16份怀地黄种质资源抗旱性评价
2021-12-09田梦平谢小龙杜权马梦雨吴廷娟冯卫生
田梦平 谢小龙 杜权 马梦雨 吴廷娟 冯卫生

摘要:探究怀地黄不同种质资源的抗旱性,为栽培区域提供抗旱品种和培育高抗旱性地黄新品种提供依据。以16份怀地黄主要种质资源为材料,采用盆栽控水法进行干旱胁迫,试验包括正常供水(CK)和6 d不浇水2个处理。测定参试怀地黄种质资源的5个抗旱生理指标,采用主成分分析、隶属函数法、相关分析以及聚类分析对其抗旱性进行综合评价。结果表明,在干旱胁迫下,16份怀地黄种质资源叶片的SOD活性、POD活性、可溶性蛋白含量、Pro含量、相对电导率与对照组相比均有不同程度的提高,5个生理指标的增幅范围分别为4.82%~119.78%、13.58%~102.94%、1.12%~75.81%、2.90%~78.88%、3.45%~123.68%。16份怀地黄种质资源中,抗旱性由强到弱依次为北京1号>狮子头>金线钓鱼>怀丰>9302>吨王>83抚育>沁怀1号>郭里茂>85-5>新科>红薯王>金九>北京3号>金状元>三块。相关分析表明,抗旱指数与SOD活性相对变化率呈显著正相关;与可溶性蛋白含量相对变化率呈极显著正相关。聚类分析表明,16份怀地黄种质资源被划分4个类群,其中类群Ⅰ中的北京1号和狮子头2份种质资源为强抗旱资源;类群Ⅱ中的金线钓鱼为较抗旱资源;类群Ⅲ中的三块为弱抗旱资源;类群Ⅳ为中等抗旱资源,数量最多,占总数75.00%。综上,供试的16份怀地黄种质资源在抗旱性上有显著差异,可为地黄抗旱品种的培育提供亲本材料。
关键词:怀地黄;种质资源;抗旱性;生理指标;隶属函数;主成分分析;相关分析
中图分类号:S567.23+9.01 文献标志码: A
文章编号:1002-1302(2021)21-0153-06
收稿日期:2021-05-10
基金项目:国家重点研发计划(编号:2017YFC1702800);中央引导地方科技发展专项(编号:171100310500)。
作者简介:田梦平(1995—),女,河南商城人,硕士研究生,研究方向为中药资源。E-mail:tmpmengping@163.com。
通信作者:谢小龙,博士,副教授,主要从事中药资源与栽培研究。E-mail:xiaolongxie@126.com。
地黄源于玄参科植物地黄(Rehmannia glutinosa Libosch.)的新鲜或干燥块根,以栽培品入药,以旧称“怀庆府”的河南省焦作地区(温县、沁阳、武陟、博爱、孟县、修武等地)为道地产区,其所产地黄习称为“怀地黄”[1-3]。在地黄栽培过程中,水分调控至关重要,怀地黄产区农民总结出素有“三浇三不浇”的经验,以合理地对地黄生产田的水分进行调控[4]。地黄根系少,吸水能力差,过分的干燥、潮湿的气候和不良的排水环境,都不利于地黄的生长发育,容易引起病害,关键时期水分供应不足,会严重影响地黄的产量和品质[5]。因此,明确地黄在干旱胁迫下的生理变化情况,筛选抗旱性强的地黄种质资源,对提高地黄产量和质量有重要意义。
