外源NO对镉胁迫下生菜种子萌发、幼苗生长及抗氧化特性的影响
2021-12-03熊明国
熊明国
(商丘职业技术学院,河南商丘 476100)
随着工业“三废”的排放、矿产资源的开采、污水灌溉以及各种含重金属有机肥料的长期使用,我国土壤重金属污染问题日益严重,其中,镉(Cd)污染尤为严重。Cd的毒性较其他重金属更强,具有致癌性。土壤中过量的Cd被植物吸收,并在可食部位大量积累,通过食物链进入人体,从而危害人体健康。Cd 污染相关问题是人类重要的研究课题。目前,很多学者都在研究Cd、铬(Cr)、铅(Pb)等重金属胁迫对植物的影响及其相应的解决方法[1‑8]。一氧化氮(NO)是植物体内重要的活性分子,对植物的萌发、生长发育、气孔运动等生理活动有重要调节作用。NO 能激活植株的过敏性反应,促使植物获得抗性,还可诱导植株对高温、缺水和盐胁迫等非生物胁迫的抗性,更可以调节体内的激素水平,调节植物生长发育。研究发现,外源NO 对植物重金属(Cr、Cd、Pb等)胁迫具有一定的缓解作用,这些植物主要集中于大豆、烟草、油菜、玉米、水稻、绿豆、黄瓜、蒙山茶、三叶鬼针草等[9‑17]。生菜是一种常见的绿叶蔬菜,易于栽植,对环境因子的胁迫也具有较强的反应,相对于其他蔬菜(番茄、黄瓜、茄子、辣椒、菠菜等)更容易吸收和积累Cd。目前,在生菜上采用的缓解重金属毒害的物质主要有农业废弃物、石灰、生物炭、金属氧化物等[18],关于外源NO 对生菜重金属胁迫的缓解作用研究很少见[19]。徐林林[19]采用盆栽,将外源NO的供体硝普钠(SNP)通过不同途径加入土壤,研究外源NO 对Cd 胁迫下花生与生菜生长的缓解效应,结果认为,叶面喷施500 μmol/L SNP 效果最好。