不同地形100年生杉木人工林土壤理化性质及林下植被多样性差异分析
2021-12-11陈义堂杨秋菊何宗明曹光球陈爱玲
张 筱 陈义堂 杨秋菊 何宗明 曹光球 陈爱玲,
(1. 福建农林大学资源与环境学院,福建 福州 350002;2. 福建省洋口国有林场,福建 南平 353211;3. 广东通略规划设计有限公司,广东 广州 510000;4. 国家林业草原杉木工程技术研究中心,福建 福州 350002)
杉木是我国南方最重要的用材树种之一,其栽培面积及蓄积量皆居全国人工造林树种首位[1-2]。长期以来的林业生产实践表明,杉木人工林高密度纯林种植不利于林下植被恢复,造成林分稳定性差,从而导致地力下降。除此之外,为提高杉木木材产出,杉木人工林普遍采用短轮伐期经营策略,这种经营策略在一定程度上加速了土壤养分消耗,也是导致杉木人工林地力下降的重要原因[3]。为提高杉木人工林物种多样性及缓解杉木人工林地力下降速度,延长杉木轮伐期是一种行之有效的途径之一。诸多研究结果表明,延长杉木轮伐期,不仅可以改善杉木人工林土壤的理化性质,提高杉木人工林的生物多样性,而且还可提高杉木人工林枯枝落叶的分解速度,增强林分自肥的能力,从而达到维持杉木人工林地力的目的[4-8]。福建省南平市王台镇溪后村安曹下现存1919年营建的杉木人工林,该林分是我国目前林龄最大、单产最高的杉木人工林,总面积285 hm2。由于该林分经营历史悠久,人为干扰低,长期以来,诸多学者对该片林分的生长量[9]、生物量[10]、土壤理化性质演变[11-14]、林下植被演替[14-16]及土壤微生物变化[17-18]等内容进行了长期的跟踪研究,具有较高的研究价值。据文献表明[9],该林分距最近一次调查的时间为2002年,此后相当长的一段时间内,该林分的土壤理化性质及林下植被发育的演变情况尚未有人开展。
地形是影响森林生态系统土壤呼吸的重要因子,可通过重新调控土壤水分、光照及温度等条件来影响土壤呼吸,同时制约着土壤理化性质和植被类型分布,而土壤养分作为植被生长的主要载体,其养分状况可因地形的改变而发生变化,从而影响森林植被群落构成及其多样性的空间布局差异。目前许多专家对森林土壤养分的空间分布规律研究较多[19-22],但对森林土壤养分空间分布的影响因素研究报道较少。地形作为影响土壤养分的主要因素,分析两者之间的关系,可为林木生长和土壤质量的提高提供理论基础。不同植被在其生长过程中会受土壤养分转变的影响[23],从而造成植物群落物种多样性改变,并且在植被对土壤养分吸收利用时又会影响土壤的稳定,所以针对不同地形土壤理化特性与植被生物多样性之间的联系研究需要加强,尤其是山坡地与山洼地理化分布差异对林下植被物种群落演变造成影响还需开展深入研究。鉴于此,本研究以福建省南平市王台镇溪后村安曹下100年生杉木人工林为研究林分,依据坡地、凹地及洼地等不同地形分别设立标准样地,在林分生长调查的基础上,通过分析不同地形老龄杉木林土壤理化性质及林下植被多样性,并进行相关性分析。研究结果为杉木人工林林下植被恢复与保护以及长期生产力维护提供理论参考依据。