旱害是指土壤水分缺乏或大气湿度相对过低对植物的伤害,干旱会导致叶片和嫩茎萎蔫,气孔开度减小[6]。此外,逆境胁迫还会导致一系列代谢危害,如活性氧大量积累、膜功能紊乱、抑制蛋白合成、脂类含量降低等[7]。当植物遭受胁迫时,植物体内会产生可溶性糖、脯氨酸(Pro)、甜菜碱等物质来调节渗透压,植物抗氧化酶系统如过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)等抗氧化物可以解除胞内活性氧毒害,进而维持植物在逆境胁迫条件下的正常生长需要[8-10]。目前,关于干旱胁迫下地黄生理变化的研究鲜有报道。因此,本研究以16份怀地黄主要种质资源为材料,对干旱胁迫下地黄苗期叶片的生理指标进行分析,采用隶属函数法对各种质资源的抗旱性进行评价,为筛选高抗旱性地黄品种提供参考。
1 材料与方法
1.1 试验材料
本研究所用16份怀地黄种质资源均来自温县地黄种质资源圃,具体名称见表1。
1.2 试验设计
干旱胁迫采用盆栽模拟干旱法, 在河南中医药大学药用植物园进行。塑料盆深8 cm、开口直径13.5 cm、底部直径8.5 cm。培养基质为温县黄河沙壤土和营养土,按3 ∶2的体积比充分混合均匀,每个盆内施入10 g复合肥。2020年7月15日进行种植,每盆播种2块大小一致的种栽,每个品种25盆,至地黄苗期时,进行干旱胁迫试验。干旱胁迫采用盆栽模拟干旱法,试验设置2个处理,对照组(CK)正常供水,干旱处理组进行不浇水处理,胁迫6 d后,取相同叶位叶片进行生理指标测定。
1.3 生理指标测定方法
SOD活性测定采用氮蓝四唑(NBT,nitro-blue tetrazolium) 法[11-12];POD活性测定采用愈创木酚法[11,13];可溶性蛋白含量测定采用考马斯亮蓝 G-250染色法[11,14];Pro含量测定采用酸性茚三酮显色法[11,15];相对电导率测定采用浸泡法[11,16]。
1.4 数据处理
采用Excel 2013进行数据整理,运用SPSS 21.0进行主成分分析、隶属函数分析、相关分析以及聚类分析。为避免参试材料间因基础性状不同对结果造成的影响,采用各指标的相对变化率进行评价[17]。
相对变化率=(测定值-对照测定值)/对照测定值×100%。
相对变化率主成分分析:将单项综合指标的相对变化率进行主成分分析,将原来单个综合指标转化成新的综合指标。
隶属函数计算[18-19]:将每个品种的各综合指标分别用下列公式求出相应的隶属值,公式为U(Xj)=(Xj-Xmin)/(Xmax-Xmin),j=1,2,…,n。式中:Xj表示第j個综合指标;Xmax表示第j个综合指标的最大值;Xmin表示第j个综合指标的最小值。
