碳对微生物–根系介导的蔬菜作物磷吸收的影响
2021-12-05李洪亮诸海焘徐四新蔡树美付子轼张德闪
李洪亮,诸海焘,徐四新,蔡树美,付子轼,张德闪
(上海市农业科学院生态环境保护研究所/农业农村部上海农业环境与耕地保育科学观测实验站/上海市设施园艺技术重点实验室, 上海 201403)
在集约化农业生产中,经常施用大量的磷肥以满足作物生长,实现产量的增加[1]。然而施入的大部分磷肥会被土壤固定,成为作物难以利用的无效态磷[2]。微生物是土壤磷循环的重要驱动因子,微生物磷周转在调节磷有效性方面起着重要的作用[3]。土壤磷有效性的变化会诱导植物调节根系形态与根际分泌物的释放,以形成高效的磷吸收策略[4]。例如,当土壤有效磷含量降低时,植物通过增加总根长,降低根系直径(增加比根长),提高根系对土壤有效磷的吸收[5–6],或者通过增强酸性磷酸酶和有机酸(如柠檬酸和苹果酸)的分泌,活化土壤难溶性有机磷和无机磷,提高土壤磷有效性[4,7–8]。然而,有关作物如何调节根系形态和根际分泌物响应微生物介导的磷周转尚不明确。
增添有机碳源可以为土壤微生物提供生长所需的能量,通过刺激微生物的增生/死亡,启动微生物介导的磷周转[9–10]。然而,微生物磷周转对磷有效性的调节作用取决于土壤碳的有效性[11]。例如,添加碳源初期,供微生物生长繁殖的有效碳含量显著提高,微生物快速生长,微生物对磷的吸收因此增加,导致土壤有效磷含量降低[12–13],微生物与根系之间对磷的竞争加剧。随着微生物呼吸作用对碳的耗竭,土壤中供微生物利用的有效碳含量逐渐下降,微生物因碳源缺乏而死亡并向土壤重新释放磷资源,从而增加了植物可利用的有效磷含量[14]。微生物磷周转诱导的土壤磷有效性的动态变化,可能会引起作物根系形态和根际分泌物的动态响应,影响作物磷吸收[15–16]。然而,外源有机碳源添加后有关微生物磷周转对土壤磷有效性的影响,以及介导作物磷吸收的根系/根际属性的研究仍然缺少系统的分析。研究微生物–根系互作的动态过程及其对作物磷吸收的影响,对于探索强化微生物磷周转协同根系提高磷利用效率的根际调控措施至关重要。
磷肥的过量施用是集约化蔬菜种植体系磷肥管理中普遍存在的问题[17]。由于蔬菜根系小而浅,磷的利用效率低,大量的磷资源残留在菜田土壤[17],导致土壤碳/磷失衡,限制了微生物介导的磷周转对磷的活化作用[18–19]。鉴于土壤有机碳源的供给能够刺激微生物增生,影响微生物代谢活动。因此,我们假设向土壤添加有机碳源可以启动微生物介导的磷周转,调节土壤磷有效性,从而影响根系形态、根际分泌物以及作物地上部磷吸收。本研究以绿叶菜上海青作为研究对象,以葡萄糖作为有机碳源,利用盆栽模拟试验,研究外源碳调控对微生物–根系互作过程及其介导的作物磷吸收的影响,探索强化微生物–根系互作,提高磷肥利用效率的根际管理措施,为集约化蔬菜种植体系磷肥的减施提供理论依据。
1 材料与方法
1.1 试验设计与材料