1 研究区概况
研究林分位于福建省南平市王台镇溪后村安曹下(117°57′E,26°28′N),该地属武夷山系南伸支脉,平均海拔200 m左右,为中亚热带季风气候,年均降水量与蒸发量分别为1 699 mm和1 413 mm,年均气温19.3 ℃,相对湿度83%。土壤为花岗岩发育而成的红壤,平均土层厚度在100 cm以上。
该林分造林地前身为天然阔叶林,于1918年10月炼山整地,并选取健壮2年生杉木萌芽条顶枝在次年3月进行插条造林,初植密度为3 600 株/hm2,造林前3年定期进行劈草等幼林抚育措施。第8年与第11年间伐部分生长不良木及病木,此后采用封育的管理措施,林分现存平均密度为848株/hm2。灌木层主要植被有紫麻(Oreocnide frutescens)、黄绒润楠(Machilus grijsii)等,草本层主要植被有赤车(Pellionia radicans)和三叶崖爬藤(Tetrastigma hemsleyanum)等。
2 研究方法
2.1 样地设置及调查
2019年4月份依据该林分地形特点将该林地分为西南坡地、西北坡地、山凹地和山洼地。分别在西南坡地、西北坡地和山凹地分别设置3块20 m×20 m样地,山洼地因面积限制,共设置3块10 m×15 m的样地;对每个样地内杉木进行每木调查,林分生长情况见表1。在各样地四周和中心位置各设立5个5 m×5 m样方调查灌木层的植被种类、名称、胸径、树高等指标,随机设置10个1 m×1 m小样方调查草本层植物的种类、株数、地径、高度、盖度等。在每个样地内按对角线挖掘5个60 cm深的土壤剖面,先用环刀分别取0~20、20~40、40~60 cm土壤样品测定土壤物理性质,再取0~20、20~40、40~60 cm土壤样品测定化学性质。同一样地同一土层土样充分混合后作为一个土样带回实验室,风干磨碎过筛后进行化学性质测定。
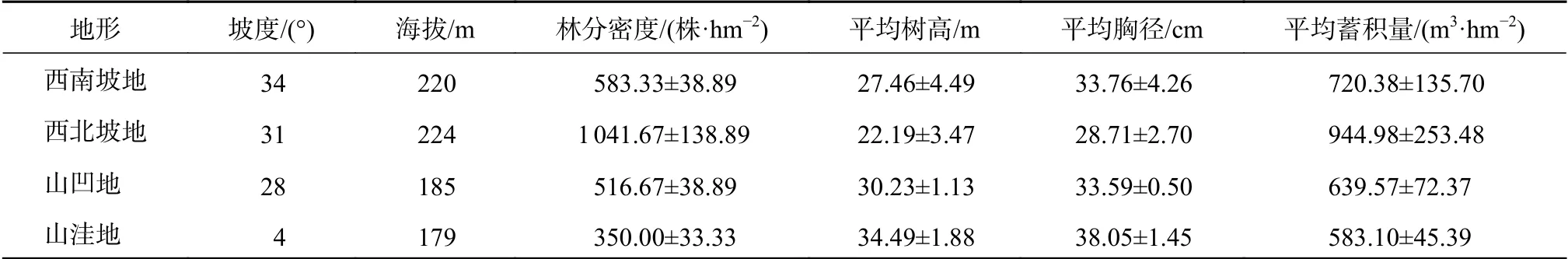
表1 林分生长情况Table 1 Basic conditions of the stand
2.2 土壤理化性质测定
用环刀法测定土壤容重、土壤含水率、总孔隙度、毛管孔隙度和非毛管孔隙度等物理性质。土壤有机质含量采用重铬酸钾容量法测定,全氮含量采用硒粉-硫酸钾-硫酸消化蒸馏滴定法测定,全磷含量采用硫酸-高氯酸消煮-钼锑抗分光光度计法测定,全钾含量采用高氯酸-氢氟酸消煮-火焰光度法测定,速效磷含量采用盐酸和硫酸溶液浸提法测定,速效钾含量采用乙酸铵浸提-火焰光度法测定[24]。
2.3 植被多样性分析
通过野外调查数据,利用样方内灌草植被的株数、频度和盖度分别计算相对应的相对密度、相对频度和相对优势度(相对盖度)以及相对重要值,并通过物种的相对重要值分别计算物种多样性指数[25],包括物种丰富度指数(S)、Shannon-Wiener指数(H)、Simpson优势度指数(D)、Pielou均匀度指数(J)。