目前,尚未见相关水培研究结果。为此,设置不同浓度SNP,研究水培条件下外源NO对Cd胁迫下生菜种子萌发、幼苗生长及抗氧化特性的影响,为进一步开展外源物质对重金属毒害的缓解效应研究提供参考。
1 材料和方法
1.1 试验材料
供试生菜为海南省海口市育新种子商行生产的“意大利生菜”(抗热、耐抽薹)。SNP 和氯化镉(CdCl2·2.5H2O)均购自天津市欧博凯化产品销售有限公司。营养液采用的是Hoagland’s(霍格兰氏)营养液。
1.2 试验设计
2019 年3—5 月在商丘职业技术学院园艺专业实训基地进行试验。挑选饱满且大小均一的生菜种子放在铺有2层滤纸的培养皿(直径90 cm)中,然后在培养皿中分别添加单一营养液(CK)、含100 μmol/L Cd2+的营养液、含100 μmol/L Cd2+和100 μmol/L SNP的营养液、含100 μmol/L Cd2+和150 μmol/L SNP的营养液、含100 μmol/L Cd2+和200 μmol/L SNP 的营养液、含100 μmol/LCd2+和250 μmol/L SNP 的营养液以及含100、150、200、250 μmol/L SNP 的营养液,共10个处理,具体处理及编号见表1,100 μmol/L Cd2+胁迫是在前期试验基础上根据生菜相对生长速度筛选的(数据未发表)。每皿添加相应处理液3 mL。每处理100 粒种子,重复3 次,完全随机区组排列。将培养皿置25 ℃下催芽,每天定时观察发芽数,用称质量法补充处理液,5 d 发芽结束后将其种植到72 孔穴盘中,每穴1 株,每处理9穴,重复3 次。