试验设置添加葡萄糖(+G)和不加葡萄糖(−G)两个处理,每个处理8次重复,分两次采样,分析添加葡萄糖后土壤微生物量磷、有效磷(Olsen-P)、根系(根系生物量、根冠比、根长、直径、比根长和根系组织密度)/根际属性(磷酸酶、柠檬酸和苹果酸)特征以及作物磷吸收的动态变化。供试土壤取自上海市农业科学院庄行实验站稻田土壤(30°53′N,121°22′E),土壤类型为潜育土,其基础理化性质如下:容重 1.41 g/cm3、有机碳 16.7 g/kg、全氮 1.2 g/kg、全磷 0.7 g/kg、无机氮 37 mg/kg、有效磷 9 mg/kg、速效钾 94 mg/kg、pH 为 6.8 (土水比 1∶5)。供试土壤风干过 2 mm 筛,每个塑料花盆 (长×宽×高 = 17 cm ×9 cm × 9 cm)装 1.5 kg 的土壤。所有处理均不加外源磷,为保证作物正常生长,所有处理添加作物生长的其他必需养分,每盆土壤的供应量分别为:Ca(NO3)2·4H2O 250 mg、K2SO4267 mg、MgSO4·7H2O 87 mg、Fe-EDTA 11.7 mg、MnSO4·H2O 13.3 mg、ZnSO4·7H2O 20 mg、CuSO4·5H2O 4 mg、H3BO32.7 mg、Na2MoO4·5H2O 0.33 mg。将土壤和养分充分混匀,用称重法按田间持水量的75%浇水,平衡3天后进行播种。
供试作物为上海青(油菜),品种为夏青3号(Brassica chinensisL. Xiaqing3),由上海市农业科学院园艺所提供。试验开始前,将种子泡入10% H2O2(v/v)中浸泡20 min消毒,用去离子水反复冲洗干净,随后将种子浸泡在饱和CaSO4溶液中,在避光通气条件下浸泡12 h;随后将吸胀后的种子摆放在垫有湿润滤纸的托盘上,在恒温培养箱中23℃条件下避光催芽2天。每盆播4颗种子,待幼苗“两叶一心”后间苗,留2棵长势一致的幼苗。
本试验于上海市农业科学院庄行实验站的温室大棚内进行。预试验结果表明,葡萄糖的添加量为C 0.2 g/(kg·土)时,作物生长状态正常。在上海青种植14天后,添加葡萄糖,葡萄糖的加入量为C 0.2 g/(kg·土),按田间持水量的75%配置葡萄糖溶液,用注射器注入土壤。基于土壤微生物量C∶N平均值为8∶1,为避免微生物氮矿化所导致的土壤速效氮含量的变化对作物根系的影响[20-21],在添加葡萄糖的同时按 N 25 mg/(kg·土)的比例添加 Ca(NO3)2·4H2O溶液,不加葡萄糖的处理加入等量的Ca(NO3)2·4H2O溶液。试验期间温室平均温度维持在白天21℃~33℃,夜晚16℃~23℃。每天采用称重法浇水,保持田间持水量的75%。
1.2 样品采集与项目测定
上海青生长期为40天,在葡萄糖添加后的第7天(营养生长第21天)和第21天(营养生长第35天)分两次采集植株、土壤、根系和根际样品,此时作物处于快速的营养生长期。收获时,沿着茎基部将植株地上部剪下,将剪下的植株地上部称重,即为地上部鲜重。将根系从土壤中取出,轻轻抖动去掉松散的土体土壤,将黏附有根际土的根系置于装有 50 mL 0.2 mmol/L CaCl2溶液中摇动 1 min得到根际土悬浊液[22],吸取8 mL悬浊液至10 mL离心管,加入2滴微生物抑制剂避免微生物代谢对根际碳源物质的消耗,于−20℃下保存,用高效液相色谱法测定有机酸[23]。吸取2份0.5 mL的悬浊液至2 mL离心管中,加入0.1 mL对硝基苯磷酸二钠,0.4 mL乙酸–乙酸钠缓冲液和0.5 mL去离子水,其中一份加入0.5 mL 0.5 mol/L氢氧化钠溶液,一份不加;30℃条件下培养30 min取出,向未加氢氧化钠装有样品的离心管加入0.5 mL 0.5 mol/L氢氧化钠溶液终止反应,在405 nm波长下比色,计算植物根际酸性磷酸酶活性[24]。