2.4 数据处理与分析
使用Microsoft Excel 2019对数据进行描述性分析和处理,通过SPSS 21.0软件进行单因素方差分析(ANOVA),使用Duncan's法进行多重比较,使用双变量相关分析计算土壤养分含量分别与林下植被物种多样性之间的Person相关系数。使用GraphPad Prsim 8绘制柱状图和热图。
3 结果与分析
3.1 不同地形杉木老龄林土壤物理性质分析
3.1.1 土壤容重
由表2可知,4种地形下的土壤容重均随土层加深呈逐渐上升的趋势,且仅在山洼地不同土层中出现显著差异,而同一深度土层中,4种地形下土壤容重均表现为山洼地最高,变化范围为1.19~1.57 g/cm3,其余3种地形数值相差不大。其中土层0~20 cm中,山洼地的土壤容重较西南西北坡地和山凹地分别增高0.79%、3.15%和3.94%,且4种地形差异不显著;土层20~40 cm中,山洼地的土壤容重较西南、西北坡地和山凹地分别增高10.96%、9.59%和8.22%,且西南坡地和山洼地与其他地形均有显著差异;土层40~60 cm中,山洼地的土壤容重较西南、西北坡地和山凹地分别增高9.87%、10.53%和6.58%,且西北坡地和山洼地与其他地形均有显著差异。
3.1.2 土壤含水率
由表2可知,4种地形下的土壤含水率均随土层加深呈逐渐下降的趋势,且仅在西北坡地不同土层中出现显著差异。而同一深度土层中,4种地形下土壤含水率均表现为山洼地最高,变化范围为18.18%~36.17%;西北坡地和山凹地次之;西南坡地最低;相较其他地形,山洼地含水率较高可能是由于该地处于水湿条件较好的环境。其中土层0~20 cm中,山洼地的土壤含水率较西南、西北坡地和山凹地分别增高41.82%、9.76%和25.47%,且4种地形差异显著,但土层20~40 cm和40~60 cm中,4种地形含水率表现为差异不显著。
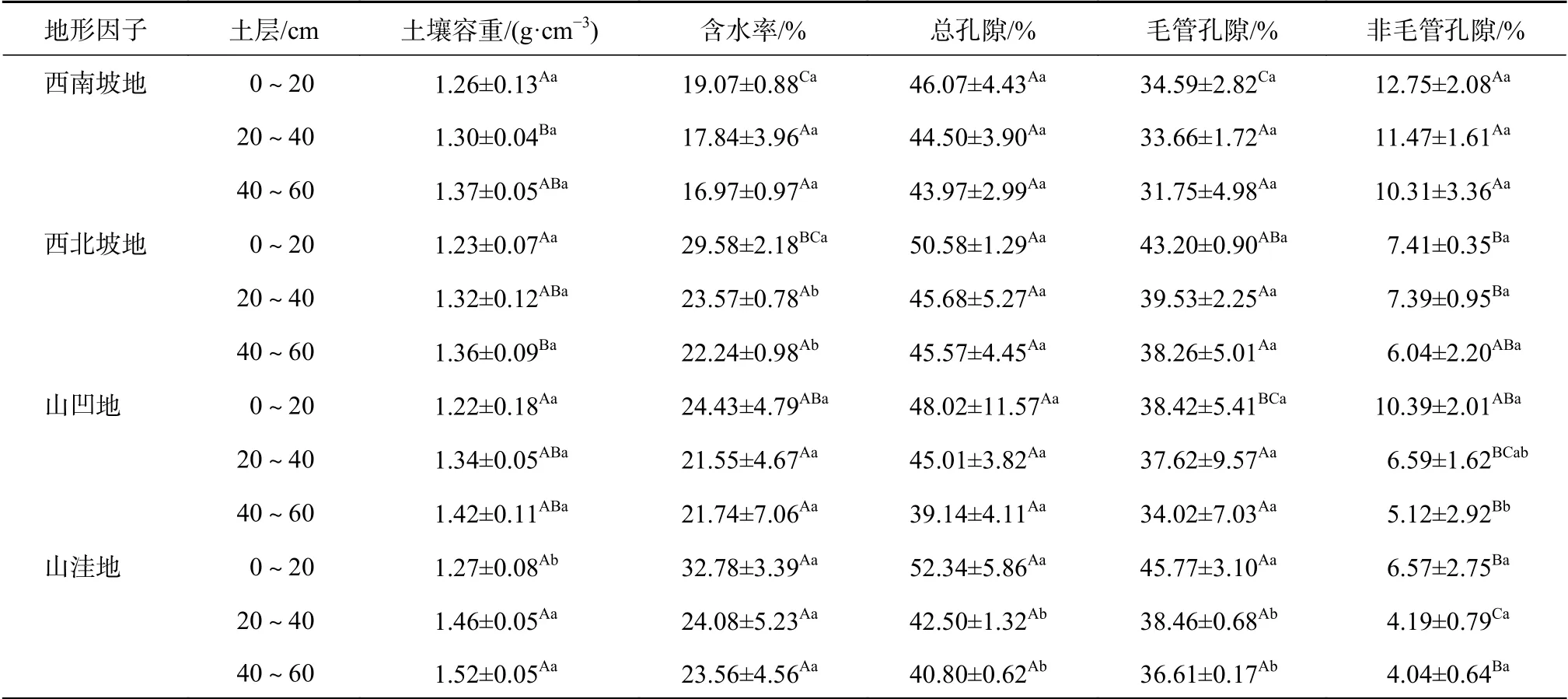
表2 不同地形老龄杉木人工林土壤物理性质Table 2 Soil physical property of aged Chinese fir plantations with different terrains
3.1.3 土壤孔隙度
由表2可知,4种地形下的土壤孔隙度均随土层加深呈逐渐下降的趋势,且仅在山凹地和山洼地不同土层中出现显著差异。而同一深度土层中,4种地形下的土壤总孔隙度与毛管孔隙度均表现为山洼地最高,变化范围分别为40.18%~58.2%和36.44%~48.87%;其余3种地形数值相差不大,但非毛管孔隙度与其相反,表现为西南坡地最高,变化范围为6.34%~14.79%。其中土层0~20 cm中,4种地形下的土壤总孔隙度无显著差异,而毛管孔隙度和非毛管孔隙度均有显著差异,山洼地的总孔隙度较西南、西北坡地和山凹地分别增高12.04%、3.36%和8.25%,毛管孔隙度分别增高24.43%、5.62%和16.06%,非毛管孔隙度分别下降48.47%、11.34%和36.77%;但在土层20~40 cm和40~60 cm中,4种地形下的土壤总孔隙度和毛管孔隙度均无显著差异,仅有非毛管孔隙度出现显著差异。