移植当天用相应处理液浇透培养基质(m蛭石∶m珍珠岩=3∶1),保持其湿度。每天定时查看维护,10 d 后采用Hoagland’s 营养液浇灌幼苗,14 d 后取样用于指标测定。
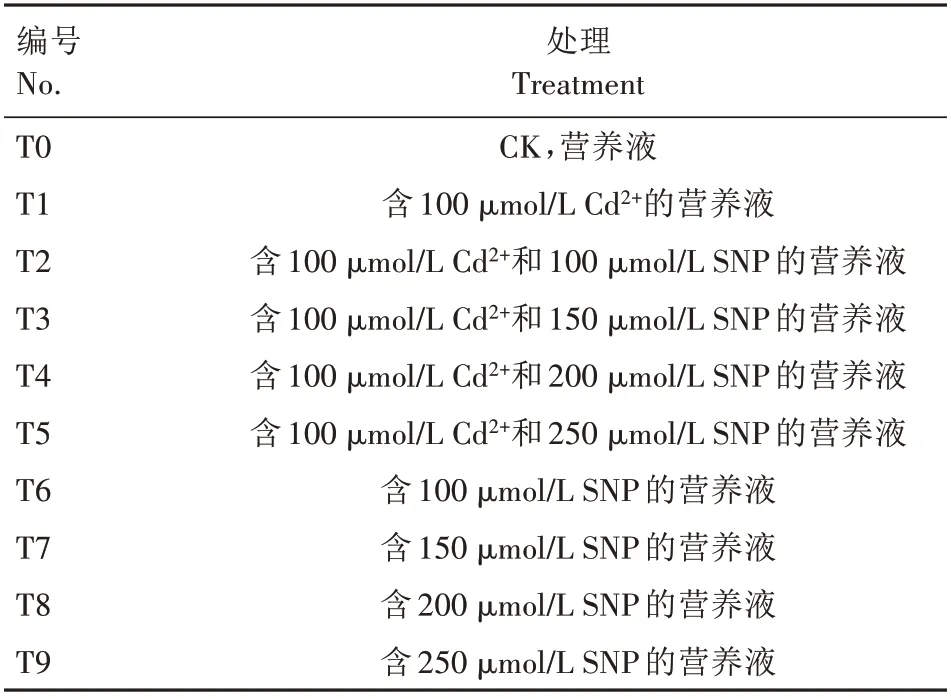
表1 处理及编号Tab.1 Treatment and number
1.3 测定项目及方法
1.3.1 发芽势、发芽率 观察3 d 内和5 d 内发芽种子数,并计算发芽势、发芽率。其中,发芽势=(3 d内发芽种子数/被测种子数)×100%,发芽率=(5 d 内发芽种子数/被测种子数)×100%。
1.3.2 叶绿素含量 叶绿素含量采用分光光度法测定[20]。
1.3.3 抗氧化特性 过氧化物酶(POD)活性的测定采用愈创木酚法,超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑法,过氧化氢酶(CAT)活性的测定采用高锰酸钾滴定法,丙二醛(MDA)含量的测定采用硫代巴比妥酸法[20]。
1.4 数据处理
数据采用Excel 2016 进行处理和作图,使用软件SPSS 22.0进行差异显著性分析。
2 结果与分析
2.1 外源NO对Cd胁迫下生菜种子萌发的影响
2.1.1 发芽率 从图1 可看出,生菜种子遭遇Cd 胁迫后发芽率较CK 显著降低,说明Cd 胁迫抑制生菜种子发芽;在Cd 胁迫下添加适宜浓度的外源NO,T2、T5 处理生菜种子发芽率有所提高,但差异不显著。在营养液中添加不同浓度的SNP,随着SNP 浓度的增加,生菜种子发芽率先升高后降低,T8 处理最高,为96.6%,与CK无显著差异。
