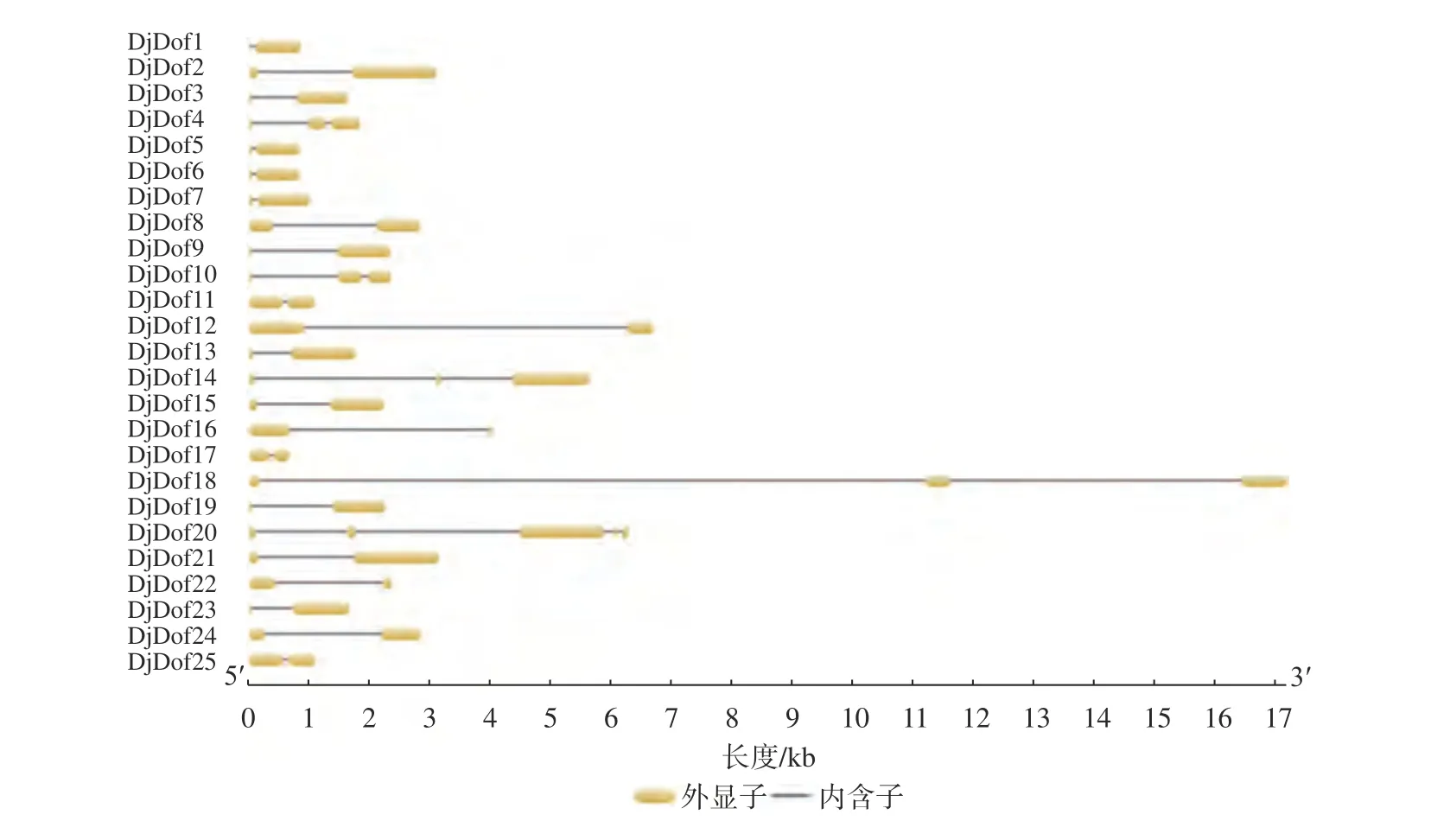
图8 黄藤Dof家族基因结构Fig. 8 Gene structure of Dof family in D. jenkinsiana
2.5.2 黄藤Dof基因家族染色体定位分析
通过染色体定位分析发现25个DjDofs基因不均匀分布于黄藤24条染色体中,未发现串联重复基因,只有第5条染色体上有2个DjDof基因的分布,其余的染色体都只有1个DjDof基因的分布。可能是黄藤的Dof基因在基因重组的过程中位置没有发生太大的变化(图9)。黄藤Dof基因家族中存在1对重复基因,DjDof5和DjDof6为1对串联重复基因。
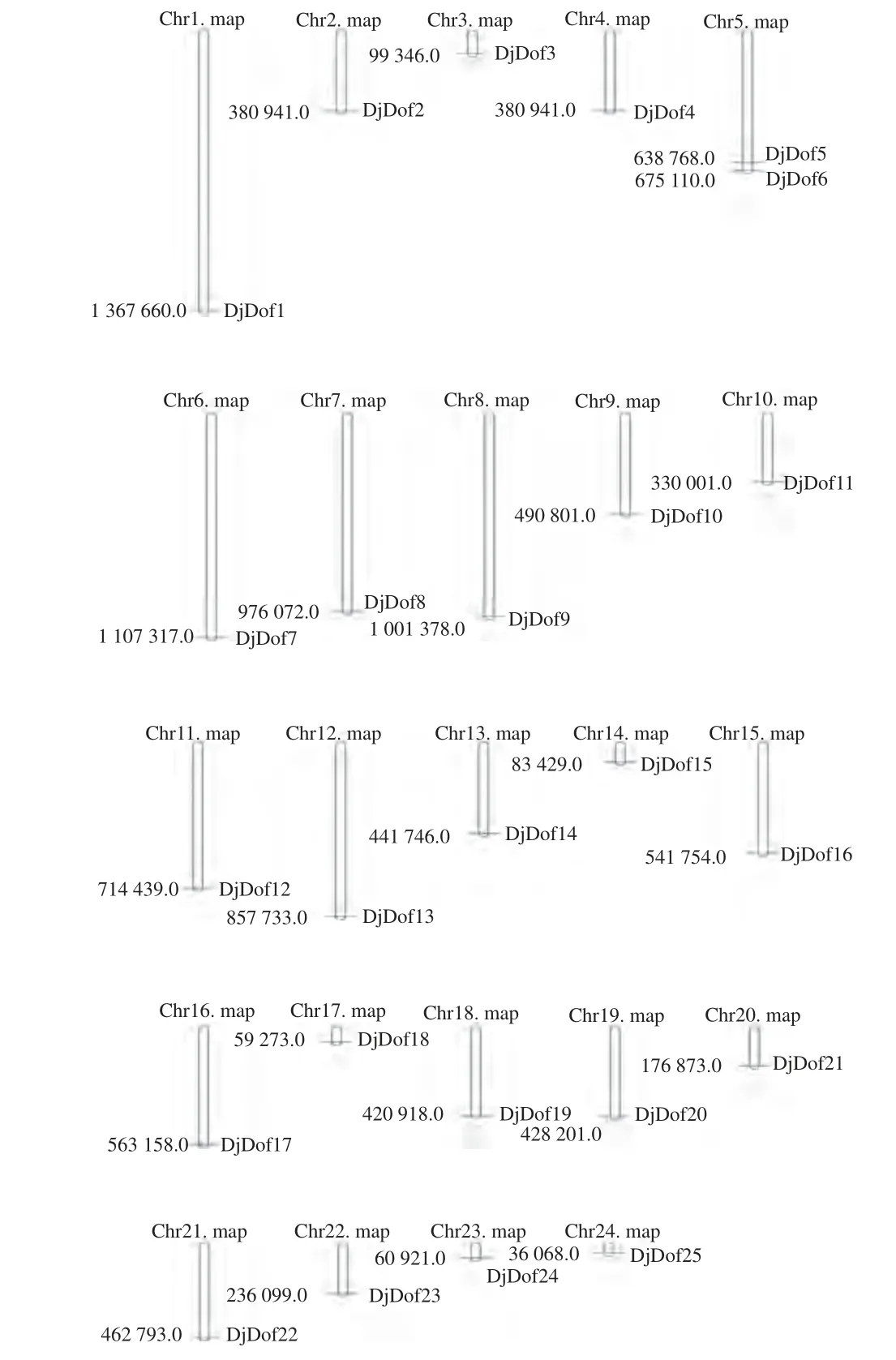
图9 黄藤Dof基因在染色体上的定位Fig. 9 The chromosome localization of Dof gene in D. jenkinsiana
3 结论与讨论
Dof家族是植物特有的一类转录因子,在植物生长发育调控和逆境胁迫响应中扮演着重要角色[16],本研究中利用黄藤全基因组测序数据信息对Dof家族进行了全面分析,共鉴定出25个Dj-Dof家族成员,其数量与水稻(30个)、番茄(34个)、黄瓜(Cucumis sativus)(36个)以及拟南芥(36个)相近,通过统计分析发现Dof家族成员数量与基因组大小并不呈正比例关系。