3.2 不同地形杉木老龄林土壤化学性质分析
3.2.1 土壤有机质含量
由表3可知,4种地形下的土壤有机质含量均随土层加深呈逐渐下降的趋势,且仅在除山凹地外其余3种地形不同土层中有显著差异,而同一深度土层中,4种地形下土壤有机质均表现为西南坡地最高,西北坡地和山凹地次之,山洼地最低。其中0~20 cm土层中,山洼地的土壤有机质含量较西南、西北坡地和山凹地分别下降29.33%、25.02%和7.1%,且4种地形差异不显著;土层20~40 cm中,山洼地的土壤有机质含量较西南、西北坡地和山凹地分别下降34.46%、27.22%和29.17%,且4种地形差异不显著;土层40~60 cm中,山洼地的土壤有机质含量较西南、西北坡地和山凹地分别下降57.77%、37.17%和49.22%,且西北坡地和山洼地与其他地形均有显著差异。

表3 不同地形老龄杉木人工林土壤化学性质Table 3 Soil chemical property of aged Chinese fir plantations with different terrains
3.2.2 土壤氮、磷、钾含量
由表3可知,4种地形下的土壤氮、磷、钾含量大多随土层加深呈逐渐下降的趋势,且土壤全氮仅在除山凹地外其余3种地形不同土层中有显著差异,全钾仅在西北坡地有显著差异,4种地形下全磷等养分含量均有显著差异,而同一深度土层中,4种地形下的全氮和速效钾含量均表现为西南坡地最高,变化范围分别为045~1.46 g/kg和83.5~164.38 mg/kg;全磷和全钾含量均表现为山洼地最高,变化范围分别为225.45~239.72 g/kg和58.11~66.18 g/kg;速效磷含量表现为山凹地最高,变化范围为1.04~2.75 mg/kg。其中土层0~20 cm中,西南坡地的全氮和速效钾含量较西北坡地和山凹山洼地分别增高22.66%、32.81%、30.47%和5.75%、23.93%、29.33%,且4种地形下的速效钾仅在西南坡地中与其他地形均有显著差异而全氮没有,但在土层20~40 cm和40~60 cm中,全氮表现为在西南坡地和山洼地中与其他地形均有差异显著且速效钾均有显著差异;土层0~20 cm中,山洼地的全磷和全钾含量较西南、西北坡地和山凹地分别增高21.83%、40.98%、32.08%和12.94%、21.45%、4.4%,且4种地形下的全磷仅在西南、西北坡地中与其他地形均有显著差异而全钾没有,但在土层20~40 cm中,全钾表现为在西北坡地和山洼地中与其他地形均有显著差异而全磷表现为在西北坡地和山凹地中与其他地形均有显著差异,土层40~60 cm中,全磷和全钾均表现为在西北坡地和山凹地中与其他地形均有显著差异;土层0~20 cm中,山凹地的速效磷含量较西南、西北坡地和山洼地分别增高8.94%、7.23%和0.85%,且4种地形差异不显著,但在土层20~40 cm中,其表现为在西北坡地和山洼地中与其他地形均有显著差异,在土层40~60 cm中,其表现为在西北坡地和山凹地中与其他地形均有显著差异。
