Ⅱ型细胞色素P450酶氧化β-香树脂醇的选择性调控研究
2021-11-29孙文涛张昕哲万盛通王茹雯李春
孙文涛,张昕哲,万盛通,王茹雯,李春,
(1 清华大学化工系,北京 100084;2 北京理工大学化学与化工学院,北京 100101)
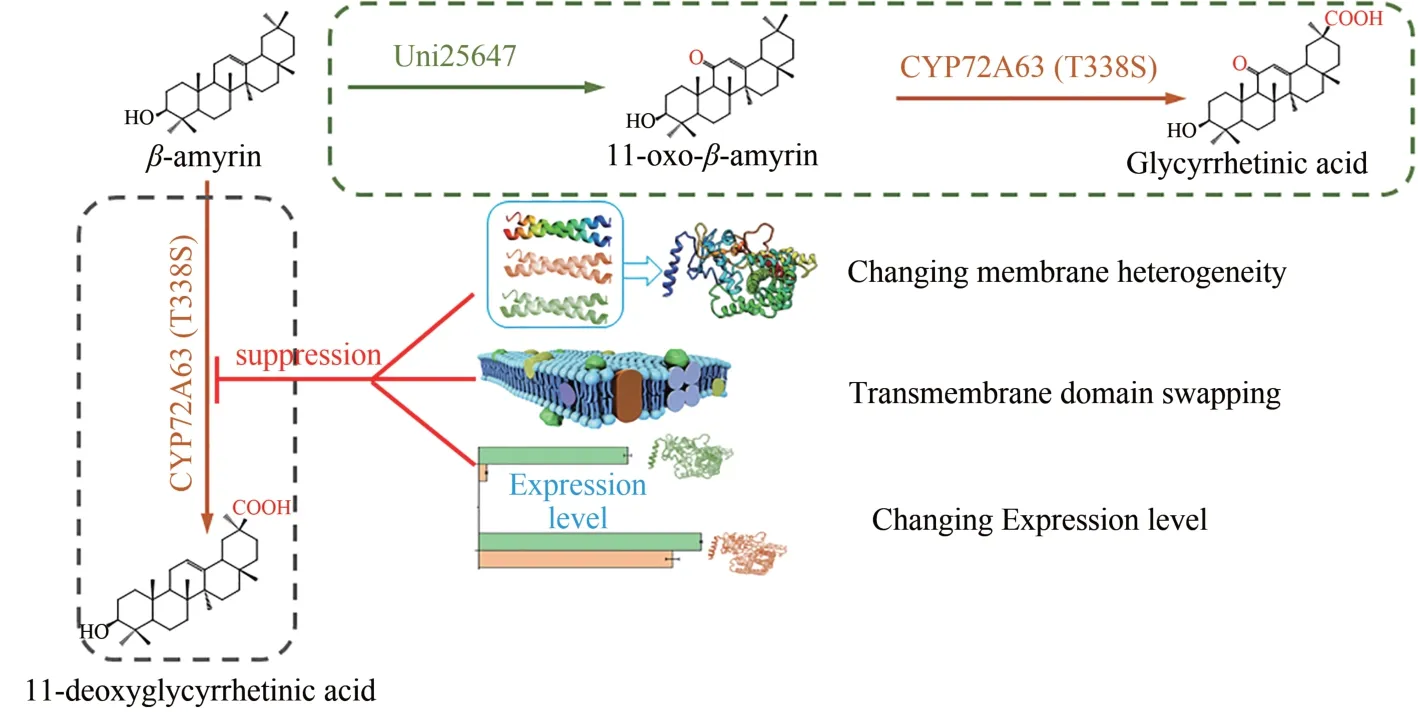
细胞色素P450 酶(P450)可催化碳氢键氧化、脱甲基、脱饱和等二十余类反应,在植物次级代谢产物的生物合成中扮演着重要的角色,是萜烯化合物、生物碱及黄酮等多种天然产物合成中的关键修饰酶[1-2],在高价值天然产物的异源合成中被广泛研究和应用[3-7],比如重要保肝护肝药物甘草次酸的合成过程中,CYP88D6 与CYP72A154 对骨架分子β-香树脂醇C-11 与C-30 进行连续氧化最终将其转化成甘草次酸[8-9]。
P450 是一种含血红素蛋白,利用分子氧及NAD(P)H 对底物进行加氧反应,根据P450 酶从NAD(P)H 获取电子的方式不同可将其分为4 种类型:Ⅰ型P450酶的氧化还原伴侣需要一个包含FAD的还原酶以及一个铁氧还蛋白构成;Ⅱ型P450酶的氧化还原伴侣为一个同时包含FMN和FAD结构域的单一蛋白又称P450还原酶(CPR);Ⅲ型P450酶为自给自足的蛋白,其实质为P450酶与氧化还原伴侣形成的多结构域蛋白;Ⅳ型P450 酶可以直接从NAD(P)H中获取电子,不需要额外的氧化还原伴侣蛋白或结构域。其中细菌等原核生物来源的P450酶多为Ⅰ型和Ⅲ型,且通常为游离蛋白,动物、植物等来源的P450酶多为Ⅱ型,且通常为膜蛋白,具有N端跨膜结构域,比如甘草来源的CYP88D6 与CYP72A154[10]。
由于Ⅱ型P450 酶难以纯化,因此目前对于其反应特性调控的研究较少。基于同源建模等方法,张学礼课题组通过对来自于犁头霉菌的CYP5311B2底物通道进行改造提高其催化活性,使工程菌的氢化可的松产量提高了3倍[11]。Schele等通过同源建模和定点突变对铁锈醇合酶CYP76AH1 以及11-羟基铁锈醇合酶 CYP76AH4、 CYP76AH22、CYP76AH23、 CYP76AH24 进 行 改 造 发 现 301、303以及478的氨基酸是决定11-羟基铁锈醇合酶选择性氧化铁锈醇的关键氨基酸位点[12]。当前对于P450 催化特性调控的相关研究主要集中于可溶性的P450酶,如来自巨大芽孢杆菌的P450BM3[13-15]。研究人员通过对其活性口袋进行重塑实现了其区域和立体选择性的调控,可控合成了对苯二酚、2β-羟基睾丸酮等化合物,并进一步拓宽了P450 酶催化反应的范畴,成功实现了S—N 键、C—Si键、C—B 键的形成以及芳基烯烃的氮杂环丙烷化、卡宾向炔烃的转移反应等一系列新型反应[16-19]。