采集两份土壤样品,过2 mm筛充分混匀,一份自然风干用于测定土壤有效磷含量,另一份于−20℃下保存,用于测定微生物量碳、磷含量。土壤微生物量碳(MBC)采用氯仿熏蒸—K2SO4浸提法测定[25]:称取相当于10 g烘干基重的新鲜土壤2份,其中1份用无乙醇氯仿避光熏蒸,另外一份不加氯仿作为对照按同样方法培养,24 h 后用 0.5 mol/L K2SO4浸提(土水比为1∶4),采用TOC-VCHP有机碳分析仪测定微生物量碳含量。微生物量碳计算公式为 MBC =EC/kC,其中,EC为熏蒸后的土壤碳含量与未熏蒸土壤碳含量的差值,kC为转化系数0.45。土壤微生物量磷(MBP)采用氯仿熏蒸—NaHCO3浸提法测定[26]:称取相当于4 g烘干基重的新鲜土壤9份,其中3份用无乙醇氯仿避光熏蒸,另外6份不加氯仿作为对照按同样方法培养,24 h后氯仿熏蒸以及无氯仿培养的 3 份土壤用 0.5 mol/L NaHCO3直接浸提 (土水比为1∶20),钼蓝法测定土壤有效磷含量,另外3份无氯仿培养的土壤样品在用 0.5 mol/L NaHCO3浸提的同时加入 0.5 mL 含 P 250 μg/mL 的 KH2PO4溶液,钼蓝法测定土壤有效磷含量,计算外加正磷酸盐态无机磷(Pi)的回收率。微生物量磷计算公式为MBP= EPi/(kP× RPi),其中 EPi为熏蒸土壤 Pi与未熏蒸土壤 Pi含量的差值,RPi= (加 Pi的土壤提取的 Pi –未熏蒸土壤提取的 Pi)/ 25 × 100%,kP为转换系数,取值0.4。Olsen法测定土壤有效磷含量[27]。
将地上部样品放入烘箱105℃杀青30 min,然后65℃下烘干至恒重,测量地上部干重,粉碎,用浓硫酸–双氧水法消煮,钼黄显色法测定消煮液磷浓度,计算地上部磷吸收量[植株地上部磷吸收量(mg/plant)=植株地上部干重(g/plant) × 植株地上部磷含量 (%) × 1000]。
将根系用清水冲洗干净,用根系扫描仪(Epson Expression 1600 pro, Model EU-35, Tokyo, Japan)扫描全部根系,用WinRhizo软件处理扫描图像,计算总根长、根直径和根体积,扫描完成后将根系65℃下烘干至恒重,测根系干重,计算根系比根长、组织密度和根冠比。
1.3 数据分析
用Studentt-test 检验法分析葡萄糖对上海青地上部鲜重、干重、磷吸收以及根系生物量和总根长的影响,同时分别分析葡萄糖和取样时间对作物根系(根冠比、根系生长速率、平均直径、比根长和根系组织密度)/根际属性(酸性磷酸酶、柠檬酸和苹果酸)以及土壤指标(微生物量碳、微生物量磷和Olsen-P)的影响。按以下公式,计算两个取样期内植株地上部、根系/根际属性以及土壤相关指标对葡萄糖的响应比(RR)[28]:
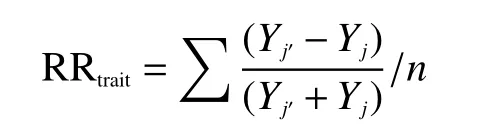