3.3 不同地形杉木老龄林林下植被物种组成及其多样性分析
本研究调查可知,该杉木老龄林林下植被共计83种植物,其中茜草科(Rubiaceae)、壳斗科(Fagaceae)和蹄盖蕨科(Athyriaceae)的物种数最多,剔除频度过低的物种,并以重要值大于11为优势种判定标准,筛选出重要值排序前10的物种,详情见表4。由表4可知,除木荷(Schima superba)和芒萁(Dicranopteris dichotoma)等优势种仅在1种地形出现外,其余优势种均出现在多种地形。其中灌木层中,重要值最高的优势种为紫麻(Oreocnide frutescens),同时出现在西南坡地和山洼地,但其在两地的重要值相差较大,表现在山洼地最高,数值可达64.77远超过其他地形的不同物种,山凹地的优势种黄绒润楠(Machilus grijsii)、大叶白纸扇(Mussaenda esquirolli)和大叶紫珠(Callicarpae Macrophyllae)分别大于各自在西北坡地、山洼地和西南坡地的重要值。在草本层中,重要值最高的优势种为赤车(Pellionia radicans),同时出现在西南坡地、山凹地和山洼地,其最高的重要值主要出现在山洼山凹地,数值分别可达60.52和46.72,不仅超过西南坡地且远超于其他地形的不同物种,西北坡地的优势种三叶青(Tetrastigma hemsleyanum)重要值为24.86,大于其在西南坡地的重要值,西南坡地的优势种毛柄断肠蕨(Allantodia crinipes)重要值为21,大于其在山凹山洼地的重要值,深绿卷柏(Selaginella doederleinii)虽同时出现在西北坡地和山凹地,但仅在西北坡地作为优势种。

表4 不同地形老龄杉木人工林灌草层植被物种及其重要值Table 4 Species and important values of shrub and herb layer vegetation in aged Chinese fir plantations
由图1可知,不同地形杉木老龄林中,灌木层多样性指数变化相对草本层变化较为明显。其中灌木层物种多样性表现为,山洼地的Simpson指数和Pielou均匀度指数分别为0.45和0.58,均显著低于其他3种地形,西南坡西北坡地的Shannon-Wiener指数分别为1.89、2.27,大于山凹地的1.75,山凹地大于山洼地的0.93,且西南坡、西北坡显著高于山洼地,而4种地形的丰富度指数没有显著性差异。草本层表现为,西南坡地和西北坡地的Pielou均匀度指数显著高于山凹地和山洼地,Simpson指数、Shannon-Wiener指数和丰富度指数在4种地形中均没有显著差异。
