盐度、光强和温度对盐生杜氏藻生长的影响及其交互作用
2021-12-09秦瑞阳李永富刘建国
秦瑞阳, 李永富, 刘建国
盐度、光强和温度对盐生杜氏藻生长的影响及其交互作用
秦瑞阳1, 2, 3, 李永富1, 2, 刘建国1, 2
(1. 中国科学院海洋研究所 实验海洋生物学重点实验室 海洋大科学研究中心, 山东 青岛 266071; 2. 青岛海洋科学与技术试点国家实验室 海洋生物学与生物技术功能实验室, 山东 青岛 266237; 3. 中国科学院大学, 北京 100049)
为明确各环境因子对盐生杜氏藻生长的影响是否存在交互作用及其影响的程度, 采用3因素2水平正交试验方法, 以7天批次培养获得的比生长速率为评价指标, 研究了盐度()、光照强度()、温度()及两因素交互作用、、对细胞增殖速率的影响, 并测定了各条件下光合放氧速率、呼吸耗氧速率、叶绿素荧光及色素含量等以阐释作用机制。结果表明: 温度是影响藻细胞增殖的最显著因素, 其次是盐度与光照, 交互作用×、×对绿色盐藻细胞生长有一定作用, 但影响程度不如单一因子。确定利于盐藻细胞增殖的最优条件为: 温度30 ℃、盐度110、光照强度120 μmol·m–2·s–1, 而利于类胡萝卜素积累的条件则需将盐度提升至160。温度、光照通过影响光合放氧速率, 而盐度通过影响呼吸耗氧速率调控藻细胞生长。
盐生杜氏藻; 正交试验; 比生长速率; 色素含量; 交互作用
盐生杜氏藻()是广盐性单细胞真核绿藻, 特定条件下能大量积累β-胡萝卜素, 是目前天然β-胡萝卜素的主要来源[1]。盐藻还含有丰富的蛋白质, 是海参等水产动物的优质饵料[2], 且其甘油含量最高能达到干重的50%, 具有生产甘油的应用潜力[3]。目前, 澳大利亚、以色列、中国等均已经实现该藻的规模化生产[4]。
盐度、光照和温度是影响盐藻生长的主要非生物环境因子[5-6]。目前, 各因素单独作用时盐藻细胞的生长响应已相对明确: 水温25~30 ℃时盐藻生长最好, 细胞最适光强约100~120 μmol·m–2·s–1[7-8]; 盐度范围因藻株不同而存在差异, 但普遍在盐度高于100时生长较好[9-11]。我们利用盐生杜氏藻(IOCAS 879ss)的实验结果也表明该藻生长的适宜盐度为110~120[12]。然而, 实际生产中盐生杜氏藻生长受多个物理化学因素共同作用, 特别是开放池养殖模式下, 自然强光、高温以及因水分大量蒸发导致的盐度剧升等条件共同影响藻细胞增殖。国内外学者也开始认识到多因素共同作用的问题, 研究重点开始由单一的环境因子影响机制向多因素的共同或交互作用规律转移[6, 13]。Bonnefond等[6]发现, 光照和温度同步波动比光照波动而温度恒定条件下的净固碳速率提升25%, 光照下升高温度对微藻光合固碳具有明显的正向协同作用。刘真等[13]发现高盐和强光均可提高盐藻β-胡萝卜素含量, 而高光高盐联合作用明显要强于单独作用。但是, 影响盐藻生长的各因素中, 其影响程度如何排序, 各因素间是否存在交互作用, 交互作用以及背后蕴含的科学规律, 这些问题尚不清楚。本研究针对以上问题, 围绕多因素共同作用开展盐藻生长响应研究。
本研究以盐生杜氏藻(IOCAS 879ss)为研究对象, 通过正交试验方法考察盐度、光照以及温度之间的交互作用对盐藻生长的影响, 并通过活体叶绿素荧光等方法挖掘各单因素或两因素交互作用的光合生理机制。
1 材料与方法
1.1 藻种培养
盐生杜氏藻(IOCAS 879ss), 由中国科学院海洋研究所藻类与藻类生物技术实验室保存。以改良版Johnson’s培养基[14]培养藻细胞, 通过向培养基中添加NaCl调节盐度。所有藻悬液均在光强为50 μmol·m–2·s–1、温度为(25±1)℃连续预培养4 d, 每天随机调换三角瓶的位置并定时充分摇动3次。
1.2 正交试验设计