权重的确定[18,20]:根据综合指标的综合特征值求出各综合指标的权重。Wj=Pj/∑nj=1Pj,j=1,2,…,n。式中:Wj表示第j个综合指标的重要程度;Pj为各资源第j个综合指标的特征值。
综合评价:抗旱指数:D=∑nj=1[U(Xj)×Wj],j=1,2,…,n。
2 结果与分析
2.1 干旱胁迫下怀地黄种质资源叶片生理变化情况
2.1.1 叶片 SOD活性变化
由图1可知,在干旱胁迫下,怀地黄各种质资源的SOD活性与对照组相比,均有不同程度的增加,且均达到显著性水平(P<0.05))。其中,北京1号的增幅最大,高达119.78%;金九、金状元、83抚育、9302、红薯王的增幅较高,范围为31.29%~49.23%;其余9份种质资源的增幅均比较小,低于30%;三块的增幅最小,仅为4.82%。
2.1.2 叶片POD活性变化
由图2可知,在干旱胁迫下,怀地黄各种质资源的POD活性与对照组相比,均有不同程度的增加,除9302外,均达到显著水平(P<0.05)。其中,吨王的增幅最大,达102.94%;其次是狮子头和北京3号,增幅分别为96.14%和62.66%;其余13份种质资源增幅较小,增幅为13.58%~37.94%;其中增幅最小的种质资源是9302。
2.1.3 叶片可溶性蛋白含量变化
由图3可知,在干旱胁迫下,怀地黄各种质资源的可溶性蛋白含量与对照组相比,均有不同程度的增加,且均达到显著性水平(P<0.05)。其中,金线钓鱼的增幅最大,达75.83%;其次为狮子头和9302,增幅分别为53.37%和48.22%;北京1号、怀丰、沁怀1号的增幅均在37%左右;其余10份种质资源增幅较小,在30%以内;增幅最小的种质资源是金九,仅为1.12%。
2.1.4 叶片Pro含量变化
由图4可知,在干旱胁迫下,怀地黄各种质资源Pro含量与对照组相比,均有不同程度的增加,除怀丰和三块2份种质资源外,均达到显著性水平(P<0.05)。其中,85-5的增幅最大,达78.88%;北京1号、新科、红薯王的增幅较大,范围为44.48%~49.62%;吨王、83抚育、狮子头、郭里茂增幅范围为27.84%~35.52%;其余8份种质资源增幅均小于20%,增幅最小的种质资源是金线钓鱼,为2.90%。
2.1.5 叶片相对电导率变化
由图5可知,在干旱胁迫下,怀地黄各种质资源的相对电导率与对照组相比,均有不同程度的增加,除北京1号、怀丰、金线钓鱼3份种质资源外,均达到显著性水平(P<0.05)。其中,北京3号增幅最大,达123.68%;其次是金状元,增幅为109.97%;吨王、85-5、三块、新科的增幅较大,范围为61.13%~81.02%;83抚育、9302、狮子头、红薯王的增幅范围为27.03%~44.78%;其余5份种质资源增幅较小,范围为3.45%~16.76%;增幅最小的种质资源是沁怀1号。