黄藤Dof家族成员内含子数在1~2之间,基因结构相对简单,表明Dof家族的基因结构具有保守性。基因复制事件是植物在进化过程中形成结构相似、功能相仿基因的基础。发现白菜Dof家族基因发生了10次染色体片段重复事件,无串联重复事件发生[16]。发现黄瓜Dof家族基因发生了6次染色体片段重复事件和2次串联重复事件。此外,在大豆、蒺藜(Tribulus terrestris)苜蓿(Modicago sativa)和番茄Dof家族基因中染色体片段重复事件分别发生37、10和8次;串联重复事件分别发生1、1和3次[31-33]。本研究中鉴定到的25个DjDof基因共发生1次串联重复事件,无染色体片段重复事件发生,这表明基因串联重复在黄藤Dof家族扩增和进化中起着重要作用。
本研究结果表明,黄藤Dof家族成员在进化中存在着一定的保守性,主要表现为:1)黄藤Dof蛋白性质与结构的稳定:鉴定得到的25个蛋白不仅在氨基酸数目上较为接近,在分子量上也均为30 kDa左右。既含有酸性蛋白又含有碱性蛋白,正电荷残基数多于负电荷残基数,并均为不稳定蛋白和亲水性蛋白。此外,在二级结构组成比例上,α−螺旋和无规则卷曲为主导。2)基因结构的保守:尽管序列长度上差异较大,但黄藤Dof家族基因在结构上较为相似,均含有1~2个内含子。本研究结果也表明,亚细胞定位显示所有的Dof蛋白都定位在细胞核上,说明Dof蛋白在植物的生长发育中发挥调控作用。