以盐度、光照强度和温度3个环境因子为考察因素, 选择正交表L8(27), 设计有交互作用的3因素2水平正交试验(表1), 表头设计见表2。按照表2模拟试验条件, 将预培养4 d至指数生长期的藻种接入已灭菌并冷却的不同盐度培养基中, 分别将藻液培养在100 mL三角瓶中, 每瓶加入50 mL上述藻液, 初始接种密度分别为6.75×104cell·mL–1(培养基盐度110)和1.725×105cell·mL–1(培养基盐度160)。在人工培养箱中连续培养7 d, 光暗比为14 h︰10 h。
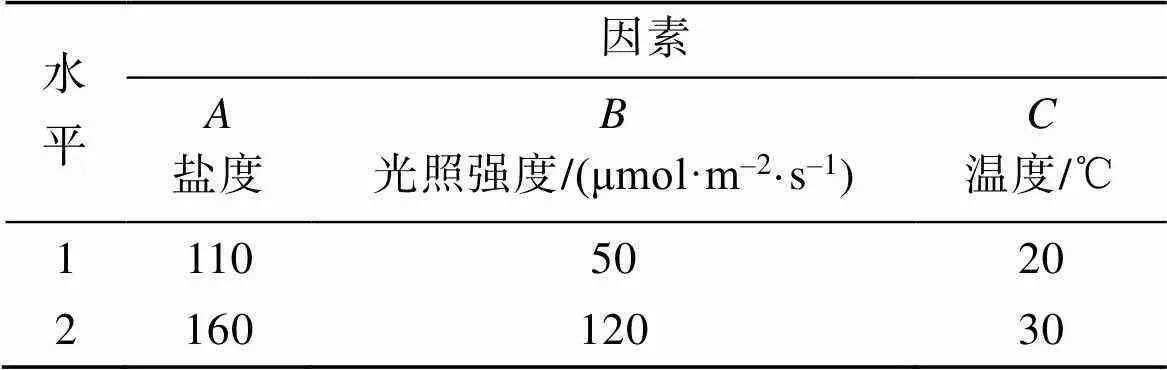
表1 因素水平表
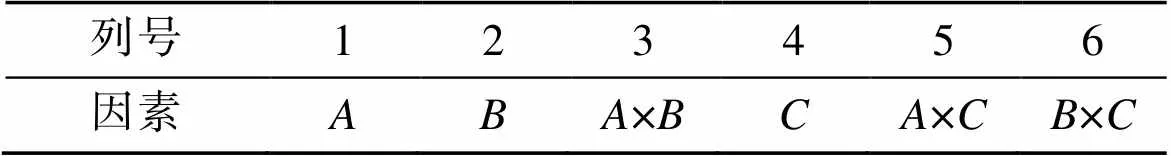
表2 L8 (27) 表头设计
1.3 评价指标
以比生长速率()为评价指标, 对试验结果进行直观分析和方差分析, 确定有显著影响的因子以及有交互作用的试验条件。比生长速率()按下式计算[15]:
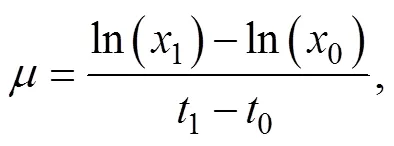
式中,0和1分别表示第0天的(0)和第7天(1)的藻细胞密度(单位: cells·mL–1)。
1.4 色素含量测定
取5 mL藻悬液于10 mL离心管中, 2 770(= 9.8 m/s2)离心5 min, 弃去上清液, 向湿样中加入5 mL 95%乙醇, 旋涡混合后放置在黑暗处4 ℃过夜, 浸泡至藻体为白色, 2 770离心5 min, 用紫外-可见光分光光度计测定液相在665 nm(665nm)、649 nm(649nm)和470 nm(470nm)处吸光度值, 按下式计算色素含量[16]:
a= 13.95665nm– 6.88649nm, (2)
b= 24.96649nm– 7.32665nm, (3)
carot= (1 000470nm– 2.05a– 114.8b)/245,(4)
式中,a、b和carot分别表示叶绿素、叶绿素和类胡萝卜素质量浓度(单位 mg·L–1)。
1.5 藻细胞呼吸耗氧速率和光合放氧速率测定
取2 mL藻液, 加入Chlorolab-2型液相氧电极(Hansatech, 英国)反应室内, 对各试验组藻细胞进行耗氧/放氧速率测定, 并计算其光合和呼吸作用速率[17]: 测定时, 打开磁力转子后首先测定暗处理10 min内氧气的平均消耗速率, 以单个藻细胞产生的氧气量(μmol)进行换算, 即为单个藻细胞的呼吸耗氧速率(μmol·min–1)。打开光源, 在培养光强下照射藻细胞, 测定10 min内氧气平均释放速率, 计算单个藻细胞产生的氧气量(μmol), 即为单个藻细胞的净光合放氧速率(μmol·min–1)。每个处理组重复3次。测定时通过水浴控温保证测定温度与试验温度一致[18]。
1.6 叶绿素荧光参数测定
实际光化学效率(PSII)、最大光化学效率(v/m)、最大天线转换效率(v′m′)、反应中心的开放程度(p)及初始荧光产量(o)使用FMS-2 脉冲调制荧光计(Hansatech, 英国)结合修正后的适配器进行测定[19-20]。测定前将样品置于暗处适应15 min, 测定光强同培养光强, 各参数可在荧光仪上直接读出, 每个处理组重复4次。
1.7 数据处理
所有试验结果均进行3~6次重复, 使用Origin 2018软件作图, 所有数据均用Minitab 18.0进行方差分析, 用方差分析中Games-Howell(方差非齐性)或Tukey(方差齐性)检验方法对组间数据进行显著性差异检验(<0.05)。
2 主要结果
2.1 正交试验条件对盐藻细胞比生长速率的影响
表3和表4为盐藻藻细胞生长的正交试验结果。极差分析和方差分析发现, 温度()和盐度()对比生长速率的影响最明显, 而光照强度()的影响相对较小。其次是盐度和温度交互作用×、光强和温度交互作用×, 最后是盐度和光照交互作用×。盐藻绿色细胞生长最好的环境因子组合为212, 即温度30 ℃、盐度110、光照强度为120 μmol·m–2·s–1。因×和×对绿色藻细胞生长有一定的影响, 进一步对二者分析发现(图1),1与2、2与2配合时, 盐藻绿色细胞比生长速率最大, 与极差分析结果一致(212, 即温度30 ℃、盐度110、光照强度为120 μmol·m–2·s–1)。