2.2 怀地黄不同种质资源抗旱性综合评价
2.2.1 生理指标相对变化率主成分分析
供试怀地黄种质资源5个抗旱生理指标相对变化率的主成分分析(表2)表明,前3个主成分的贡献率分别为29.692%、24.005%、22.530%,累计贡献率为76.227%。这表明前3个综合指标可以概括5个原有指标的绝大部分信息,可以用这3个综合指标对地黄品种的抗旱性进行概括分析。第1主成分(F1)得分系数最大的指标是可溶性蛋白含量,第2主成分(F2)得分系数最大的指标是Pro含量,第3主成分(F3)得分系数最大的指标是POD活性。它们对应的特征向量表达式分别为:F1=0.153×SOD活性+0.148×POD活性+0.661×可溶性蛋白含量-0.173×Pro含量-0.452×相对电导率,F2=0.597×SOD活性+0.043×POD活性-0.087×可溶性蛋白含量+0.683×Pro含量-0.070×相对电导率,F3=-0.002×SOD活性+0.900×POD活性+0.268×可溶性蛋白含量+0.047×Pro含量+0.240×相对电导率。
2.2.2 抗旱综合指标隶属函数分析 根据各主成分(F1、F2、F3)的特征值求出各综合指标的权重分别为0.442、0.309、0.249,根据综合评价公式求出各怀地黄种质资源抗旱指数D值。16份种质资源抗旱能力强弱排序依次为北京1号>狮子头>金线钓鱼>怀丰>9302>吨王>83抚育>沁怀1号>郭里茂>85-5>新科>红薯王>金九>北京3号>金状元>三块,即北京1号抗旱性最强,三块抗旱性最弱(表3)。
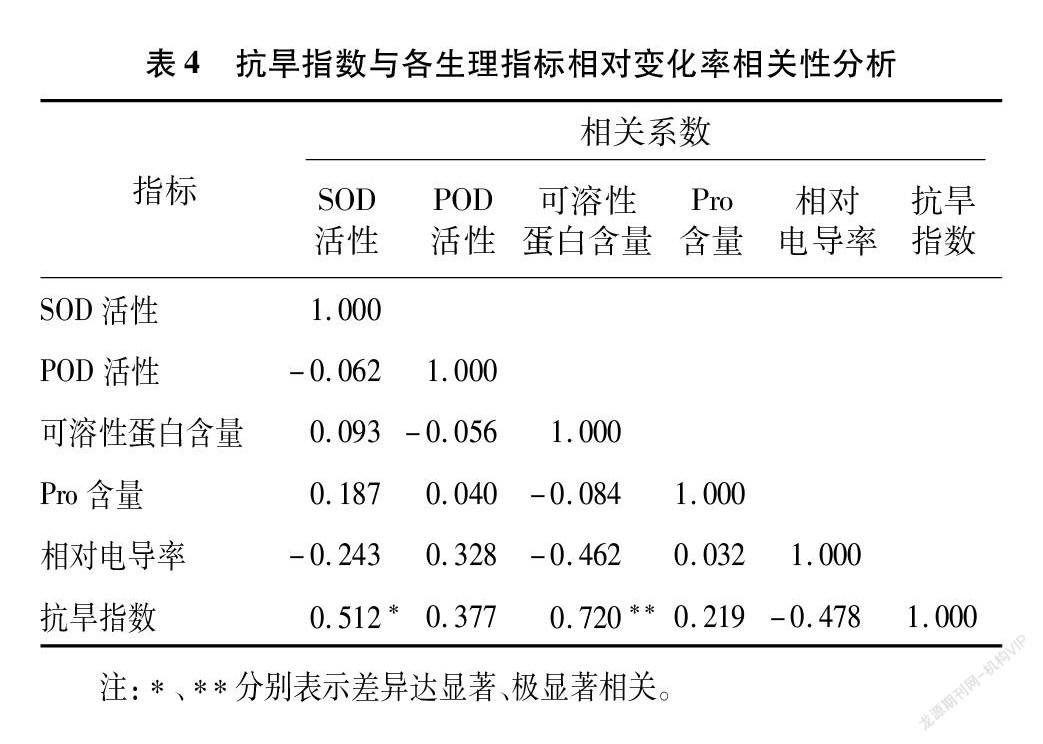
2.2.3 抗旱指数与各生理指标相对变化率相关性分析
抗旱指数与各生理指标相对变化率相关性分析(表4)表明,抗旱指数与SOD活性相对变化率呈显著正相关,与可溶性蛋白含量相对变化率呈极显著正相关,这表明在苗期,地黄主要通过提高SOD活性和可溶性蛋白含量来抵抗干旱胁迫。此外,抗旱指数除与相对电导率相对变化率的相关系数为负值外,其余均为正值,这与植物遭受干旱胁迫时,植物生理指标的变化趋势相符。
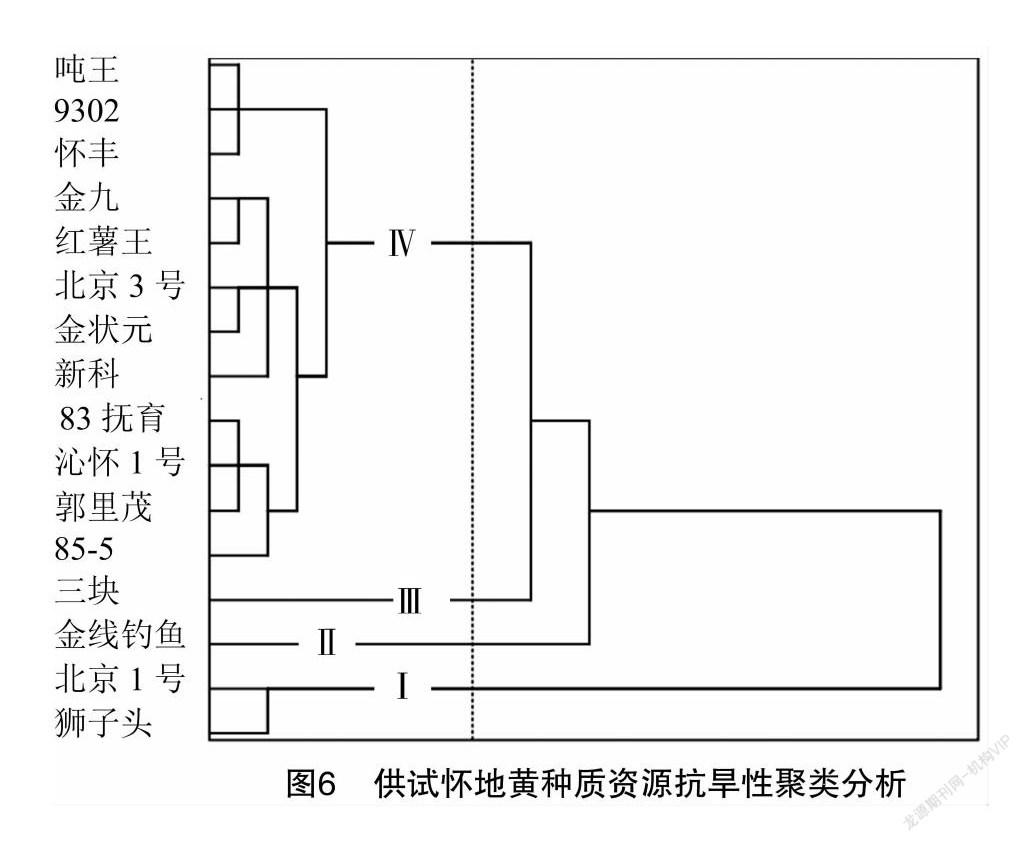
2.2.4 抗旱性聚类分析
基于抗旱指数对供试地黄种质资源作聚类分析,结果(图6)表明,16份怀地黄种质资源被划分为4个类群。类群Ⅰ抗旱指数均值为0.686,是强抗旱资源,包括北京1号和狮子头2份种质资源,占总数12.5%;类群Ⅱ抗旱指数均值为0.518,是较强抗旱资源,包括金线钓鱼1份种质资源,占总数6.25%;类群Ⅲ抗旱指数为0.045,是弱抗旱种质资源,包括三块1份种质资源,占总数6.25%;类群Ⅳ包含种质资源最多,占总数75.00%,抗旱指数均值为0.293,是中等抗旱资源,包括噸王、9302、怀丰、金九、红薯王、北京3号、金状元、新科、83抚育、沁怀1号、郭里茂、85-5等12份种质资源。