式中:j′和j分别代表+G处理和–G处理中4个重复的其中一个样本;Yj′和Yj为对应的样本中植物地上部、根系和土壤指标的数值;n为Yj′−Yj的组合数,在本试验中因为处理数为4,所以n为16。响应比为正说明葡萄糖的添加对该指标表现为促进效应,为负则说明添加葡萄糖对该指标为抑制效应,当添加葡萄糖的影响不显著时,不进行响应比的计算。用Studentt-test检验法分析葡萄糖添加后第7天与第21天响应比的差异。使用Canoco 4.5软件进行冗余分析(RDA),分析土壤微生物量磷以及Olsen-P含量对上海青根系/根际属性特征的影响。
2 结果与分析
2.1 添加葡萄糖对上海青地上部鲜重、干重和磷吸收的影响
葡萄糖添加第7天和第21天的上海青地上部鲜重、干重和磷吸收量在与不添加葡萄糖处理的差异不显著(图1)。

图1 添加葡萄糖第7天和第21天上海青的地上部鲜重、干重和含磷量Fig. 1 Shoot fresh weight, dry weight and P content of Brassica chinensis at 7 and 21 days after glucose addition
2.2 添加葡萄糖对介导作物磷吸收的根系/根际属性的影响
添加葡萄糖对上海青根系生物量、根冠比以及总根长有显著的影响(图2)。添加葡萄糖后第7天,上海青根系生物量、根冠比显著增加,根系总根长降低;与不加葡萄糖处理(−G)相比,添加葡萄糖后上海青根系生物量增加了26%,总根长减少33%。从第7天到第21天,添加葡萄糖导致上海青根冠比增加。第21天,与不加葡萄糖的对照处理(−G)相比,添加葡萄糖促进根系生物量增加,根冠比提高,根系总根长增加31%。
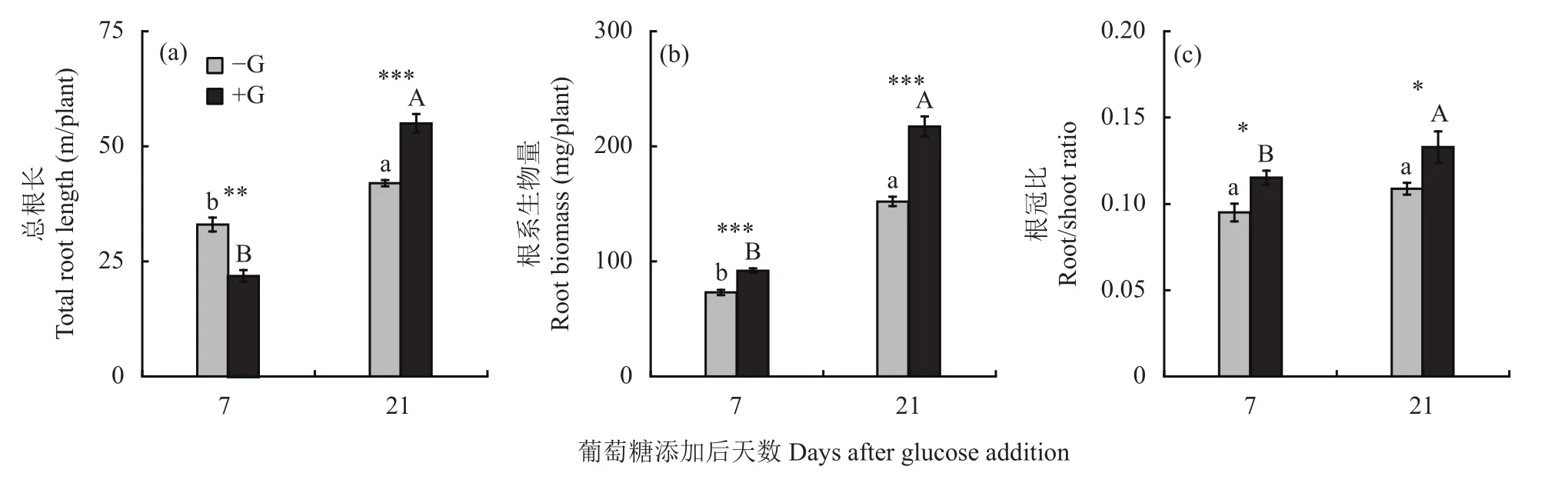
图2 添加葡萄糖对第7天和第21天上海青根系总根长、生物量和根冠比的影响Fig. 2 Total root length, root biomass and root/shoot ratio of Brassica chinensis at 7 and 21 days after glucose addition
如图3所示,添加葡萄糖后上海青根系生长速率、根系平均直径和比根长产生了显著变化,对根系组织密度没有显著影响。与不加葡萄糖处理(−G)相比,添加葡萄糖(+G)后第7天上海青根系生长速率显著降低了36%,根系平均直径增加了27%,比根长降低了46%;第21天,添加葡萄糖促使根系生长速率增加了295% (P< 0.01),根系平均直径、比根长以及根系组织密度无显著变化。

图3 添加葡萄糖第7天和第21天上海青根系生长速率、平均直径、比根长和组织密度Fig. 3 Root growth rate, average root diameter, specific root length and tissue density of Brassica chinensis at 7 and 21 days after glucose addition