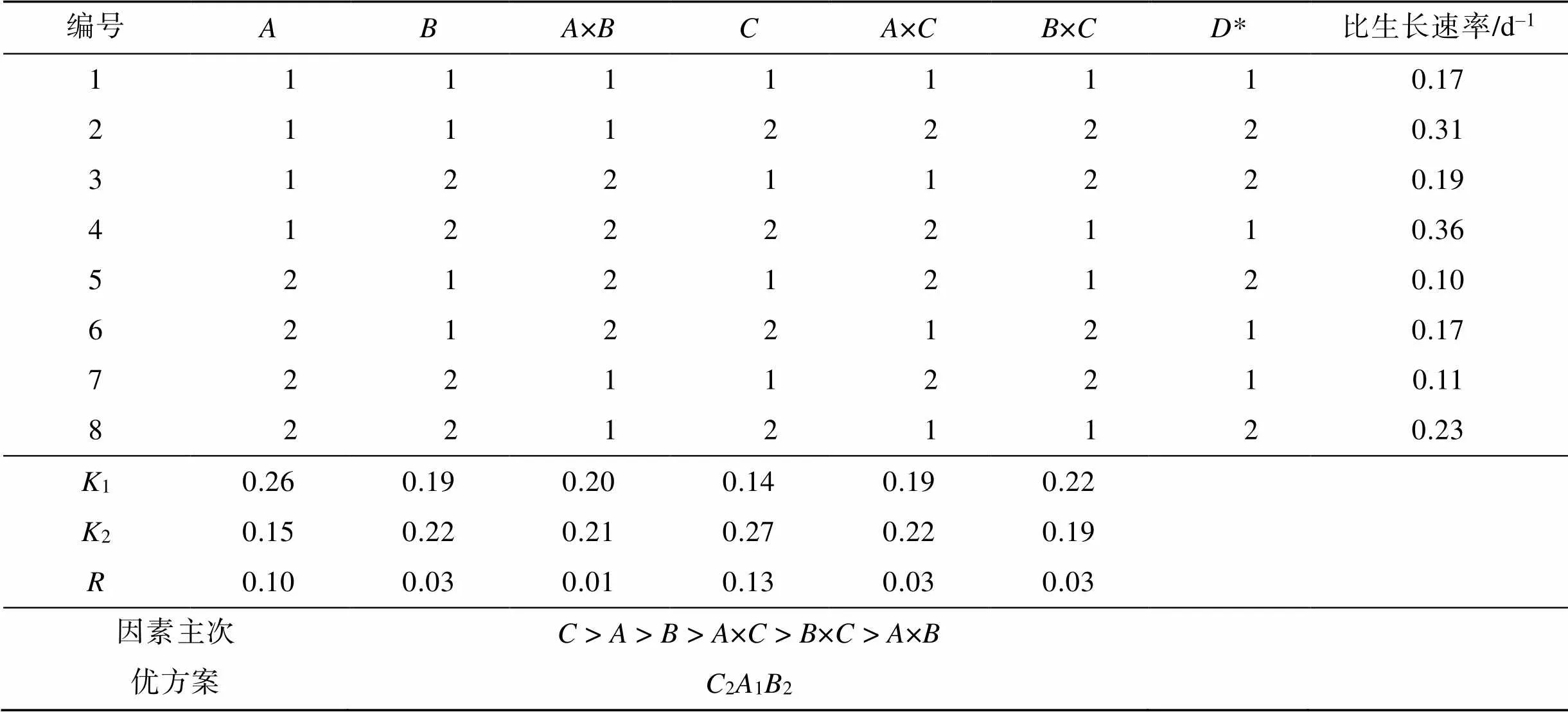
表3 正交试验设计与结果
注: *表示列为空白列[21],1表示各因素在水平1处理条件下的平均值,2表示各因素在水平2处理条件下的平均值,表示各因素的极差。

表4 正交试验结果方差分析
注: 若>0.1, 则< 0.1, 说明处理间有一定差异, 在值右上方标记*-, 若>0.01, 则< 0.01, 说明处理间差异极显著, 在值右上方标记**。
2.2 盐度、光强和温度对细胞色素含量的影响
不同盐度处理下绿色盐藻细胞中色素含量的变化如图2(a)所示。盐度增加, 叶绿素含量和类胡萝卜素含量均显著增加(<0.05)。光照强度由50 μmol·m–2·s–1升至120 μmol·m–2·s–1, 叶绿素、叶绿素含量均显著降低。与20 ℃相比, 30 ℃下藻细胞内叶绿素和类胡萝卜素含量变化差异均不显著(>0.05), 但单位细胞内叶绿素的含量随温度的增加显著增加(<0.05),a/b比值结果由4.96下降至3.67, 意味着温度上升后, 细胞光利用能力提高[22]。
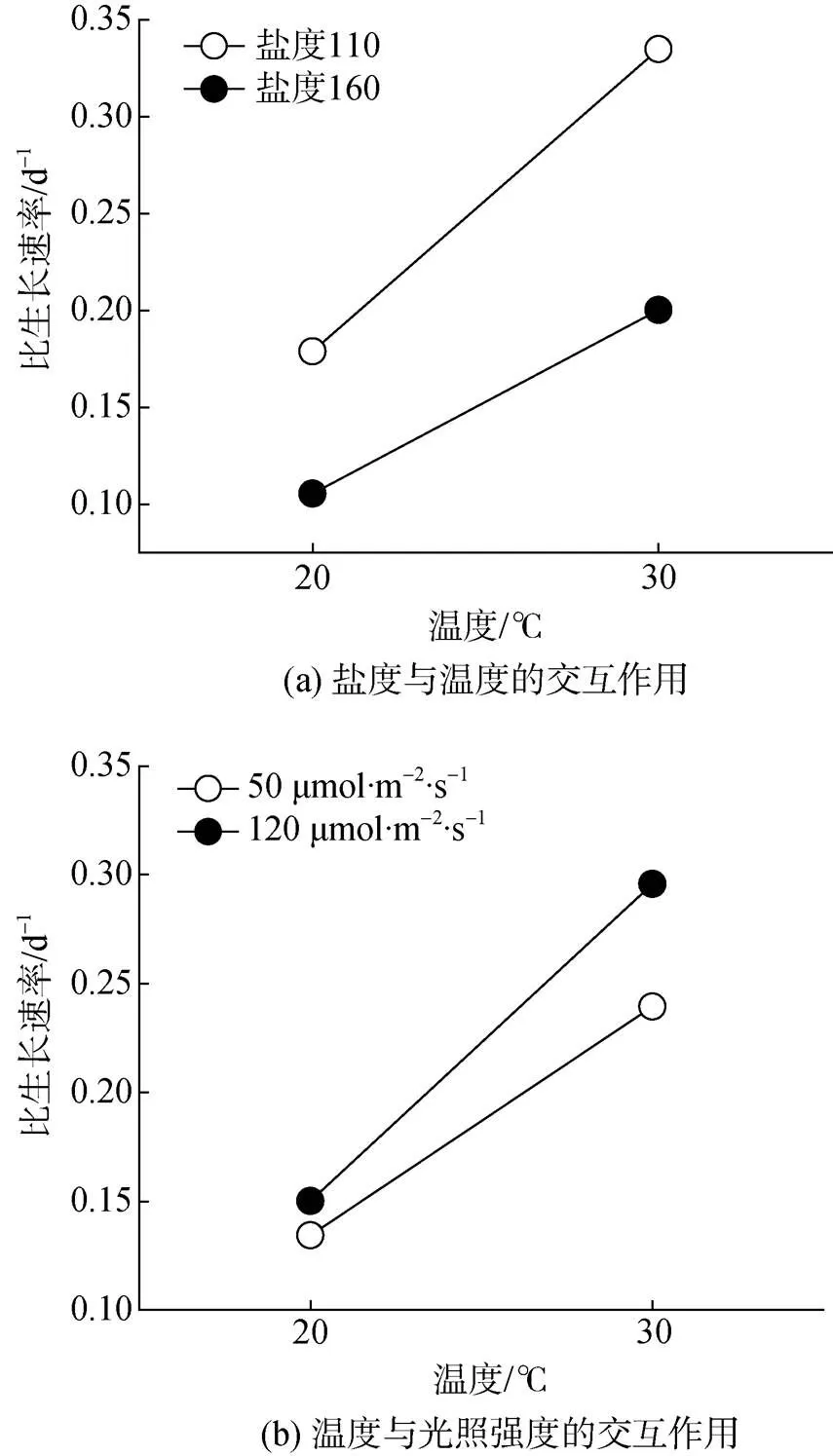
图1 交互作用因素二元图
2.3 盐度、光强和温度对光合放氧及呼吸耗氧速率的影响
图3列出了藻细胞在各因素水平下的呼吸耗氧速率和光合放氧速率变化。可见, 盐藻光合作用过程受光照强度和温度变化影响显著, 而受盐度影响轻微; 但呼吸作用受盐度影响较大, 受另外两因素影响较小。因此, 强光下净光合放氧速率降低应归因于光合作用速率的下降; 而高温下因提高了光合作用速率, 使得净光合放氧速率升高。相应地, 高盐度下出现净放氧速率上升, 主要原因是呼吸速率下降。
2.4 盐度、光强和温度对叶绿素荧光参数的影响
vm,v′m′和PSII分别表示PSII反应中心最大光能转换效率, 最大天线转换效率和实际光化学效率, 反映了不同条件下盐藻细胞的光合生理活性。p是光化学淬灭系数, 表示PSII开放的反应中心所占比例;o为初始荧光, 来自天线色素, 是已经暗适应的光合机构全部PSII中心都开放时的荧光强度[23]。对不同条件下盐藻细胞的测定结果显示(图4), 高、低盐度下vm,v′m′,PSII和p的差异很小, 仅高盐下o升高, 这印证了光合放氧速率和色素的测定结果(图3), 即本研究设置的盐度范围在整体上对光合作用影响较小, 但可提高藻细胞内捕光色素的含量。与盐度影响不同, 光强升高后, 所有参数均明显降低[图4(b)], 光强由50 μmol·m–2·s–1升至120 μmol·m–2·s–1,vm极显著下降(<0.001), 表明藻细胞光合机构所吸收的光能已超出藻细胞本身所能利用的能力, 发生了光抑制[24]。与盐度影响规律类似, 温度升高对反应中心效率参数, 如vm,v′m′,PSII和p的影响较小, 但o明显增加。

图2 不同盐度(110、160)、光照强度(50、120 μmol·m–2·s–1) 和温度(20 ℃、30 ℃)下绿色盐藻细胞色素含量变化
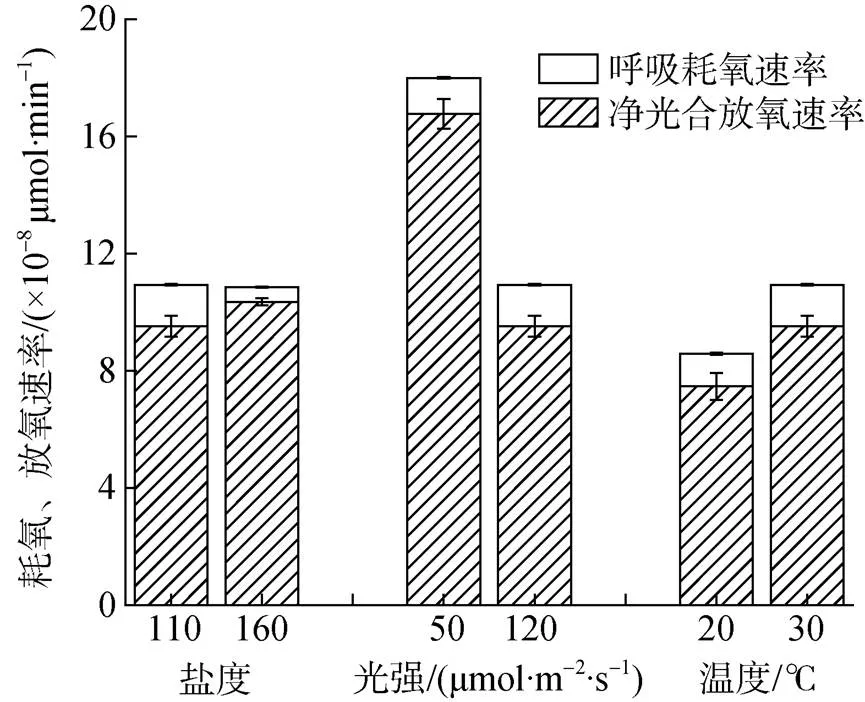
图3 不同盐度(110、160)、光照强度(50、120 μmol·m–2·s–1)和温度(20 ℃、30 ℃)下绿色盐藻细胞呼吸耗氧速率、光合放氧速率
3 讨论