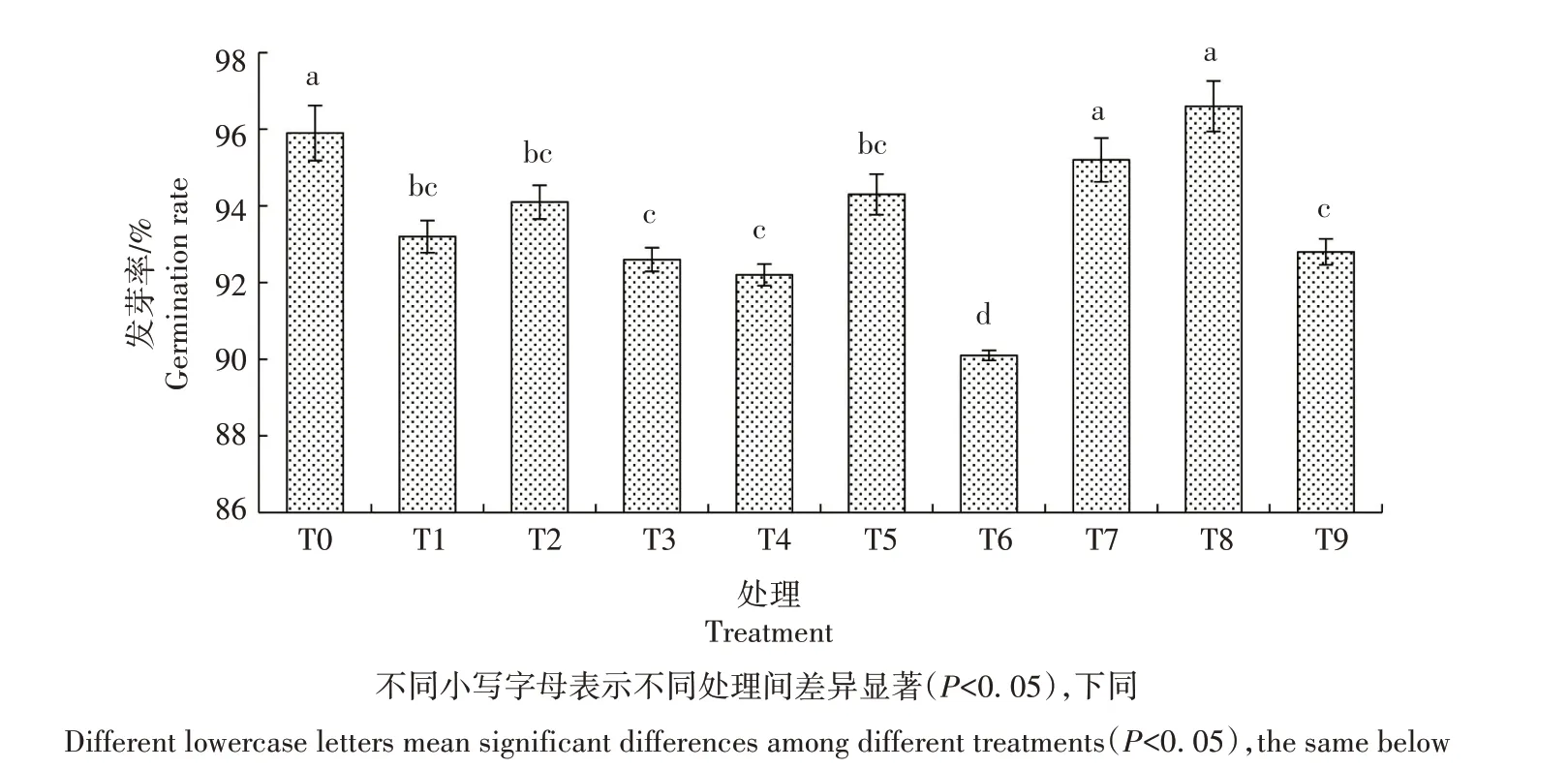
图1 外源NO对Cd胁迫下生菜种子发芽率的影响Fig.1 Effect of exogenous NO on seed germination rate of lettuce under cadmium stress
2.1.2 发芽势 由图2 可知,生菜种子遭遇Cd 胁迫后发芽势较CK 降低,但差异不显著;在Cd 胁迫下添加不同浓度的NO,其发芽势与CK、T1 处理均无显著差异。在营养液中添加不同浓度的SNP,随着SNP 浓度的增加,生菜发芽势总体升高,T8 处理最高,但与CK 无显著差异。综上可知,在Cd 胁迫下生菜种子的发芽势并未明显受到影响。
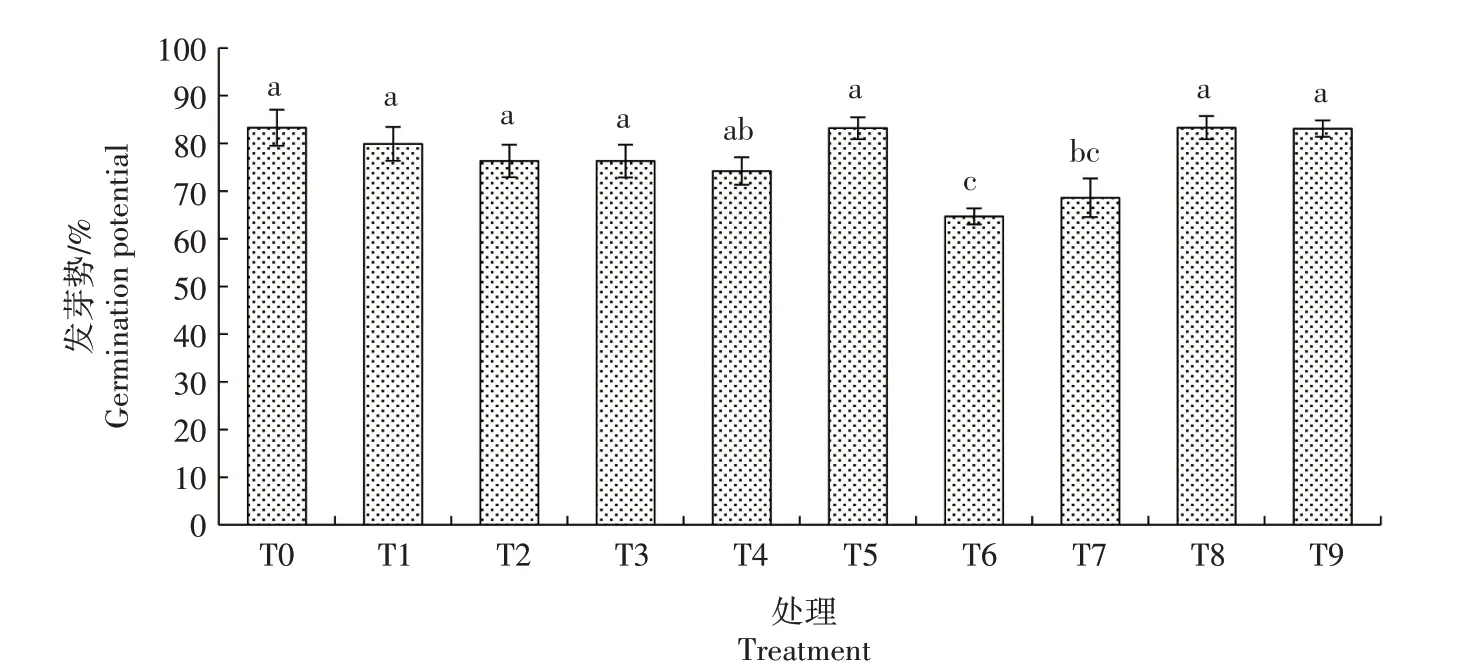
图2 外源NO对Cd胁迫下生菜种子发芽势的影响Fig.2 Effects of exogenous NO on seed germination potential of lettuce under cadmium stress
2.2 外源NO对Cd胁迫下生菜幼苗生长的影响
2.2.1 叶绿素含量 由图3 可知,生菜幼苗遭遇Cd胁迫后叶片叶绿素较CK 显著降低,在Cd 胁迫下添加不同浓度的外源NO,叶绿素含量显著提高,且随着NO 浓度的升高而升高,以T5 处理最高,T2、T3、T4、T5 处理分别较T1 处理提高131.00%、145.01%、151.38%、155.63%,但这些处理仍显著低于CK。