综上所述,16份种质资源的抗旱性分为强抗旱、较强抗旱、中等抗旱、弱抗旱4个等级,其中类群Ⅰ的北京1号和狮子头2份强抗旱种质资源和类群Ⅱ的金线钓鱼1份较抗旱种质资源可为抗旱地黄品种的培育提供优质素材。
3 讨论与结论
在植物生长史中,幼苗是最关键的阶段之一。在植物不同生育期中,幼苗期一般对外界环境忍耐力最小,抗性最弱[21-23]。因此,本研究选择在幼苗期对怀地黄种质资源的抗旱性进行评价。
SOD和POD是植物体内重要的抗氧化酶系统,干旱胁迫下,SOD和POD活性会增强,能有效清除细胞内活性氧自由基,提高植物抗氧化能力[24-25]。本研究表明,干旱胁迫下,16份怀地黄种质资源的SOD和POD活性均有不同程度的增加,其中SOD活性增幅最大的种质资源是北京1号,增幅最小的是三块,POD活性增幅最大的种质资源是吨王,增幅最小的是9302。脯氨酸是一种重要的渗透调节物质,植物受到胁迫时,脯氨酸会大量积累,来减缓胁迫造成的植物细胞生理代谢不平衡,进而降低干旱带来的伤害[26-28]。本研究表明,干旱胁迫下,16份怀地黄种质资源的脯氨酸含量均有不同程度的增加,其中85-5的增幅最大,金线钓鱼的增幅最小。干旱时,植物体内蛋白减少,这可能与蛋白质合成酶活性降低有关[29],此外,干旱胁迫还可以诱导一些蛋白质产生和积累,这类蛋白可以保护生物大分子,维持特定细胞结构,改变细胞膜水分通透性,有利于渗透调节[18]。本研究表明,干旱胁迫下,16份怀地黄种质资源的可溶性蛋白含量均有不同程度的增加,这表明,在一定程度上,地黄可以通过增加逆境蛋白含量来提高其抗旱性。当植物细胞膜选择透性遭受逆境破坏时,细胞内电解质外渗,外渗越多,相对电导率越高。本研究表明,干旱胁迫下,16份怀地黄种质资源的相对电导率有不同程度的增加,增幅最大的是北京3号,增幅最小的是沁怀1号,在一定程度上,相对电导率增幅越大,抗性越弱。
植物的抗旱性是由多个生理指标作用的结果,很难由某一个单独指标来界定[30-31]。因此,可以利用隶属函数法对多个生理指标進行综合评价,可靠性高。本试验使用隶属函数法对16份怀地黄种质资源的抗旱性进行综合评价,抗旱性由强到弱依次为北京1号>狮子头>金线钓鱼>怀丰>9302>吨王>83抚育>沁怀1号>郭里茂>85-5>新科>红薯王>金九>北京3号>金状元>三块。
抗旱指数与各生理指标相对变化率相关分析表明,抗旱指数与SOD活性呈显著正相关,与可溶性蛋白含量呈极显著正相关,这表明SOD活性和可溶性蛋白含量可能是评价地黄抗旱性的关键指标。