盐生杜氏藻是一种单细胞耐盐绿藻, 可以在接近淡水至饱和盐度下存活[25-26]。本研究表明, 在绿色盐藻增殖阶段, 温度是最重要的影响因素, 其次是盐度, 温度和光照的交互作用、温度和盐度的交互作用分别对绿色盐藻细胞生长有一定作用。当前, 盐藻养殖的大规模商业化培养主要通过户外浅塘(如澳大利亚)或人工跑道池(如印度、中国和以色列)实现, 尚未见利用光生物反应器培养成功的报道[27]。但是, 由于封闭式光生物反应器具有培养条件可控、光径小、利用人工光源补充光能、混合效率高、CO2供应方便等优点[28], 利用反应器进行盐藻培养尝试的报道日益增多[29-31]。例如, Prieto等[29]的研究结果显示, 利用管道反应器开展的盐藻两阶段培养模式, 即先在第一阶段(盐藻生长阶段)提供适宜的生长条件, 待其增殖到一定密度时, 再转入第二阶段(β-胡萝卜素积累阶段), 通过强光、高盐、高温、营养胁迫等手段诱导β-胡萝卜素大量积累, 其生物质产率和β-胡萝卜素产率均明显高于开放池半连续培养模式。

图4 不同盐度(110、160)、光照强度(50、120 μmol·m–2·s–1)和温度(20 ℃、30 ℃)下盐藻细胞叶绿素荧光参数
温度对藻类生长和发育具有重要的调控作用。盐藻在–5 ℃到40 ℃的极端环境条件下均能存活, 但一般认为适宜其快速增殖的温度是25~32 ℃[29, 32-34]。本研究结果支持这一观点, 即30 ℃下盐藻生长速率和净光合放氧速率均高于20 ℃。本研究进一步显示, 高温时藻细胞o明显增加, 这与盐藻的色素含量变化相一致, 即高温通过增加捕光色素含量, 提高光能捕获效率, 最终提高了光合作用速率[35]。与温度影响规律类似, 强光下盐藻细胞比生长速率略有增长, 但单个藻细胞的净光合放氧速率均明显下降, 由1.68× 10–7μmol·min–1降至9.53×10–8μmol·min–1(表3, 图3)。叶绿素荧光分析结果显示, 强光下发生光抑制(图4b)。微藻对光照的需求既与接种密度有关, 又受到单个藻细胞、藻细胞内部捕光色素及其他化学成分消光的影响[36]。在120 μmol·m–2·s–1的强光下, 藻细胞通过减少天线色素含量或反应中心数量来减少对光能的吸收, 从而减少强光照射对光合机构造成的光损伤。但是, 高光下盐藻比生长速率高于弱光, 可能是由于盐藻通过快速分裂增殖, 增加了藻细胞密度, 从而强化细胞间遮挡, 弱化光照对单个细胞造成的损伤。这可能是强光诱发光抑制, 而促进细胞增殖的另一可能原因, 具体机制需更多研究。
有意思的是, 高盐条件下藻细胞的呼吸耗氧速率被显著抑制; 但此时藻细胞内色素含量上升(图2a),净光合放氧速率升高(图3), 但比生长速率降低(表3)。这一现象说明, 在本试验盐度范围内, 相对低的盐度有利于藻细胞的增殖(表3), 而在高盐度条件下有利于类胡萝卜素的积累, 这与Ahmed等的研究结果一致[37-40]。盐藻在高盐下会大量积累甘油, 且胞内甘油含量与外部渗透压成正比[41]。单位体积内和单个细胞的净光合放氧速率均明显上升, 说明生物质积累增加, 且增加可能因甘油积累所致。光合作用和呼吸作用活性分析结果进一步说明, 高盐条件下甘油积累的还原力和底物来源是光合作用, 被显著抑制的呼吸耗氧所提供的ATP和还原力难以维持甘油含量的瞬时提高[42]。因此, 从细胞密度角度考虑, 应选用盐度110; 而从生物量积累的角度考虑, 宜选择盐度160。以上选择取决于由第一阶段(即细胞增殖阶段)向第二阶段转移(即类胡萝卜素积累阶段)时是否采用高盐胁迫条件。若存在盐度升高条件, 往往因大量积累甘油而导致生物量提高[43-44], 此时提供尽可能多的藻细胞有利于最终生物质的产出, 选择110盐度进行绿色细胞增殖优于160。反之, 若无盐度明显变化, 宜选用盐度为160。一般而言, 现有报道的二步法培养, 均为在低盐条件下使盐藻快速增殖到一定细胞密度后再转入高盐条件下使其积累大量的β-胡萝卜素, 从而达到高密度培养和高β-胡萝卜素积累, 尚未见两阶段盐度一致的报道[5, 45]。因此, 确定110为生产选用盐度为宜。