为实现Ⅱ型P450 酶催化特性的调控,解决甘草次酸合成关键酶,来源于蒺藜状苜蓿的CYP72A63(AB558146.1)催化的底物选择性、区域选择性以及氧化进程不可控的问题,提高甘草次酸合成特异性与合成效率。课题组前期通过计算机辅助的理性设计与酿酒酵母验证平台,重塑了CYP72A63 的催化口袋,获得高活性突变体CYP72A63(T338S)可对 11-氧-β-香树脂醇 C-30 特异性的连续氧化,实现甘草次酸的可控合成。然而仍然无法解决其底物选择性差的问题,CYP72A 63(T338S)可同时对β-香树脂醇以及 11-氧-β-香树脂醇的C-30 进行氧化,导致副产物11-脱氧甘草次酸的合成。在酶改造过程中我们发现,除了活性口袋重塑外,Ⅱ型P450 酶与其氧化还原伴侣CPR之间的相互作用可改变其催化活性以及选择性,影响工程菌甘草次酸产量及产物谱[20]。这种现象同样出现在Ⅰ型P450 酶中,李盛英等通过重构P450酶MycG与氧化还原伴侣的组合,重建了其催化功能,实现了4种新产物的合成[19],这说明P450酶催化系统组成成分之间的相互作用会对P450的催化特性产生显著影响。由于Ⅱ型P450酶多定位于内质网,除CPR外,P450、CPR与膜之间的相互作用都可能对P450 酶催化特性产生影响[21]。为实现CYP72A63(T338S)底物选择性的调控,本文对于跨膜域、膜组分代谢等因素对P450酶催化特性调控的影响进行了研究,以期获得能够调控其底物选择性的新方法,以CYP72A63(T338S)为对象,开发Ⅱ型细胞色素P450酶催化选择性调控的新策略。
1 材料与方法
1.1 菌株与Ⅱ型P450酶突变体
1.2 基因克隆与表达质粒的构建
本研究中利用诺唯赞Phanta HS Super-Fidelity DNA Polymerase 进 行 PCR 扩 增 , PCR 产 物 用Thermo Scientific GeneJET 胶回收试剂盒进行纯化。通过吉布森组装将启动子、基因、终止子、同源臂等按照需求进行多片段组装(成分如表1所示),构建含有目标表达盒的质粒,随后利用化学转化法转化至E.coliDH5α 感受态并涂布到含有100 mg/L 氨苄青霉素的LB固体培养基,置于37 ℃静置培养过夜。通过PCR验证,获得条带正确的质粒由擎科生物技术有限公司进行测序验证后备用。

表1 本研究所涉及的菌种信息Tab.1 Strains used in this study
(1)5×ⅠSO缓冲液配制(表2)
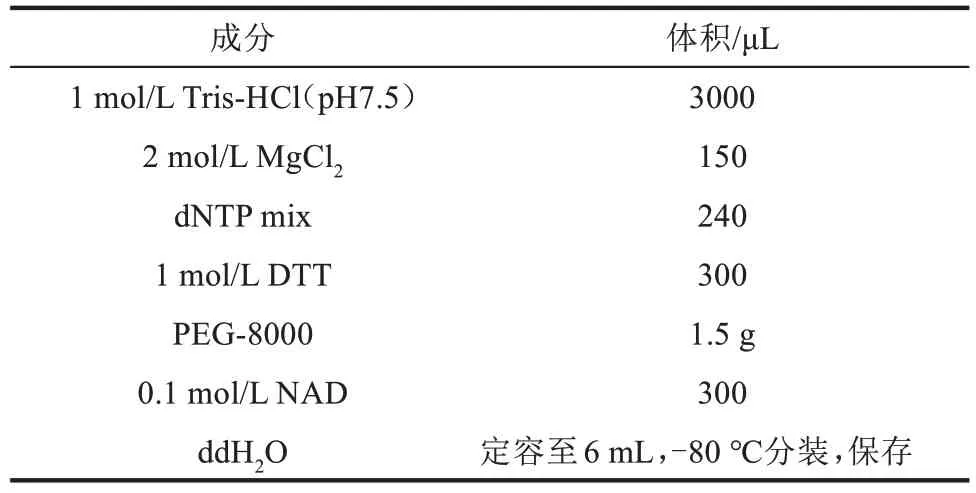
表2 5×ⅠSO缓冲液组分Tab.2 Components of the 5×ⅠSO buffer
(2)Gibson反应液配制(表3)

表3 Gibson反应液组分Tab.3 Components of Gibson reaction solution
1.3 蛋白跨膜域预测
使用跨膜域预测软件TMHMM对蛋白质跨膜域进行预测(http://www.cbs.dtu.dk/services/TMHMM/)[22]。
1.4 酵母转化与培养