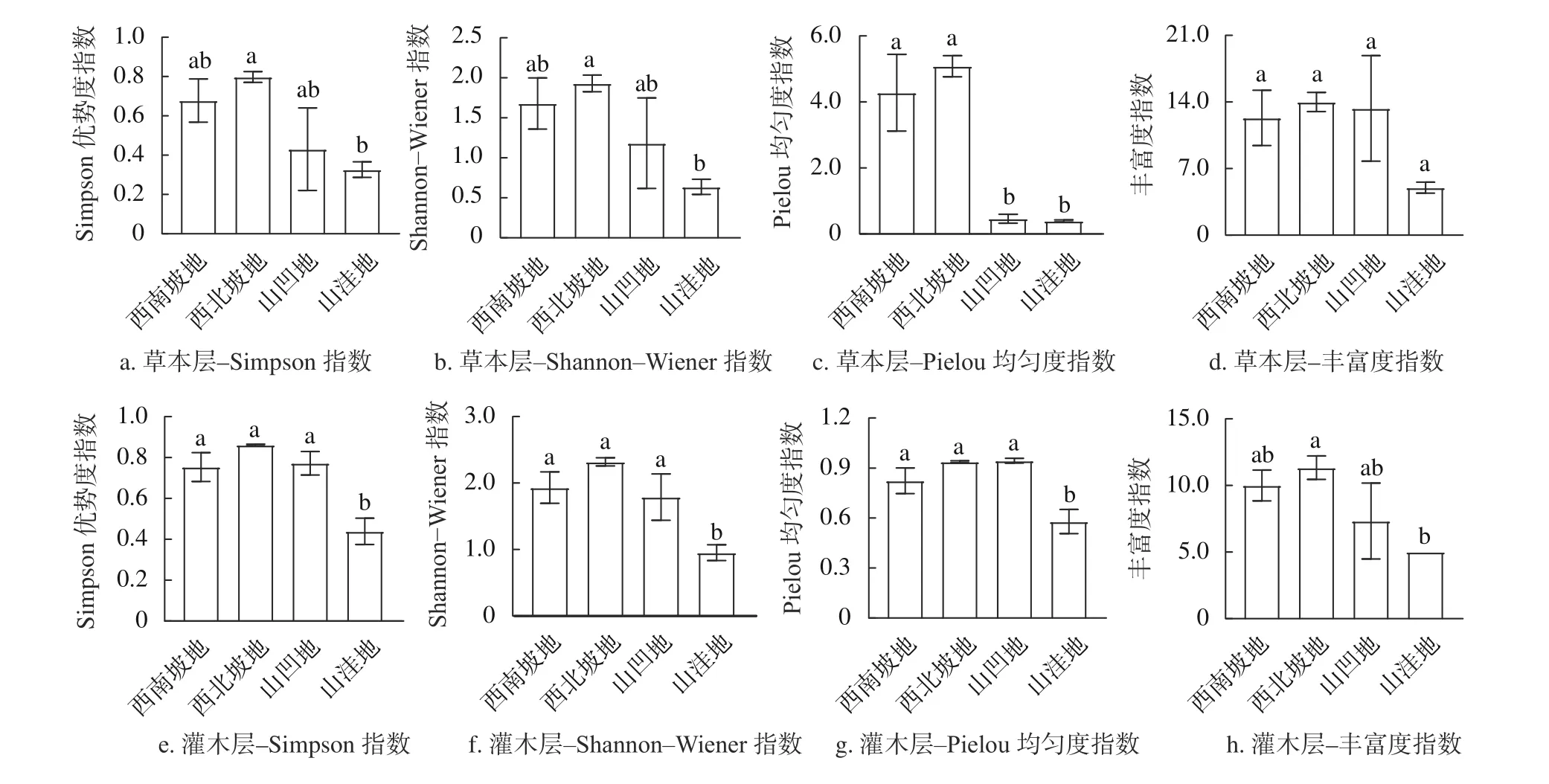
图1 不同地形老龄杉木人工林林下植被物种多样性指数Fig. 1 Species diversity index of undergrowth vegetation of aged Chinese fir plantation in different terrains
3.4 不同地形杉木老龄林林下植被多样性与土壤理化性质相关性分析
由图2可知,植物群落物种多样性指数与土壤理化性质关系密切,灌木层中,土壤含水率与Simpson指数、Shannon-Wiener指数、Pielou均匀度指数呈极显著负相关(P<0.01),毛管孔隙度与4种多样性指数均呈显著负相关(P<0.05),其中丰富度指数与毛管孔隙度呈显著负相关(P<0.05),其他3种指数均与毛管孔隙度呈极显著负相关(P<0.01),土壤全磷含量与4种多样性指数呈显著正相关(P<0.05)。草本层中,Shannon-Wiener指数与土壤含水率、毛管孔隙度呈显著负相关(P<0.05),Pielou均匀度指数与土壤全磷含量呈极显著正相关(P<0.01),与土壤速效钾含量呈显著正相关(P<0.05),丰富度指数与土壤含水率呈显著负相关(P<0.05),与毛管孔隙度呈极显著负相关(P<0.01)。
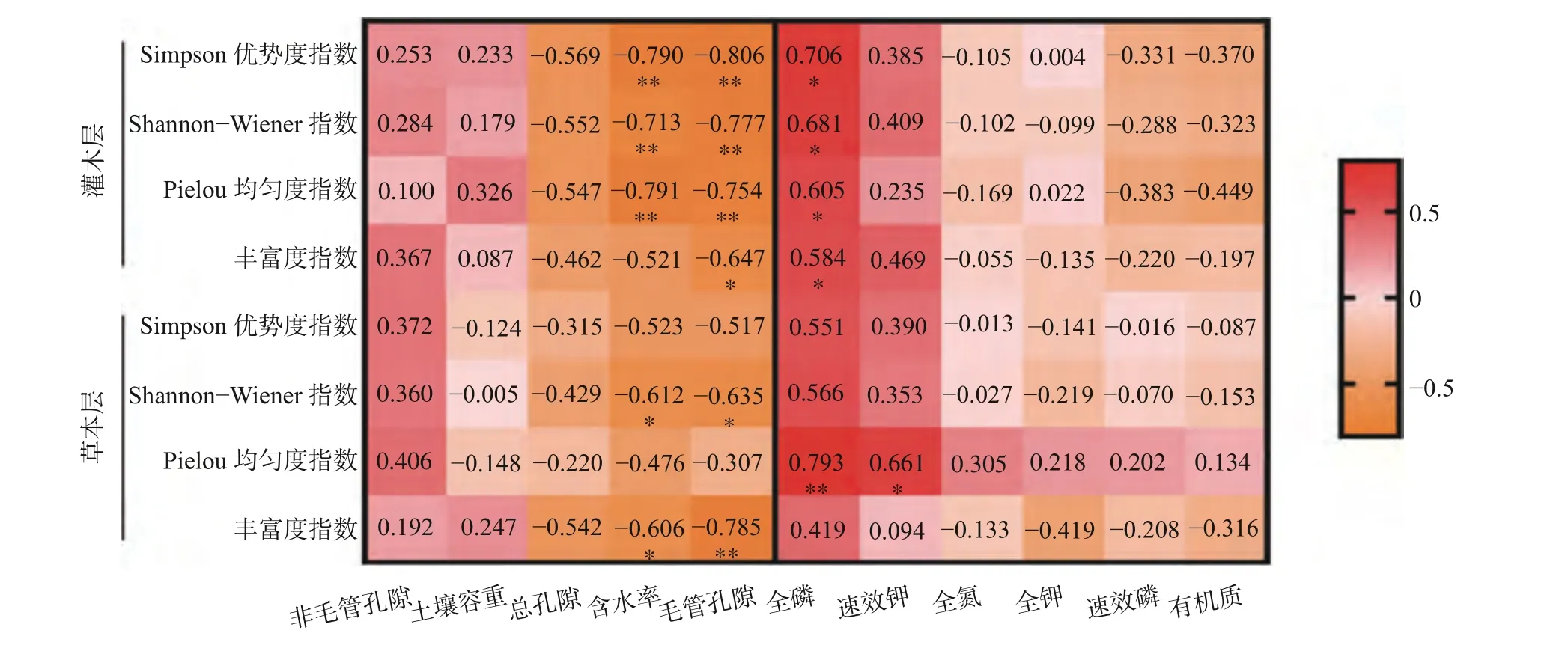
图2 老龄杉木人工林林下植被物种多样性与土壤理化性质的相关性Fig. 2 Correlation between species diversity and soil physicochemical properties of undergrowth vegetation in aged Chinese fir plantation
4 结论与讨论
4.1 讨论
4.1.1 不同地形杉木老龄林土壤理化性质的差异分析
土壤是植物生长的必要介质,是植物养分供给的重要场所,而地形作为土壤性质的主要影响因子,在很大程度上影响着整个林分的土壤理化性质。Griffiths等[26]和Song等[27]的研究发现地形所处的位置和环境决定了光照强度、土壤温度和湿度,进一步影响土壤整个生化过程。不同地形土壤理化性质受控于水和光温的再分配影响,从而造成不同地形条件下土壤水分和养分状况的差异[28-32]。除此之外,随着林分林龄的增加,林下植被组成及林分环境的水、气、光、热等条件随之发生改变,从而对土壤的理化性质变化产生影响[33]。Geroy等[34]的研究发现北坡的太阳辐射量较少,蒸散蒸发速率较低,有助于减少土壤水分损失,此外,北坡土壤含水量较高有利于植物生长和生长季节有机质积累,为土壤生化过程提供了更多的碳基质。土壤水分的保持与地形、光照和气温密切相关,结合实地勘测,相比于西北坡地与山洼地,西南坡地光热条件较好,但其土壤水分含量也因此相对较低,这与上述研究结果相同。此外,土壤的物理性状,受林木和林下植被双重影响,林分密度越大,根系增多会降低土壤的紧密程度,增大土壤中的孔隙度和含水量[35]。本研究中西北坡地的林分密度显著高于其他3种地形,且该地的含水量相对较高,说明林分密度也是含水率的影响因素。