除单因素作用外, 本研究还发现盐度-温度交互作用和温度-光照交互作用能明显影响盐藻的细胞增殖, 具体表现为: 在低盐条件下, 高、低温之间的比生长速率差异明显高于高盐条件下; 而高温条件更利于通过提升光强加速盐藻细胞增殖(图1)。单因素分析发现, 低盐条件下, 藻细胞的o明显降低, 叶绿素和类胡萝卜素含量均低于高盐条件下的含量; 而30 ℃与20 ℃相比, 叶绿素明显升高。低盐条件下, 因温度升高而色素含量上调, 最终导致藻细胞的捕光能力高于高盐条件下, 这可能是低盐条件下, 高-低温增殖速率出现明显差异的原因(图1)。其次, 高-低温条件下, 藻细胞最大光化学效率vm差异较小, 但强光下调了该参数和总光合放氧速率(图3, 图4)。30 ℃高温尚不足以对藻细胞产生热损伤, 反而有利于光合作用进行(图3)。高温条件下, 强光对藻细胞产生的光抑制程度明显降低, 这可能是高温条件下强-弱光藻细胞比生长速率差异大于低温条件下强-弱光藻细胞比生长速率差异的原因。值得一提的是, 本研究发现盐度和光照的交互作用对藻细胞增殖影响较小, 但不足以说明这种交互作用不存在。如前所述, 本研究所设置的盐度范围(110~160)均是盐藻的适宜生长条件, 强、弱光对藻细胞比生长速率的影响差异在不同盐度下尚未显现。理论上, 光照和盐度分别是调控β-胡萝卜素合成和甘油代谢的关键因素, 而β-胡萝卜素合成和甘油需共同的光合作用产物丙酮酸[46-47], 虽然尚无光照强度直接影响甘油代谢途径的生物学证据, 但光照至少可通过影响光合作用强度间接调控甘油代谢。光盐交互作用是否会在增大盐度梯度范围时显现尚需更多研究。
4 结论
1) 在设置的水平范围内, 各因素对藻细胞比生长速率的影响次序为温度>盐度>光照; 最适细胞增殖条件为: 盐度110、光照强度120 μmol·m–2·s–1、温度30 ℃。若以获取生物量为培养目的, 盐度提升至160为宜。
2) 在被前人优选的培养条件下, 盐度通过影响呼吸耗氧速率调控藻细胞生长, 而温度、光照主要通过影响光合放氧速率起作用。盐度和温度交互作用(), 光照和温度交互作用(×)对藻细胞增殖均有影响。这可能与温度上升时色素含量升高, 捕光能力和光合放氧速率上调有关。
[1] TAFRESHI A H, SHARIATI M.biotechnology: methods and applications[J]. Journal of Applied Microbiology, 2010, 107(1): 14-35.
[2] ORSET S, YOUNG A J. Low-temperature-induced synthesis of α-carotene in the microalga(Chlorophyta)[J]. Journal of Phycology, 2010, 35(3): 520-527.
[3] ROCHE J L, BERTRAND A M. Light intensity-induced changes in cab mRNA and light harvesting complex II apoprotein levels in the unicellular chlorophyte[J]. Plant Physiology, 1991, 97: 147-153.
[4] BERNER T, DUBINSKY Z, WYMAN K, et al. Photoadaptation and the ‘Package’ effect on(Chlorophyceae)[J]. Journal of Phycology, 2010, 25(1): 70-78.
[5] 周世水, 姜建国, 林炜铁, 等. 盐藻生长及其β-胡萝卜素累积的作用因子和最适条件探讨[J]. 食品与发酵工业, 2002, 28(11): 1-3.
ZHOU Shishui, JIANG Jianguo, LIN Weitie, et al. The determination of the optimum conditions of growth and β-carotene accumulation for[J]. Food and Fementation Industries, 2002, 28(11): 1-3.