酵母转化采用醋酸锂转化法,将新鲜酵母宿主细胞接入2 mL YPD 培养基中,在30 ℃下200 r/min 过夜培养后,以10%的接种量接入新鲜YPD 培养基中继续培养4~5 h,至细胞浓度大约为2×107cells/mL。吸取适量菌液到无菌的1.5 mL离心管中,5000 r/min 离心2 min 后水洗一次并收集细胞。用1 mL 浓度100 mmol/L 的醋酸锂溶液将细胞重悬并静置5 min,随后4000 r/min 离心3 min弃上清。依次在菌体中加入适量DNA 片段、10 µL ssDNA、240 µL 500 g/L PEG3350、36 µL 1 mol/L 醋酸锂,加水至总体积360 µL。涡旋振荡充分混匀,依次30 ℃保温30 min、42 ℃水浴20~30 min,4000 r/min 离心3 min 收集沉淀随后用1 mL 无菌水清洗菌体。取适量菌液涂平板,30 ℃培养2~4天,PCR验证正确后进行发酵验证[23]。
挑取少量PCR 验证正确的菌体接入适量新鲜YPD培养基中培养18~24 h制备种子液,以10%接种量接入新鲜YPD 培养基中,30 ℃,200 r/min,培养5天后提取发酵产物。
1.5 基因表达水平检测
将工程酿酒酵母接在YPD 培养基中培养后提取总RNA 并反转录,以反转录产物为模板利用FastStart Essential DNA Green Master 试剂盒(Roche)通过实时荧光定量PCR 对基因转录进行相对定量。管家基因ACT1为内参基因。
1.6 酵母RNA-Seq
工程酵母接在YPD 培养基中30 ℃培养30 h后,5000 r/min 离心2 min 收集菌体,并用无菌水洗涤菌体3 次后转入-80 ℃短暂保存,随后由北京博云华康基因科技有限公司完成RNA-seq。
1.7 工程酵母代谢产物的样品处理
酵母发酵结束后,取60 mL 菌液用6 mol/L 盐酸调节pH 至3 以下,对其进行超高压破碎。取2 mL 超高压破碎的匀浆,加入等体积乙酸乙酯,涡旋振荡10 min。将匀浆12 000 r/min离心10 min,吸取上清并转移至液相小瓶。在真空干燥仪中60 ℃干燥至乙酸乙酯完全挥发。加入100µL吡啶与100µLN,O-双(三甲基硅基)三氟代乙酰胺,混匀后80 ℃反应30 min。冷却至室温后,将反应液转入200µL套管后置于液相小瓶中,准备上机检测。
1.8 GC-MS产物鉴定
使用岛津GCMS-QP2010 Ultra,配备低流失色谱柱SH-Rxi-5Sil MS(30 m×0.25 mm×0.25 μm,岛津,日本)。进样口温度250 ℃,分流比5∶1,进样1 μL,载气氦气的流速为1.2 mL/min。升温程序:80 ℃保持 1 min,以 20 ℃/min 升至 290 ℃,保持38 min;色谱质谱接口温度为290 ℃。质谱扫描范围为m/z50~700。使用外标法对其进行定性定量[24-25]。
2 结果与讨论
2.1 跨膜域对CYP72A63(T338S)催化特性的影响