仅添加SNP 的T6、T7、T8、T9 处理叶绿素含量随着SNP浓度增加而增加,以T9处理最高,与CK 无显著差异,其他处理均显著低于CK。因此,Cd 胁迫会抑制生菜叶叶绿素含量,且外源NO 可在一定程度上缓解此影响。
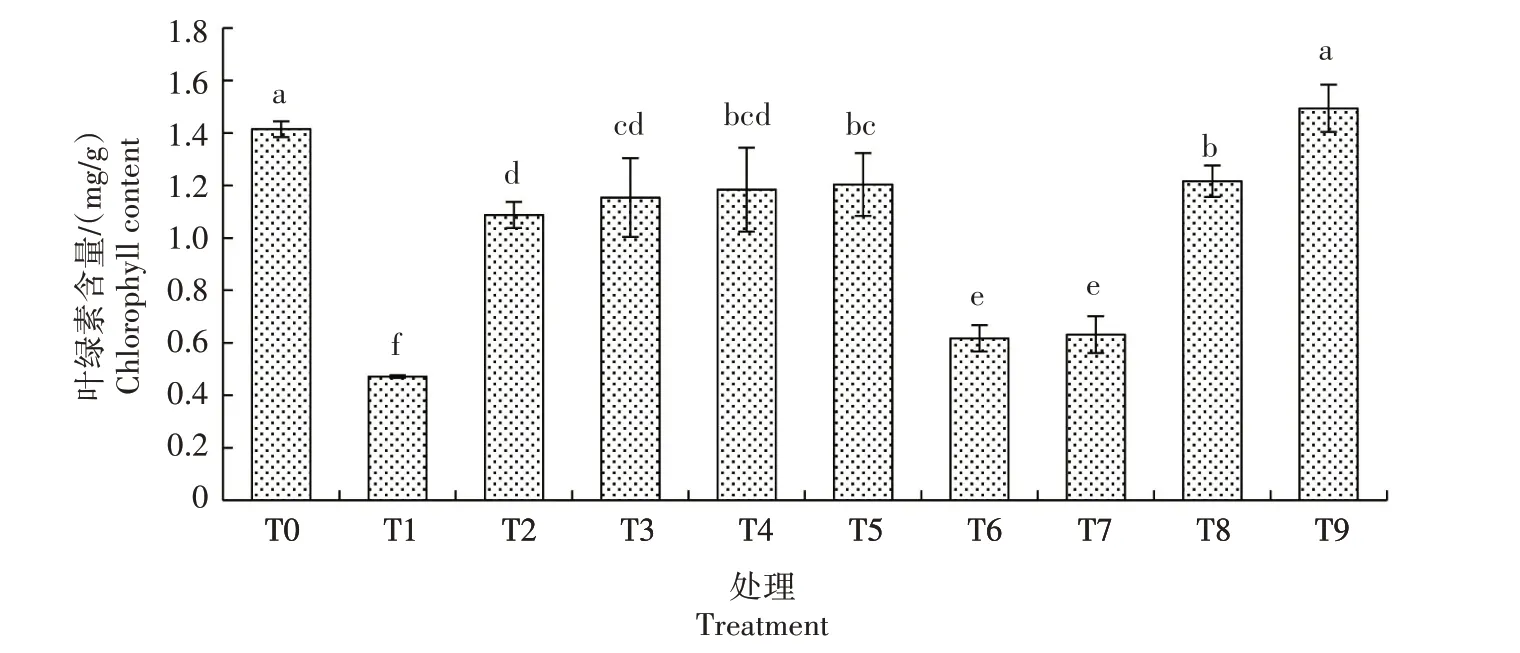
图3 外源NO对Cd胁迫下生菜叶绿素含量的影响Fig.3 Effects of exogenous NO on chlorophyll content of lettuce under cadmium stress
2.2.2 地上部、地下部长度 从图4可看出,生菜幼苗遭遇Cd 胁迫后地上部、地下部长度均较CK 显著降低;在Cd胁迫下添加不同浓度的外源NO,总体上地上部、地下部长度均显著提高,地上部长度以T5处理最高,地下部长度以T4处理最高。仅添加SNP的T6、T7、T8、T9 处理,地上部长度随着SNP 浓度的增加而降低,地下部长度随着SNP 浓度的增加先增加后降低,除T9处理较CK降低外,其他处理总体上均高于CK。由此可知,Cd 胁迫会抑制生菜的生长,添加外源NO则会适当缓解这种抑制作用。
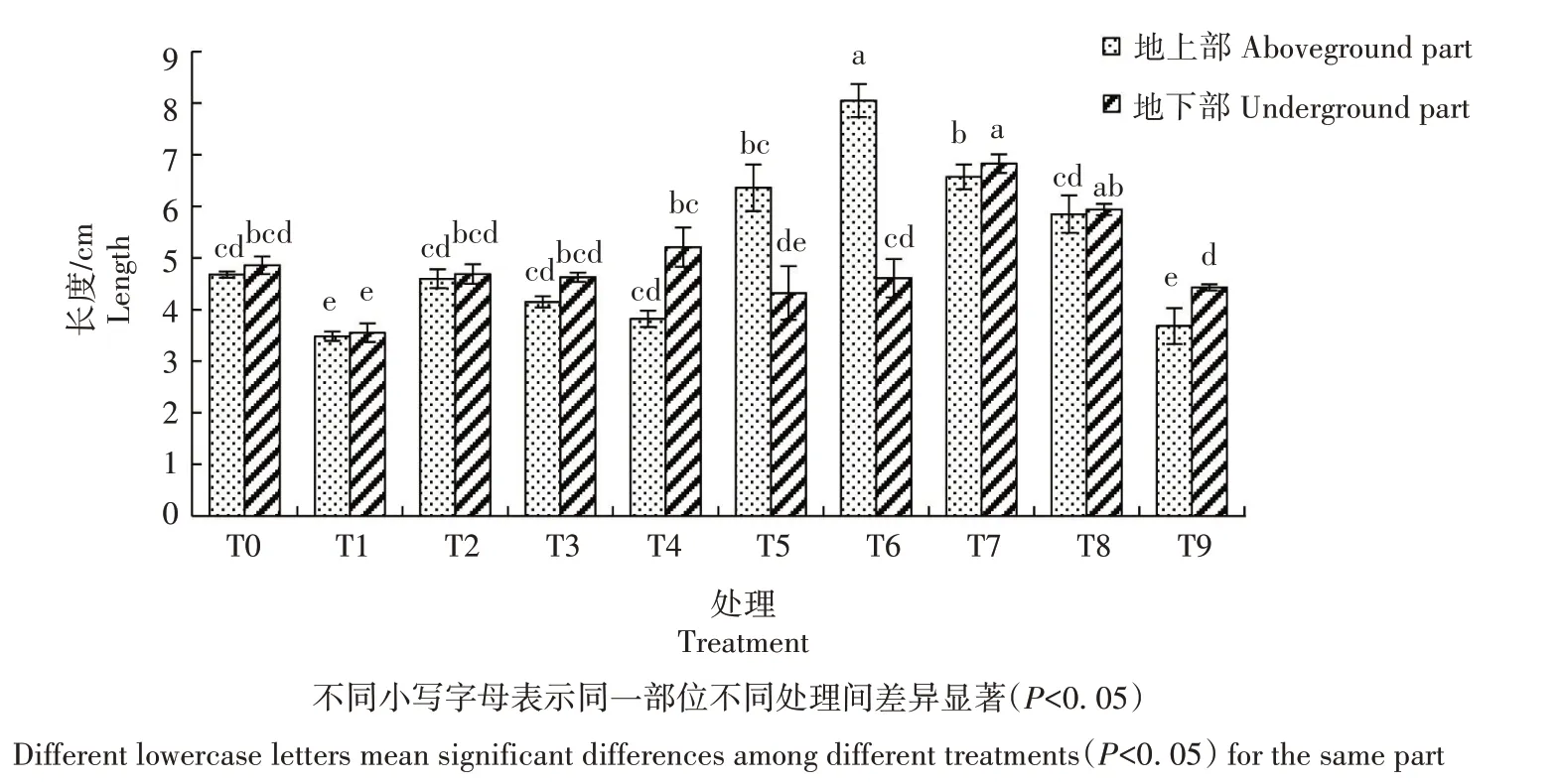
图4 外源NO对Cd胁迫下生菜幼苗生长的影响Fig.4 Effects of exogenous NO on growth of lettuce seedlings under cadmium stress
2.3 外源NO对Cd胁迫下生菜幼苗抗氧化特性的影响