[6] BONNEFOND H, MOELANTS N, TALEC A, et al. Concomitant effects of light and temperature diel variations on the growth rate and lipid production of[J]. Algal Research, 2016, 14: 72-78.
[7] GÓMEZ P I, GONZÁLEZ M A. The effect of temperature and irradiance on the growth and carotenogenic capacity of seven strains of(Chlorophyta) cultivated under laboratory conditions[J]. Biological Research, 2005, 38(2/3): 151-162.
[8] 张学成, 孟振, 时艳侠, 等. 光照、温度和营养盐对三株盐生杜氏藻生长和色素积累的影响[J].中国海洋大学学报(自然科学版), 2006, 36(5): 754-762.
ZHANG Xuecheng, MENG Zhen, SHI Yanxia, et al. The effect of light, temperature and nutrition on growth and pigment accumulation of three strains of[J]. Periodical of Ocean University of China, 2006, 36(5): 754-762.
[9] FUGGI A, POLLIO A. Effects of NaCl, Na2SO4, H2SO4and glucose on growth, photosynthesis, and respiration in the acidophilic algae[J]. Phycologia, 1988, 27(3): 334-339.
[10] NELLIS M, FRANCISCO M, ERIC T. Effect of nitrate concentration on growth and pigment synthesis ofcultivated under low illumination and preadapted to different salinities[J]. Journal of applied phycology 1998, 10: 405-411.
[11] CHEN H, JIANG J G, WU G H. Effects of salinity changes on the growth ofand its isozyme activities of glycerol-3-phosphate dehydrogenase[J]. Journal of Agricultural and Food Chemistry, 2009, 57(14): 6178-6182.
[12] 秦瑞阳. 绿色盐藻细胞对强光高盐的生理响应及其胁迫缓解方法研究[D]. 北京: 中国科学院大学, 2020.
QIN Ruiyang. Study on the physiological response ofto strong light and hypersaline and the methods to alleviate the stress[D]. Beijing: University of Chinese Academy of Sciences, 2020.
[13] 刘真, 刘姗姗, 张玉洁, 等. 光照强度及盐浓度对杜氏盐藻生长及β-胡萝卜素积累的影响[J]. 郑州大学学报(医学版), 2016, 51(1): 31-35.