Ⅱ型细胞色素P450 酶的跨膜结构对于酶的催化活性具有重要影响[26-27]。在大肠杆菌中表达时,增溶修饰(N 端融合增溶序列,截除跨膜域等)可以提高Ⅱ型P450 酶的溶解性,但是酶与膜相互作用的破坏会对其活性产生显著的抑制作用,CYP11B1 的N 端突变甚至会对其选择性产生影响[26-27]。为进一步探究在酵母体内表达时跨膜域对CYP72A63(T338S)催化特性的影响,本研究将酵母内源细胞色素P450 酶ERG11(YHR007C)、ERG5(YMR015C)与其他内质网定位蛋白ERG1(YGR175C)、ERG2(YMR202W)、NTE1(YML059C)的 N 端跨膜域,以及 GPⅠ8(YDR331W)、KAR1(YNL188W)的C 端跨膜域,直接融合在突变体CYP72A63(T338S)的 N 端以对其 跨 膜域进行改造。
利用TMHMM 对改造后的嵌合蛋白跨膜域结构进行预测,结果如图1 所示。其中红色柱状区域出现的次数代表跨膜螺旋的数量,横坐标代表氨基酸位点,融合ERG5N(ERG5 的N 端跨膜域)、GPⅠ8C(GPⅠ8 的 C 端跨膜域)后仍为单跨膜域蛋白,跨膜域由29 个氨基酸分别延长为69 个和43 个氨基 酸 ;融 合 ERG2N(ERG2 的 N 端 跨 膜 域 )、KAR1C(KAR1 的C 端跨膜域)后,由单跨膜域改变为双跨膜域蛋白;融合ERG11N(ERG11 的N 端跨膜域)与NTE1N(NTE1 的N 端跨膜域)后则变为三跨膜域蛋白,这说明内源内质网蛋白跨膜域的融合显著改变了CYP72A63(T338S)的跨膜结构(图1)。

图1 跨膜域改造后CYP72A63(T338S)的TMHMM预测结果Fig.1 TMHMM prediction for the transmembrane-domain-modified CYP72A63(T338S)
为确定跨膜域改造对CYP72A63(T338S)催化特性的影响,所获得的嵌合蛋白突变体均在酿酒酵母底盘GA0-1 体内进行功能验证。结果表明,跨膜域的改造对CYP72A63(T338S)的催化特 性 影 响 显 著( 图 2)。 ERG5N、 KAR1C、ERG2N 对其活性有显著的抑制作用,KAR1C、ERG2N 直接导致其失活,这也说明这些双跨膜域的结构对该酶活性有显著的抑制作用,其可能原因为改造后的跨膜域无法在内质网上正确定位与折叠从而导致了该酶的失活,在改造过程中应避免这种结构的形成。但是融合NTE1N 后降低了CYP72A63(T338S)对底物β-香树脂醇选择性,显著抑制了其氧化β-香树脂醇的能力,降低甘草次酸合成过程中副产物11-脱氧甘草次酸的合成。而 GPⅠ8C 进一步提高了 CYP72A63(T338S)对底物β-香树脂醇的选择性,促进了甘草次酸合成过程中副产物11-脱氧甘草次酸的合成(图2)。这一结果说明,跨膜域的改造对于细胞色素P450酶的催化特性具有显著的影响,通过对跨膜区域进行改造是一种调控Ⅱ型P450 酶底物选择性的有效策略。