如图4所示,添加葡萄糖显著影响了上海青根际酸性磷酸酶、柠檬酸的分泌,对苹果酸的分泌没有影响。添加葡萄糖后的第7天,根际酸性磷酸酶活性降低,根际柠檬酸的分泌与不加葡萄糖处理(−G)相比增加了106%。从第7天到第21天不加葡萄糖处理上海青根际酸性磷酸酶、柠檬酸的分泌增加,而添加葡萄糖处理上海青根际柠檬酸分泌降低。第21天,与不添加葡萄糖的对照处理(−G)相比,添加葡萄糖处理(+G)上海青根际酸性磷酸酶活性降低了52%,根际柠檬酸含量降低了71%。

图4 添加葡萄糖第7天和第21天上海青根际土壤酸性磷酸酶活性以及柠檬酸和苹果酸含量Fig. 4 Acid phosphatase activity, citrate and malate content in Brassica chinensis rhizosphere soil at 7 and 21 days after glucose addition
2.3 添加葡萄糖对土壤微生物量碳、微生物量磷和Olsen-P含量的影响
如图5所示,添加葡萄糖显著影响土壤微生物量碳、微生物量磷和土壤Olsen-P含量。在添加葡萄糖后第7天,添加葡萄糖刺激土壤微生物量碳和微生物量磷含量显著增加,其中微生物量磷含量与不加葡萄糖处理(−G)相比增加了70%,土壤Olsen-P含量下降了35%。从第7天到第21天,对照土壤(−G)微生物量磷含量显著增加而土壤Olsen-P含量降低;添加葡萄糖的土壤(+G),微生物量磷含量降低,土壤Olsen-P含量增加。第21天,添加葡萄糖(+G)的土壤Olsen-P含量与不加葡萄糖处理(−G)相比增加了47%。
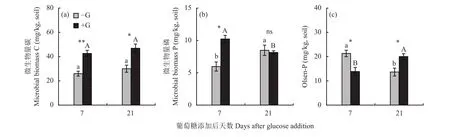
图5 添加葡萄糖第7天和第21天根际土壤中的微生物量碳、微生物量磷和Olsen-P 含量Fig. 5 Microbial biomass C, microbial biomass P and Olsen-P content in Brassica chinensis rhizosphere soil at 7 and 21 days after glucose addition
2.4 上海青根系形态、根际分泌物特征和土壤磷有效性对葡萄糖的响应比
如图6所示,上海青根系生物量对葡萄糖的响应比由添加葡萄糖后第7天的0.12增加至第21天的0.18,总根长从第7天的−0.19增加至第21天的0.14,根系生长速率由第7天的−0.22增加至第21天的0.5,比根长由第7天的−0.30增长至第21天的−0.03。然而,上海青根系平均直径的响应比由0.1降低至0。从第7天至第21天,上海青根际土酸性磷酸酶活性和柠檬酸浓度的响应比显著降低,其中根际酸性磷酸酶活性的响应比由−0.25降至−0.34,柠檬酸浓度的响应比由0.36降至−0.54。土壤微生物量碳的响应比在第7天和第21天无显著变化,微生物量磷的响应比由第7天的0.27降低至第21天的0,Olsen-P的响应比则由第7天的−0.22增加至第21天的0.20。

图6 添加葡萄糖后第7天和第21天上海青地上部生长特征、根系/根际与土壤特征对葡萄糖的响应Fig. 6 Crop growth, root/rhizosphere and soil traits of Brassica chinensis at 7 and 21 days after glucose addition