LIU Zhen, LIU Shanshan, ZHANG Yujie, et al. Effects of light intensity and salt concentration on growth ofand accumulation of β-carotene[J]. Journal of Zhengzhou University (Medical Sciences), 2016, 51(1): 31-35.
[14] 刘建国, 赵学武, 王玉君, 等. 胁迫条件下盐藻β-胡萝卜素及其异构体累积的研究-盐度的影响[J]. 海洋与湖沼, 1994, 25(1): 71-76.
LIU Jianguo, ZHAO Xuewu, WANG Yujun, et al. Study on accumulation of β-carotene and its isomers inunder environmental stress conditions- salinity effect[J]. Oceanologia et Limnologia Sinica, 1994, 25(1): 71-76.
[15] 杨宋琪, 史邵华, 王丽娟, 等. 杜氏盐藻生长及PSⅡ对不同磷源的响应[J]. 海洋科学, 2016, 40(10): 1-7.
YANG Songqi, SHI Shaohua, WANG Lijuan, et al. Response of growth and Photosystem Ⅱ ofon different phosphorus sources[J]. Marine Sciences, 2016, 40(10): 1-7.
[16] 蔡永萍. 植物生理学实验指导[M]. 北京: 中国农业大学出版社, 2014.
CAI Yongping. Experimental guidance of Plant Physiology[M]. Beijing: China Agricultural University Press, 2014.
[17] 袁朝杰, 张莹, 李涛, 等. 迦得拟微球藻在不同硝酸钾浓度下的生长和生理生化特征[J]. 海洋与湖沼, 2019, 50(1): 110-119.
YUAN Chaojie, ZHANG Ying, LI Tao, et al. The growth, physiological and biochemical characteristics ofin different potassium nitrate concentration[J]. Oceanologia et Limnologia Sinica, 2019, 50(1): 110-119.
[18] LI Y F, LIU J G, ZHANG L T, et al. Effects of temperature on the photosynthetic performance in mature thalli of the red alga(Gelidiaceae)[J]. Aquaculture, 2019, 512: 734320.
[19] ZHANG L T, ZHANG Z S, GAO H Y, et al. Mitochondrial alternative oxdiase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species inK-1 leaves[J]. Physiology Plant, 2011, 143: 396-407.
[20] ZHANG L T, ZHANG Z S, GAO H Y, et al. The mitochondrial alternative oxidase pathway protects the photosynthetic apparatus against photodamage inK-1 leaves[J]. BMC Plant Biology, 2012, 12: 40.
[21] 李春喜, 姜丽娜, 邵云, 等. 生物统计学(第五版)[M]. 北京: 科学出版社, 2013: 1-298.
LI Chunxi, JIANG Lina, SHAO Yun, et al. Biostatistics (Fifth Edition)[M]. Beijing: Science Press, 2013: 1-298.
[22] 赵新宇. 漂浮状态浒苔()光合系统对典型环境变化的适应特征及其机理的研究[D]. 青岛: 中国海洋大学, 2015.
ZHAO Xinyu. Study of adaptive features and mechanism of photosynthetic system in Floatingagainst typical environmrntal changes[D]. Qingdao: Ocean University of China, 2015.
[23] 李钦夫, 李征明, 纪建伟, 等. 叶绿素荧光动力学及在植物抗逆生理研究中的应用[J]. 湖北农业科学, 2013, 52(22): 5399-5402.
LI Qinfu, LI Zhengming, JI Jianwei, et al. Applications of chlorophyll fluorescence kinetics in the physiological resistance studies of plant[J]. Hubei Agricultural Sciences, 2013, 52(22): 5399-5402.
[24] 陈建明, 俞晓平, 程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报, 2006, 18(1): 51-55.
CHEN Jianming, YU Xiaoping, CHEN Jiaan. The application of chlorophyll fluorescence kinetics in the study of physiological responses of plants to environmental stresses[J]. Acta Agriculturae Zhejiangensis, 2006, 18(1): 51-55.
[25] BEN-AMOTZ A, AVRON M. The biotechnology of cultivating the halotolerant alga Dunaliella[J]. Trends in Biotechnology, 1990, 8(5): 121-126.
[26] BEN-AMOTZ A, AVRON M. Glycerol and β-carotene metabolism in the halotolerant alga: a model system for biosolar energy conversion[J]. Trends in Biochemical Sciences, 1981, 6: 297-299.
[27] BOROWITZKA M A.: Biology, Production, and Markets[M]//Handbook of Microalgal Culture: Applied Phycology and Biotechnology. Second Edition. USA: John Wiley & Sons, Ltd, 2013: 359-368.
[28] TREDICI M R. Mass Production of Microalgae: Photobioreactors[M]//Handbook of Microalgal Culture: Biotechnology and Applied Phycology. Oxford: Blackwell Publishing Ltd, 2004: 178-214.
[29] PRIETO A, CAÑAVATE J P, GARCÍA-GONZÁLEZ M. Assessment of carotenoid production byin different culture systems and operation regimes[J]. Journal of Biotechnology, 2011, 151(2): 180- 185.
[30] GARCÍA-GONZÁLEZ M, MORENO J, MANZANO J C, et al. Production ofbiomass rich in 9-cis-β-carotene and lutein in a closed tubular photobioreactor[J]. Journal of Biotechnology, 2005, 115(1): 81-90.
[31] HEJAZI M A, ANDRYSIEWICZ E, TRAMPER J, et al. Effect of mixing rate on beta-carotene production and extraction byin two-phase bioreactors[J]. Biomolecular Engineering, 2003, 20(4): 591-596.
[32] WU Z, DUANGMANEE P, ZHAO P, et al. The effects of light, temperature, and nutrition on growth and pigment accumulation of threestrains isolated from saline soil[J]. Jundishapur Journal of Microbiology, 2016, 9(1): e26732.
[33] SIEGEL B Z, SIEGEL S M, SPEITEL T, et al. Brine organisms and the question of habitat-specific adaption[J]. Origins of Life, 1984, 14: 757-770.