图2 不同跨膜结构对CYP72A63(T338S)底物选择性的影响Fig.2 Ⅰnfluence of the transmembrane structures on the substrate selectivity of CYP72A63(T338S)
2.2 细胞膜组分对CYP72A63(T338S)底物选择性的影响
脂质是生物膜的主要组成成分,细胞中脂质组分的调整对于维持膜的结构稳定性具有重要作用[28],膜结构为膜蛋白发挥功能的支架,也会对膜结合的P450 酶与CPR 之间的相互作用产生影响,从而影响P450 的催化活性[29]。研究表明,在蛋白与蛋白以及蛋白与脂质之间的相互作用下,内质网上能够形成富含甾醇以及鞘脂的脂质微结构域,这些微结构域是P450 以及CPR 在内质网上定位的重要场所,鞘脂分子结构以及种类的改变可能会重塑这些微结构域,影响定位其中的蛋白质与蛋白质之间的相互作用,以及蛋白质与脂质之间的相互作用,进而影响P450 酶的催化特性[30-31]。
在前期研究中我们发现,相同的P450 酶在不同的宿主中表达时,其活性差异较大。以Sgib[32]以及与SynV[33]为宿主,利用 CYP72A63(T338S)在胞内重构甘草次酸的合成途径时,在相同条件下,底物11-氧-β-香树脂醇有大量剩余时,其甘草次酸产量差异显著,分别为8 mg/L 与30 mg/L,这说明两种宿主中P450的活性存在差异。
本研究对两种工程菌进行转录组分析时发现,参与鞘脂合成的FAA1以及SUR2在Sgib 中显著高表达(表4),为验证这些差异对CYP72A63(T338S)催化特性的影响,本研究在GA160 中对其进行了敲除,结果表明SUR2的敲除显著改变了CYP72A63(T338S)的催化底物选择性。该基因敲除虽然降低了甘草次酸的合成能力,但是显著提高了 CYP72A63(T338S)对底物 11-氧-β-香树脂醇的选择性,抑制了甘草次酸合成过程中β-香树脂醇氧化导致的副产物11-脱氧甘草次酸的合成,使甘草次酸所占比例由82.9%提高至97.6%(图3)。SUR2敲除抑制了神经酰胺向植物神经酰胺的转化过程,造成神经酰胺的积累,从而改变了膜结构中鞘脂的组成成分,这说明膜结构中鞘脂结构的改变对膜结合的Ⅱ型细胞色素P450 酶的催化特性具有显著的影响。
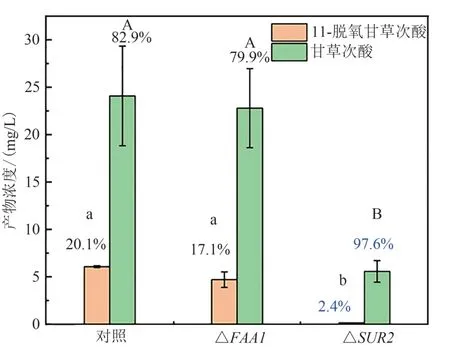
图3 敲除FAA1及SUR2的工程菌株产物分布Fig.3 Product profile of the strains with FAA1or SUR2 knock out

表4 不同宿主中脂质代谢相关基因的转录丰度Tab.4 Transcription levels of genes associated to lipid metabolism in different strains
2.3 调节鞘脂合成途径调控CYP72A63(T338S)底物选择性