2.5 微生物介导磷周转过程对植物地上部生长和根系功能属性的影响
如图7所示,冗余分析结果显示了土壤微生物量磷以及土壤Olsen-P对上海青根系形态和根际分泌物的影响。冗余分析RDA1轴和RDA2轴分别解释了61.5%和15.7%的根系形态和根际分泌物变化特征。微生物介导的磷周转对上海青根系平均直径、根系生长速率、比根长、根际柠檬酸和苹果酸分泌有较强的影响。其中,土壤微生物量磷与根系生物量、根冠比、根系直径、柠檬酸的分泌呈正相关;Olsen-P含量则与根系生长速率呈显著正相关。
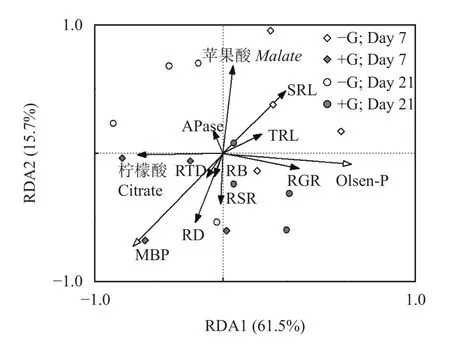
图7 根际土壤微生物量磷和Olsen-P对根系功能属性影响的冗余分析Fig. 7 Redundancy analysis (RDA) of the effects of rhizosphere soil microbial biomass P (MBP) and Olsen-P content on root indexes of Brassica chinensis
3 讨论
与不加葡萄糖处理相比,添加葡萄糖后土壤微生物量磷在第7天增加而第21天降低,土壤Olsen-P则在第7天降低而第21天上升,该结果表明添加葡萄糖启动了微生物介导的磷周转,并驱动了土壤有效磷含量的动态变化(图5~图7)。添加葡萄糖后的第7天(作物生长的第21天),土壤Olsen-P含量显著下降(图5~图6), 这可能是因为葡萄糖为微生物提供足够碳源,刺激微生物的快速增生,微生物为满足自身生长需从土壤中快速吸收大量的磷,导致土壤可利用的有效磷(Olsen-P)含量下降[29],植物根系与微生物之间对土壤磷的竞争加剧[3,30]。以往的研究表明土壤磷有效性的降低会诱导植物根系伸长,比根长增加,根际酸性磷酸酶活性增加,提高对土壤磷的吸收[4,31]。然而,本试验发现添加葡萄糖后第7天上海青根系总根长降低,根系平均直径增加,比根长降低,酸性磷酸酶活性降低(图2~图4),这说明还存在其他环境因素直接或间接影响根系形态和根际分泌物的响应。例如,微生物增殖能够直接改变根系生长调节剂(如生长素)的分泌[32],可能导致根系变粗,伸长速率降低(图2、图3);另外,在添加葡萄糖后的第7天,即作物生长的前21天,作物处于幼苗期,根系较小,竞争能力弱,植物根系为了适应微生物特别是有害微生物的快速增殖,会提高碳向地下根系的分配,增加粗根的生长,增强作物抵御有害微生物侵袭的能力[33–35]。作物通过协调根系形态和根际生理过程响应土壤环境的变化,以获取最大的磷吸收[4]。例如,微生物增生刺激粗根的增生[33–35],粗根的生长伴随着有机酸或磷酸酶分泌的增强(图4b)[4,15],然而,微生物增生尤其是编码碱性磷酸酶phoD基因的磷代谢微生物群落的增生,加剧了微生物与根系对有机磷的竞争[36],这可能会诱导植物分泌大量的有机酸以躲避微生物对有机磷源的竞争[33],进而提高对土壤难溶性无机磷的活化与利用[2,15]。粗根降低了根系的周转速率,有助于根系有效的保持养分,避免由于根系组织脱落造成的磷损失,以维持作物生长发育的需要[37]。综上所述,添加葡萄糖初期,微生物的大量繁殖导致土壤Olsen-P的显著降低,上海青增加根际柠檬酸的分泌,活化并利用土壤难溶性无机磷,促进粗根的生长,降低根系周转造成的磷损失[37],进而提高磷的吸收与利用效率,维持地上部磷吸收和生物量累积(图1、图7)。