2.3.1 MDA含量 从图5可知,生菜幼苗遭遇Cd胁迫后MDA含量较CK 显著升高;在Cd胁迫下添加不同浓度的外源NO,MDA 含量均显著降低,且均显著低于CK,以T5 处理最低。仅添加SNP 的T6、T7、T8、T9 处理MDA 含量普遍较低,显著低于CK 和T1处理。由此可知,Cd 胁迫会造成生菜幼苗MDA 含量增加,而外源NO则会抑制MDA的产生。
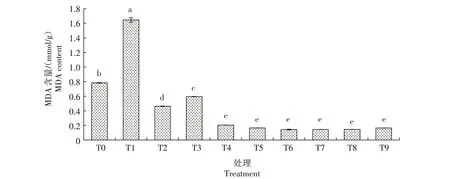
图5 外源NO对Cd胁迫下生菜幼苗MDA含量的影响Fig.5 Effects of exogenous NO on the MDA content of lettuce seedlings under cadmium stress
2.3.2 抗氧化酶活性
2.3.2.1 POD 活性 由图6 可看出,生菜幼苗遭遇Cd胁迫后POD活性较CK显著降低;在Cd胁迫下添加不同浓度的外源NO,POD 活性显著升高,且总体随着SNP 浓度的升高而升高,以T5 处理最高,较T1处理显著提高85.00%,与CK 无显著差异,其他处理均显著低于CK。仅添加SNP 的处理中,T6、T8、T9处理POD 活性与CK 无显著差异,T7 处理显著低于CK,但均显著高于T1处理。由此可知,Cd胁迫下生菜幼苗的POD 活性会受到抑制,而一定量的外源NO 则会在一定程度上缓解Cd 胁迫对POD 活性的抑制作用。
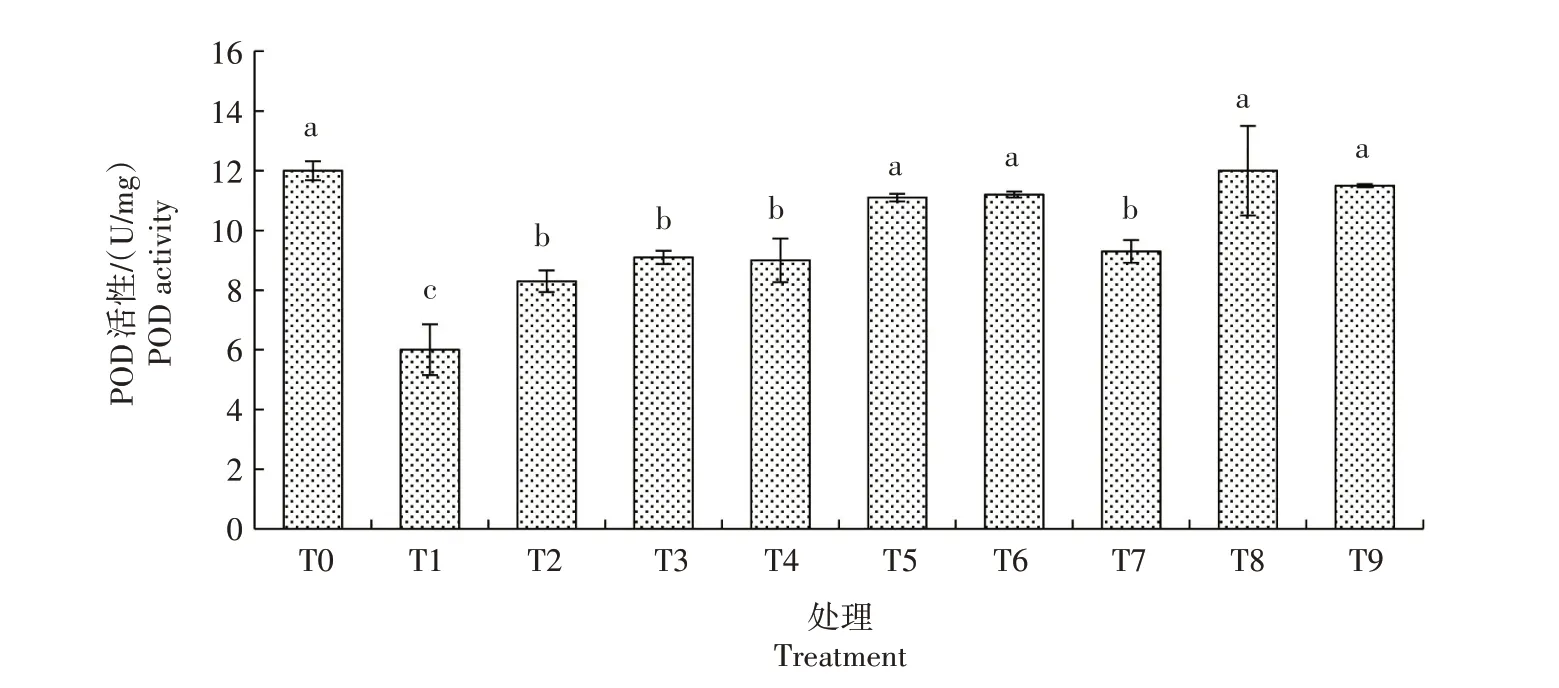