SUR2的敲除对CYP72A63(T338S)的底物选择性产生了显著的影响,这表明脂质成分的改变,尤其鞘脂的成分和结构的改变对于P450 酶的催化特性影响显著。鞘脂由神经酰胺骨架、极性头和超长链脂肪酸(C18~C26)构成,鞘脂分子中超长链脂肪酸的长度的改变会对生物膜的结构产生影响,同时游离超长链脂肪酸的含量的改变对细胞膜结构也有一定的影响[34-35]。
在酿酒酵母中,脂肪酸碳链的延长由elo1p、elo2p、elo3p 三种酶催化,其中C18CoA 主要由elo1p 与 elo2p 合成,C26CoA 主要由 elo2p、elo3p 合成。因此,本研究对工程菌中编码这三种蛋白的基因ELO1、ELO2、ELO3基因进行了过表达,促进超长链脂肪酸合成。结果表明,三个基因的过表达对于CYP72A63(T338S)的底物选择性没有显著影响,11-脱氧甘草次酸和甘草次酸的产量没有产生显著变化,但是这些基因的过表达显著促进了其前体物质β-香树脂醇的积累,其可能原因为鞘脂结构的改变提高了膜结构对于无羧基三萜产物的容纳能力(图4)。

图4 过表达ELO1、ELO2、ELO3工程菌中三萜化合物的产量Fig.4 Production of triterpenoids in the engineered strains overexpressing ELO1,ELO2 and ELO3
植物来源的酶在酵母中进行异源表达时,活性低、特异性差是一种常见现象。由于细胞色素P450 酶为膜蛋白,影响其活性的原因之一可能为植物与酵母细胞内质网膜脂质成分的不同。
葡萄糖神经酰胺是植物鞘脂的主要成分之一,由鞘氨醇和脂酰-CoA经葡萄糖神经酰胺合酶催化合成[36],通过对酿酒酵母基因组进行检索发现,其基因组中不存在葡萄糖神经酰胺合酶及其同工酶,对酿酒酵母代谢组进行检索也未发现葡萄糖神经酰胺[37],该差异可能为导致植物来源的酶在酿酒酵母中催化特性差的原因。为验证葡萄糖神经酰胺对三萜合成的影响,本研究在酿酒酵母工程菌GA160的YPRCtau3位点中引入的来源于苜蓿(Mt)、毕赤酵母(Pa)以及解脂酵母(Ya)的葡萄糖神经酰胺合酶(GCS)。结果表明,来自于毕赤酵母的葡萄糖神经酰胺合酶PaGCS2提高了CYP72A63(T338S)对底物β-香树脂醇选择性,显著促进了11-脱氧甘草次酸的合成(图5)。综合以上结果,调节酵母鞘脂代谢,改变其组成对P450酶的催化特性具有显著的影响,通过控制鞘脂合成可实现对Ⅱ型P450酶底物选择性的调控。
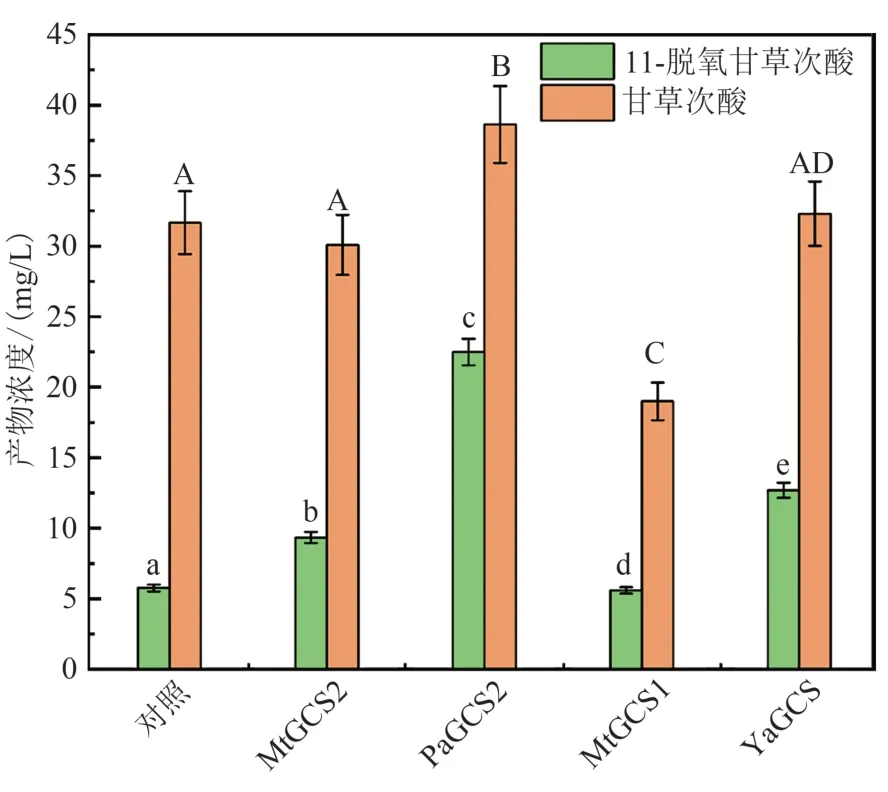
图5 过表达外源葡萄糖神经酰胺合酶工程菌的三萜产量Fig.5 Production of triterpenoids in the engineered strains overexpressing heterogenous GCSs
2.4 调控双步氧化比例提高CYP72A63(T338S)反应特异性