微生物磷周转是由微生物增生与死亡驱动的动态过程,从而决定了土壤磷有效性的动态变化以及根系/根际分泌物的动态响应。随着碳的耗竭,微生物因缺乏碳源死亡裂解[19],而根系逐渐变大,竞争能力逐渐增强,同时微生物死亡并向土壤中释放磷,促进土壤有效磷(Olsen-P)含量的增加(图5)[35]。微生物死亡和土壤有效磷(Olsen-P)的提高意味着植物根系生长环境的改善,以及作物根系形态与根际生理过程的响应[38]。例如,从添加葡萄糖后第7天到第21天,添加葡萄糖处理上海青根系生长速率显著高于未添加葡萄糖的处理,总根长增加(图2)。另外,尽管第7天添加葡萄糖处理根系平均直径显著高于未添加葡萄糖的处理,但是在第21天,二者的差异缩小(图3),这表明在第7天到第21天的营养生长期上海青加快了细根的增生,以提高根系对有效磷的吸收[15]。因此,上海青通过降低根际酸性磷酸酶活性和柠檬酸的分泌,将资源供给根系的快速伸长(图2c、图3a)[4,39]。由此可知,添加葡萄糖刺激微生物量磷库的增加,而随着碳的耗竭,磷从死亡微生物转移到土壤,促进土壤有效磷含量的提高,诱导作物根系的快速伸长(图5、图7)。
本研究通过根系碳驱动的微生物磷周转与植物根系/根际特征的共变关系,阐明了微生物协同根系影响蔬菜作物(上海青)磷吸收的互作效应。虽然在本研究中,葡萄糖的添加对植物地上部磷吸收和生物量没有显著的影响,但是大量的研究表明外源有机碳的调控对作物磷吸收和产量有显著的促进效应[40–43]。这可能是由于本试验是在作物生长后的第14天添加的外源葡萄糖,微生物爆发对根系造成的直接伤害影响了碳驱动微生物磷周转协同根系对作物磷吸收作用的发挥[34–35]。另外,添加葡萄糖后第21天便停止观测,此时添加葡萄糖后的土壤有效磷含量仍然比未加葡萄糖的土壤高,土壤中植物可利用的有效磷资源未被充分吸收,微生物磷周转对作物磷吸收的促进效应没有得到充分发挥。为了明确微生物磷周转及其对作物磷吸收的调控效应,我们需要通过进一步的研究来探索强化微生物磷周转调控根系-微生物互作及其对作物磷利用的长期影响。尽管在本试验条件下碳调控对蔬菜作物生长和磷吸收没有显著的影响,然而本研究证明碳驱动的微生物磷周转与作物根系形态及根际分泌物的即时响应共同决定作物磷吸收。由此可见,碳驱动的微生物-根系互作过程是外源碳调控促进作物磷吸收效率的重要机制,通过外源碳调控强化微生物-根系互作增效作用是提高集约化蔬菜种植体系磷利用效率的有效措施。
4 结论
添加葡萄糖显著增加了前期(添加葡萄糖后第7天)根际土壤中的微生物量碳和磷,降低了土壤有效磷含量,作物通过增加柠檬酸的分泌来满足自身对磷的需求。后期(添加葡萄糖后第21天)微生物量磷的降低,促进了根际有效磷含量的增加,诱导根系的快速增生,改善作物的磷素营养。因此,碳通过驱动土壤微生物,从土壤磷有效性和作物根系磷吸收策略两方面影响了作物的磷素营养。