图6 外源NO对Cd胁迫下生菜幼苗POD活性的影响Fig.6 Effects of exogenous NO on the POD activity of lettuce seedlings under cadmium stress
2.3.2.2 CAT 活性 由图7 可知,生菜幼苗遭遇Cd胁迫后CAT 活性较CK 显著降低;在Cd 胁迫下添加不同浓度的外源NO,POD 活性显著升高,且总体随着SNP 浓度的升高而升高,以T5 处理最高,较T1 处理显著提高81.20%,与CK 无显著差异,其他处理均低于CK。仅添加SNP 的处理间CAT 活性差异不显著,均显著高于T1 处理,稍低于CK。由此可知,生菜幼苗在Cd 胁迫下的CAT 活性会受到一定程度的抑制,而添加适当的外源NO 则可一定程度地缓解这种抑制作用。
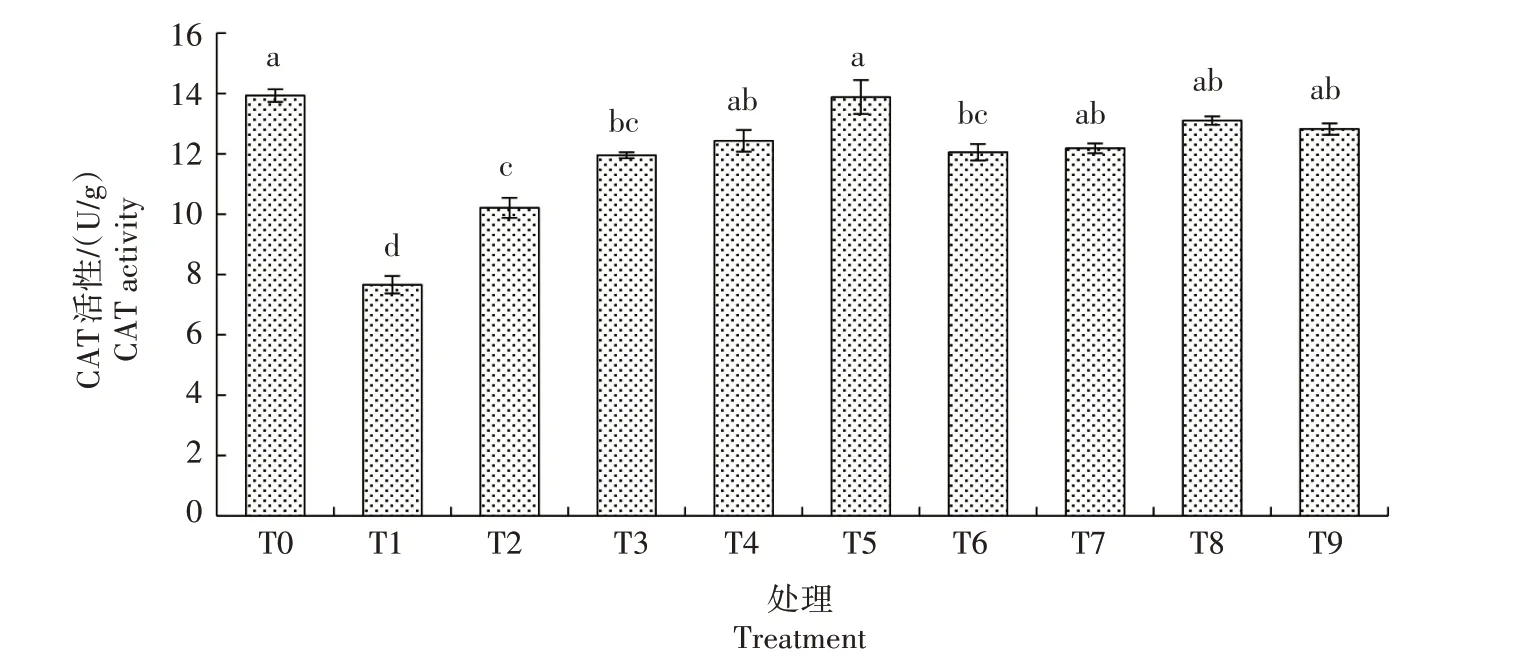
图7 外源NO对Cd胁迫下生菜幼苗CAT活性的影响Fig.7 Effects of exogenous NO on the CAT activity of lettuce seedlings under cadmium stress
2.3.2.3 SOD 活性 由图8 可知,生菜幼苗遭遇Cd胁迫后SOD 活性较CK 显著降低;在Cd 胁迫下添加不同浓度的外源NO,POD 活性显著升高,且随着SNP 浓度的升高而升高,以T5 处理最高,较T1 处理显著提高63.27%,所有处理均低于CK。仅添加SNP的处理中,T6、T8、T9处理间POD活性差异不显著,均显著高于CK;T7 处理与CK 无显著差异。由此推断,Cd 胁迫对生菜幼苗SOD 活性有抑制作用,而外源NO则可以适当缓解Cd胁迫的毒害效应。