[34] BOROWITZKA L J. The microflora. Adaptions to life in extremely saline lakes[J]. Hydrobiologia, 1981, 81: 33-46.
[35] MATORIN D N, ANTAL T K, OSTROWSKA M, et al. Chlorophyll fluorimetry as a method for studying light absorption by photosynthetic pigments in marine algae[J]. Oceanologia, 2004, 46(4): 519-531.
[36] LEHMUSKERO A, CHAUTON M S, BOSTROM T. Light and photosynthetic microalgae: a review of cellular- and molecular-scale optical processes[J]. Progress in Oceanography, 2018, 168: 43-56.
[37] AHMED R A, HE M, AFTAB R A, et al. Bioenergy application ofSA 134 grown at various salinity levels for lipid production[J]. Scientific Reports, 2017, 7(1): 8118.
[38] FARHAT N, RABHI M, FALLEH H, et al. Optimization of salt concentrations for a higher carotenoid production in(Chlorophyceae)[J]. Journal of Phycology, 2011, 47(5): 1072-1077.
[39] ARAÚJO O Q, GOBBI C N, CHALOUB R M, et al. Assessment of the impact of salinity and irradiance on the combined carbon dioxide sequestration and carotenoids production by: a mathematical model[J]. Biotechnology & Bioengineering, 2009, 102(2): 425-435.
[40] BOROWITZKA M A, BOROWITZKA L J, KESSLY D. Effects of salinity increase on carotenoid accumulation in the green alga[J]. Journal of Applied Phycology, 1990, 2(2): 111-119.
[41] 周丽, 孟祥红, 刘成圣, 等. 渗透胁迫对杜氏盐藻胞内甘油含量及相关酶活性影响[J]. 植物学通报, 2006, 23(2): 145-151.
ZHOU Li, MENG Xianghong, LIU Chengsheng, et al. Effects of osmotic stress on intracellular glycerol content and enzyme activity in[J]. Chinese Bulletin of Botany, 2006, 23(2): 145-151.
[42] XU Y N, IBRAHIM I M, HARVEY P J. The influence of photoperiod and light intensity on the growth and photosynthesis of(chlorophyta) CCAP 19/30[J]. Plant Physiology and Biochemistry, 2016, 106: 305-315.
[43] 孙晓菲, 黄非, 梁雪, 等. 不同胁迫下盐生杜氏藻3-磷酸甘油脱氢酶基因表达及其与甘油合成的响应[J]. 四川大学学报: 自然科学版, 2007, 44(2): 433-438.
SUN Xiaofei, HUANG Fei, LIANG Xue, et al. Expression of GPD gene fromtreated with different stress and glycerol synthesis of the cells[J]. Journal of Sichuan University (Natural Science Edition), 2007, 44(2): 433-438.
[44] 陈慈美, 方志山, 郑晓玲, 等. 盐藻生产β-胡萝卜素两阶段养殖新模式研究[J]. 海洋通报, 1998, 17(2): 28-36.
CHEN Cimei, FANG Zhishan, ZHENG Xiaoling, et al. A study of new mode of two-phase culture offor producing β-carotene[J]. Marine Science Bulletin, 1998, 17(2): 28-36.
[45] BEN-AMOTZ A. New mode ofbiotechnology: two-phase growth for β-carotene production[J]. Journal of Applied Phycology, 1995, 7(1): 65-68.
[46] NISAR N, LI L, LU S, et al. Carotenoid metabolism in plants[J]. Molecular Plant, 2015, 8(1): 68-82.
[47] CHEN H, JIANG J G. Osmotic responses ofto the changes of salinity[J]. Journal of Cellular Physiology, 2009, 219(2): 251-258.
Effects of salinity, light, and temperature and their interactions ongrowth
QIN Rui-yang1, 2, 3, LI Yong-fu1, 2, LIU Jian-guo1, 2
(1. CAS Key Laboratory of Experimental Marine Biology, Center for Ocean Mega-Science, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266237, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China)
To confirm the effects of salinity (), light intensity (), and temperature () and interactions between the factors, namely×,×, and×, on the growth ofand the degree of interaction, a three-factor, two-level orthogonal experiment was conducted. The specific growth rate was obtained from eight groups of batch cultures grown for 7 days at different factor combinations as an evaluated index. Rates of photosynthetic oxygen evolution and respiratory oxygen consumption, chlorophyll fluorescence, and pigment content were also measured to clarify the physiological mechanism of the interactions. The results revealed that temperature affected algal growth most significantly, followed by salinity and light intensity. Interactions×and×played important roles in regulating the growth of; however, the degree of influence was not better than that of a single factor. Optimal conditions for algal proliferation included temperature 30 ℃, salinity 110, and light intensity 120 μmol·m−2·s−1, whereas optimal salinity for carotenoid accumulation was 160. Moreover, both temperature and light intensity primarily regulatedgrowth by affecting the rate of photosynthetic oxygen evolution, whereas salinity affected respiratory rate.
; orthogonal experiment; specific growth rate; pigment content; interaction
Aug. 5, 2020
S917.3
A
1000-3096(2021)11-0073-09
10.11759/hykx20200805001
2020-08-05;
2020-09-07
国家自然科学基金项目(32002411, U1706209)
[National Natural Science Foundation of China, No. 32002411, No. U1706209]
秦瑞阳(1992—), 女, 山东日照人, 研究生, 硕士, 主要从事藻类生理生化过程与调控研究, E-mail: qinruiyang@qdio.ac.cn; 刘建国(1964—),通信作者, 男, 山东寿光人, 研究员, 博士, 主要从事藻类和藻类生物技术研究, 电话: 0532-82898709, E-mail: jgliu@qdio. ac.cn
(本文编辑: 丛培秀)