甘草次酸合成过程中,中间产物11-氧-β-香树脂醇以及副产物11-脱氧甘草次酸的合成前体均为β-香树脂醇,因此催化11-氧-β-香树脂醇合成的Uni25647 以及催化副产物11-脱氧甘草次酸合成的CYP72A63(T338S)之间存在底物竞争关系,通过提高Uni25647的活性,增强其竞争底物β-香树脂醇的能力,有望削弱CYP72A63(T338S)氧化β-香树脂醇合成11-脱氧甘草次酸的能力。强化酶的表达是提高酶催化能力的一种直接有效的方式,因此本研究在GA160基础上,将Uni25647基因的启动子更换成表达强度更高的Gal7启动子,构成菌株GA180-1。结果表明,更换强度更高的Gal启动子后对甘草次酸的合成无显著的促进作用,但显著促进了CYP72A63(T338S)对底物 11-氧-β-香树脂醇的特异性氧化,完全抑制了甘草次酸合成过程中副产物11-脱氧甘草次酸的合成,甘草次酸产量为(31.6 ±2.2)mg/L相比于对照(30.3±1.7)mg/L没有显著变化(图 6)。通 qPCR 对菌株 GA160 与 GA180-1 中Uni25647及CYP72A63(T338S)基因转录水平进行测定发现,在GA180-1中Uni25647基因转录水平提高了18.8倍,Uni25647与CYP72A63(T338S)转录丰度的比例由1∶24.5提高至1∶1.5。Uni25647表达水平的提高,强化了其对于β-香树脂醇的竞争力,导致β-香树脂醇代谢流均流向Uni25647 催化的11-oxo-βamyrin合成反应从而抑制了11-脱氧甘草次酸的合成,使工程菌11-氧-β-香树脂醇产量由(51.0±2.4)mg/L提高至(67.9±7.4)mg/L,由于CYP72A63(T338S)催化活性没有得到显著的促进,因此甘草次酸的产量并未获得显著的提高。这一结果说明通过调节具有底物竞争关系的催化酶比例的策略可实现对其反应、特异性的控制(图6)。
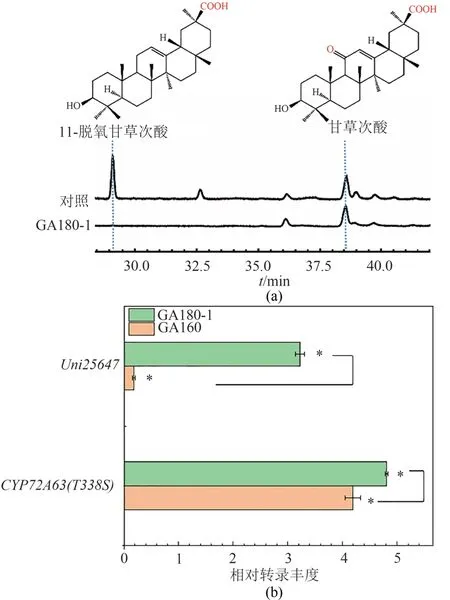
图6 Uni25647表达强化后工程菌的产物的分布(a)及P450酶的相对转录丰度(b)Fig.6 Product profile(a)and the relative transcription abundance of P450 enzymes after up-regulating Uni25647 expression(b)
3 讨论
相比于化学催化,P450 酶是一种高选择性的催化剂,具有更强的区域和立体选择性,但是许多P450 酶催化仍然具有一定的杂泛性,比如CYP72A63 可以氧化β-香树脂醇的 C-30 与 11-氧-β-香树脂醇的C-29和C-30两个位点,合成11-脱氧甘草次酸、甘草次醇、甘草次醛、29-羟-11-氧-β-香树脂、甘草次酸五种产物,其突变体CYP72A63(T338S)虽然可以实现良好的C-30区域选择性,但是可以同时氧化β-香树脂醇和11-氧-β-香树脂醇导致合成甘草次酸过程中同时合成副产物11-脱氧甘草次酸,即导致底物的消耗也增加了目标产物分离纯化的难度[18]。