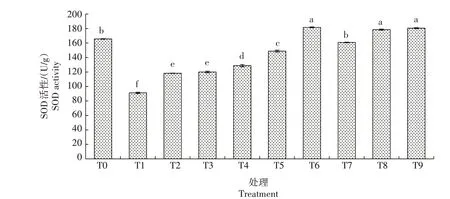
图8 外源NO对Cd胁迫下生菜幼苗SOD活性的影响Fig.8 Effects of exogenous NO on the SOD Activity of lettuce seedlings under cadmium stress
3 结论与讨论
本研究结果表明,外源NO 对Cd 胁迫下生菜种子发芽率、发芽势均无显著影响,但总体上可显著提高生菜幼苗叶绿素含量、地上部长度、地下部长度及POD、CAT、SOD 活性,显著降低MDA 含量,以250 μmol/L SNP处理效果最佳。
近年来,关于重金属胁迫对植物生长的影响及缓解措施研究较多[1‑3,5‑7,9‑23],大多学者[9‑17]均认为,外源NO 对植物重金属胁迫存在剂量效应,低浓度有缓解效应,高浓度有毒害效应。本研究得出类似结论,Cd 胁迫会严重抑制生菜的生长,适宜浓度外源NO 可以缓解Cd 对生菜的毒害。徐林林[19]通过将SNP直接施入土壤、包进胶囊颗粒和纸袋、制成缓释颗粒肥、制成溶液叶面喷施5 种方式向Cd 污染土壤中生长的生菜供应NO,5 种方式对缓解生菜Cd 毒害的效应各异,效果最差的是SNP 直接施入土壤,最佳方式则是叶面喷施,SNP 最佳喷施浓度为500 μmol/L。本研究采用水培方式,设置100、150、200、250 μmol/L 4 个浓度,发现250 μmol/L SNP 效果最好。可能是因为水培方式,生菜根系漂浮水中,更易吸收SNP 溶液,所以SNP 浓度较徐林林[19]认为的低。