聚类分析表明,16份怀地黄种质资源共分为4个类群,其中,类群Ⅰ的北京1号和狮子头为强抗旱种质资源,类群Ⅱ的金线钓鱼为较抗旱种质资源。抗旱等级排序和抗旱指数D值大小排序结果一致。
植物在不同生育期受到干旱胁迫后,其抗旱性是否表现的一致,会因植物种类的不同而变化。苗昊翠等研究了2个花生品种不同生育期的抗旱性,结果表明花育25号在4个不同生育期的抗旱性均高于花育33号[32];李显航等研究了7个烟草品种不同生育期的抗旱性,研究表明不同品种在不同生育期抗旱性不同[33];张龙龙等研究发现,6个小麦品种在不同生育时期抗旱性不同[34]。本研究主要是对怀地黄种质资源苗期抗旱性进行比较。 因此,怀地黄种质资源不同生育时期的抗旱性是否一致,不同生育期受到干旱胁迫后对产量和品质的影响程度如何,均需进一步研究。
参考文献:
[1]王太霞,李景原,胡正海. 地黄的形态结构与化学成分研究进展[J]. 中草药,2004,35(5):585-587.
[2]张中朋. 地黄国际市场前景看好——中国地黄及其制品国内外市场简介[J]. 中药研究与信息,2005,7(4):43-44.
[3]冯文龙,解红娥. 种脱毒地黄效益好[J]. 山西农业,2002(5):51.
[4]张莲晓. 地黄的高产栽培技术[J]. 农业开发与装备,2017(5):140.
[5]祁春苗. 氯化胆碱对地黄耐旱性的诱导效应[D]. 郑州:河南农业大学,2007:11.
[6]施海涛. 植物逆境生理学实验指导[M]. 北京:科学出版社,2016:1.
[7]武维华. 植物生理学[M]. 北京:科学出版社,2008:444-457
[8]程 林. 干旱胁迫黄芩生理生态变化及其黄芩苷生物合成的分子生态机制[D]. 长春:吉林农业大学,2018:9.
[9]冯树林. 干旱胁迫和复水对侧柏和紫穗槐幼苗生长和生理特征的影响研究[D]. 杨凌:西北农林科技大学,2020:9-13.
[10]陈 丽. 牧草间作对干旱胁迫下油橄榄根系生长与生理特性的影响研究[D]. 兰州:甘肃农业大学,2020:3-4.
[11]吴强盛. 植物生理学实验指导[M]. 北京:中国农业出版社,2018:63-64,119-123,133-135,138-139.
[12]Durak I,Yurtarslanl Z,Canbolat O,et al. A methodological approach to superoxide dismutase (SOD) activity assay based on inhibition of nitroblue tetrazolium (NBT) reduction [J]. Clinica Chimica Acta,1993,214(1):103-104.