要实现目标产物的高效选择性合成,需要进一步增强P450 酶的催化选择性。酶催化口袋的几何结构、化学环境等因素是决定其催化特性的关键,通过对P450 酶活性口袋进行重塑,获得了多种具有良好区域选择性、立体选择性等催化特性的突变体[14,34]。同时,研究发现与P450 酶存在蛋白与蛋白之间相互作用的氧化还原伴侣也会对P450 的催化特性产生显著影响。在Ⅱ型P450 酶中,除了CPR 与P450 酶的相互外,还存在膜、P450 酶和CPR 三者之间的相互作用。这些相互作用同样对P450 的催化特性具有显著影响,研究表明,膜组分磷脂酰胆碱是内质网中促进CPR 与P450 相互作用的主要成分,除此之外,另外一种组分磷脂酰乙醇胺能够促进CPR 与CYP2B4 之间的电子传递效率从而提高其催化活性[31]。为进一步探索膜对P450 酶的影响,本研究中对连接膜与蛋白的跨膜域以及膜组分鞘脂合成的关键途径进行了改造,发现跨膜域对CYP72A63(T338S)催化的底物选择性具有显著的影响,本研究中利用该特性开发了一种通过改造CYP72A63(T338S)跨膜域控制其底物选择性的策略。通过对鞘脂合成途径的调控发现,改变鞘脂成分也是一种实现对P450 催化底物选择性的调控的有效方法,然而鞘脂成分改变以及跨膜域改造对酶活性的调控机制并不清晰,在后期的研究中,通过蛋白质晶体结构解析、分子动力学模拟等方法,阐明膜结构以及P450 跨膜域结构变化对其催化结构域构象的影响将会促进鞘脂成分改变以及跨膜域改造对P450酶活性调控机制的解析。
除了改变催化剂本身的催化特性外,从化合物合成过程的角度出发,也可以为我们提供反应调控的新思路。在天然产物的生物合成过程中,由于酶的杂泛性,导致下游反应的酶可以催化上游酶的底物,在两种酶之间形成底物竞争关系,从而导致副产物的合成。这种情况下提高上游酶的催化活性,强化其底物竞争能力会显著降低甚至消除下游酶对上游底物的消耗,起到抑制副产物合成的作用。在本研究中,通过强化甘草次酸合成过程中的上游P450 酶Uni25647 的催化活性,增强其竞争底物β-香树脂醇的能力,完全消除了下游酶CYP72A63(T338S)对于β-香树脂醇的氧化,这也进一步说明,调控合成途径中具有底物竞争关系的酶的表达比例,也是一种改变酶催化的底物选择性、消除副产物产生的有效策略。
不同于传统的P450 酶催化结构域改造以及还原伴侣工程来调控P450 酶的催化特性,本研究通过改造P450 酶的跨膜结构域以及调控膜组分的代谢实现了Ⅱ型P450 酶催化的底物选择性的调控,改变了CYP72A63(T338S)对于两种底物β-香树脂醇以及11-氧-β-香树脂醇的氧化选择性;同时本研究发现,通过改变具有底物竞争关系的酶的表达比例,是一种消除级联反应过程中由于上下游酶均可氧化同一种底物而导致的副产物的有效方法,为Ⅱ型P450 酶催化特异性的调控提供了更加丰富的策略,然而跨膜域结构以及膜组分代谢对于P450催化特性影响的机制仍需更加深入的研究。