土壤有机质是土壤可持续肥力的主控因子,不仅为植物提供养分来源,为土壤微生物活动提供动能,同时还会对土壤中植物必需的3种大量营养元素氮磷钾产生作用。本研究结果表明,有机质含量总体表现为西南、西北坡地相对较高,山洼地较低,并发现西南、西北坡地有机质含量高与该地形下林下植被物种组成复杂导致的凋落物常年积累量加大相关[36]。研究区4种地形的土壤是燕山晚期白云母化中细粒花岗岩发育的山地暗红壤,比较4种地形的土壤氮磷钾养分情况,总体表现为西南坡地氮磷钾含量相较其他3种地形氮磷钾含量较为丰富。土壤养分主要来源于林分内凋落物的分解,林分密度影响凋落物的数量,并通过光照、温湿度等环境因素影响凋落物的分解速率,从而把养分归还到土壤中去[36]。另有研究表明土壤磷含量在中等密度(1 050株/hm2)相对较低,可能与样地植被种类有关,本研究发现磷含量在西北坡地(1 041.67株/hm2)表现最低,与上述研究结果一致,且除西北坡地外,其他3种地形的林分密度相对较低且相差不大,但养分分布有所差异,全氮和速效钾含量均表现为西南坡地最高,全磷和全钾含量均表现为山洼地最高,说明林分密度并不是不同地形理化差异的主因,更多是跟地形的光照及植被组成有关。对比杨玉盛等[37]在该林分70年生时的土壤肥力研究,结果发现,研究区100年生人工林经过30年的演替,土壤肥力呈下降趋势,有机质、全氮、全磷、速效磷和速效钾含量分别下降了22.62%、14.81%、53.19%、49.81%和19.39%,其中N∶P值也表现为100年生大于70年生,而N∶P值作为凋落物分解速率和养分归还速率的主要影响因子,这说明凋落物分解速率会随林龄增大而降低,利于养分存储,该研究区为针叶林树种的杉木纯林,养分归还比例小,地力维护差,经过30年的演替,老龄杉木林已呈衰退趋势,林冠稀疏,因而林下植被繁多,总生物量变大,同时,林龄过大该林区自身养分利用率下降,且土壤容重、含水率、孔隙度和养分含量均处于较低水平,这说明该林分土壤肥力下降与地形有关外还可能与林分林龄较大相关。
4.1.2 不同地形杉木老龄林林下植被多样性的差异分析
生物多样性作为人工林的研究重点一直是林业领域的主要关注话题,就杉木人工林而言,其研究重心主要是林下植被物种结构和物种多样性[38]。林下植被不仅是杉木人工林反映群落物种多样性的重要指标,还是人工林碳储量的主要来源,其与林分生态环境的稳定息息相关,影响着土壤理化性质、林下光照强度和林下植被生长空间[39-40]。