根据进化关系将拟南芥Dof基因家族划分为9个进化亚群[34],本研究中黄藤Dof基因家族成员同样归为9个进化亚群(ClassⅠ.1,ClassⅠ.2,ClassⅡ.1,ClassⅡ.2,ClassⅡ.3,ClassⅢ.1,ClassⅢ.2,ClassⅣ.1,ClassⅣ.2)。结合黄藤Dof基因家族蛋白的保守基序发现,同一亚群中的Dof蛋白所包含的保守基序类型、数量、排列顺序较为类似;正如已经报道的拟南芥、水稻、胡椒和菊花相同组的Dof蛋白的保守基序一样[35-37],黄藤同一组的Dof蛋白保守基序的氨基酸序列也是高度保守的(图3),该结果进一步支持了系统发育树中黄藤Dof蛋白的分组。本研究还发现motif1存在于25个黄藤Dof蛋白中,这与拟南芥、黄瓜和番茄的研究结果[38-39]相吻合。亲缘关系的远近用于判断2种植物在结构、功能上的异同点,黄藤与拟南芥的进化关系表明两者亲缘关系不是很近。此外,还构建了黄藤与拟南芥等10种植物的Dof基因家族系统进化树,结果显示,黄藤与甘蓝型油菜和小麦有较近的亲缘关系,序列同源性也较高,与可可亲缘关系最远。
本研究利用生物信息手段对黄藤Dof基因家族编码蛋白的理化性质、跨膜结构、蛋白二三级结构建模、分子进化、基因结构、染色体定位等进行了详细的分析,为了解其调控机制以及通过基因和蛋白来育种,增加其经济价值打下了一定的基础,同时也填补了关于黄藤Dof基因家族生物信息学方面的空白,对深入研究其生物学功能和调控机制具有很重要的意义。