[13]Molaei Rad A,Ghourchian H,Moosavi-Movahedi A A,et al. Spectrophotometric assay for horseradish peroxidase activity based on pyrocatechol-aniline coupling hydrogen donor[J]. Analytical Biochrmistry,2007,362(1):38-43.
[14]Chikakiyo H,Kohama T,Nagata J,et al. Determination of the blood acid-soluble glycoproteins using coomassie Brilliant Blue G-250:its clinical application[J]. The Japanese Journal of Clinical Pathology,1983,31(5):543-548.
[15]Abrahám E,Hourton-Cabassa C,Erdei L,et al. Methods for determination of proline in plants[J]. Plant Stress Tolerance,2015,639:317-331.
[16]Espino S,Schenk H J. Mind the bubbles:achieving stable measurements of maximum hydraulic conductivity through woody plant samples [J]. Journal of Experimental Botany,2011,62(3):1119-1132
[17]李 冰,张敬敬,高秀瑞,等. 低温胁迫下不同基因型西瓜抗寒性综合评价[J]. 中国瓜菜,2019,32(4):16-19.
[18]张 丽,贾志国. 紫薯对干旱胁迫的生理响应及耐旱性评价[J]. 贵州农业科学,2015,43(9):51-55.
[19]朱向涛,金松恒,哀建国,等. 牡丹不同品种耐涝性综合评价[J]. 核农学报,2017,31(3):607-613
[20]张 耿,王 赞,高洪文,等. 21份偃麦草属牧草苗期耐盐性评价[J]. 草业科学,2008,25(1):51-54.
[21]王娅玲,李维峰. 干旱胁迫对植物生长及其生理的影响概述[J]. 南方农业,2015,9(6):37,39.
[22]贺转转,邢佳佳,陈 玲,等. 植物幼苗抗逆机制研究进展[J]. 生物技术通报,2013(2):1-7.
[23]高瑞如,赵瑞华,杨学军,等. 盐分和温度对盐节木幼苗早期生长的影响[J]. 生态学报,2009,29(10):5395-5405.
[24]聂石辉. 大麦抗旱的生理生化机理研究及种质资源抗旱性评价[D]. 石河子:石河子大学,2011:36-37.
[25]李海霞. 六个牡丹品种对干旱胁迫的生理响应机制及抗性评价[J]. 北方园艺,2021(4):64-71.
[26]郑 颖. 6个新引进杜鹃品种的抗旱性评价[D]. 福州:福建农林大学,2018:3-4.
[27]由佳辉,高 林,王海鸥,等. 干旱胁迫对9个葡萄砧木品种生理指标的影响[J]. 经济林研究,2020,38(3):180-189.
[28]王文麗,贾生海,张 芮,等. 不同生育期水分胁迫对设施延后栽培葡萄生理生长及产量的影响[J]. 安徽农业科学,2018,46(20):41-44.
[29]王 鑫,师文俊,李一杰,等. 8个平欧杂种榛品种抗旱性评价[J]. 经济林研究,2020,38(4):161-168.
[30]郭彦宏,张晶星,杨永娟,等. 六种野生广义菊属植物对干旱胁迫的生理响应[J]. 浙江农业学报,2018,30(8):1349-1354.
[31]沈思言,徐艳霞,马春雷,等. 干旱处理对不同品种茶树生理特性影响及抗旱性综合评价[J]. 茶叶科学,2019,39(2):171-180.
[32]苗昊翠,李 强,侯献飞,等. 不同生育期干旱对花生生长发育及产量的影响[J]. 新疆农业科学,2021,58(3):441-449.
[33]李显航,夏志林,喻奇伟,等. 烟草品种不同生育期抗旱性评价方法探讨[J]. 山地农业生物学报,2019,38(1):35-42,92.
[34]张龙龙,杨明明,董 剑,等. 三个小麦新品种不同生育阶段抗旱性的综合评价[J]. 麦类作物学报,2016,36(4):426-434.