地形可通过控制不同生态因子的组合和空间分布来影响植被的群落结构[41]。因此,受地形条件影响,人工林林下植被物种组成及其各种功能指数都受到显著影响,且不同地形下的林下植被物种构成和多样性指数变化趋势不同,导致不同地形土壤养分改变。有研究表明[42-43],林内光照变强,林木生长空间扩大,个体间对土壤肥力和地下空间的争夺能力减弱,促进林内植被发育,从而对灌木层有显著影响,促使灌木层植被种类增加,优势种发生改变。何宗明等[44]表明山坡地林下植被生物多样性指数明显高于山洼地。本研究结果表明,光照充足的西南坡地,其林下植被物种多样性显著高于山洼地,其中山洼地赤车和紫麻的重要值分别占草本层和灌木层的60%以上,优势种突出,从Shannon-Wiener指数、均匀度指数和丰富度指数来看,山洼地的指数相对较低,说明该地林下植被稀疏,物种单一,这与上述研究结果相同。王梅等[45]表明不同地形林下植被多样性指数差异可能是由于坡位因素引起的土壤水分和光照等多种生态环境因素差异综合的结果,该研究还发现植被群落总体多样性表现为阴坡稍大于阳坡,这与本研究结果有相似之处,本研究中西北坡的植被物种多样性高于西南坡,说明西北坡地(阴坡)的水热条件适合更多物种生存,物种多样性丰富。
4.1.3 不同地形杉木老龄林土壤理化性质与林下植被多样性的关系
植被生长与其所处的环境密不可分,植被与土壤二者相互制约,其相互间协同变化的作用机理不仅维持着生态系统的动态稳定,并形成植被-土壤系统为生态环境的改善发挥着极其重要的作用。土壤水肥条件优越的地形往往也对应着较高的灌草层物种多样性。本研究相关性分析发现土壤理化性质与林下植被物种多样性关系密切,且存在显著或极显著相关,因此调节土壤含水量、改善土壤通气性、补充土壤营养成分可促进林下植被生长发育。杉木林下物种多样性主要受土壤全磷和速效钾的影响,且灌草层植被在生长过程中对全磷和速效钾的吸收有一定差异,这与植被自身结构及生长发育模式不同有关,不同植被对养分的吸收利用能力有差异,部分对养分吸收作用强的植被抑制其他植被的吸收。Fisk等[46]表明植被物种组成的变化可能与土壤性质的差异有关,这与本研究得出灌草层中物种丰富度和优势度指数均受土壤含水率、孔隙度和全磷的影响较大的结果一致。不同地形土壤养分含量分布差异、环境中水肥和光照的作用及林分密度大小都是影响植被物种多样性的关键因子,这也与相关研究[32]结果一致。
4.2 结论
林下植被的存在,对土壤养分的消耗与流失起到缓冲与补偿作用,可有效缓解土壤肥力的下降速率。Trettin等[47]、王媚臻等[48]认为植物群落林下多样性越丰富,土壤结构相应越好、稳定性越高。本研究发现受地形条件影响,林下植被多样性丰富的山坡地土壤养分状况较好,并得出如下结论:
1)本研究区100年生杉木人工林土壤养分虽然呈下降趋势,但仍具有相对较高的土壤肥力,这也是研究区丰产林持续增长的原因。
2)土壤理化性质和林下植被物种多样性受地形条件约束在不同地形呈现一定差异,西北西南坡地植被物种多样性显著高于山凹山洼地,且山坡地的土壤养分含量也相对更高,更适于植被健康生长。
3)林下植被物种多样性受土壤理化性质影响较大,其中全磷和速效钾是主要影响因素。
4)结合本实验结果,可考虑通过人为方式对杉木林进行水肥调控并加大施肥力度,以此来均衡不同地形的养分含量并适量补植不同地形所缺失的植被,从而稳定不同地形林下植被多样性。
