环境因子对浙江沿岸春季日本鲐(Scomber japonicus)幼鱼生长、分布的影响*
2021-11-27朱文斌崔国辰胡翠林朱海晨王雅丽蒋日进张亚洲冯春雷
朱文斌 崔国辰 胡翠林 朱海晨 王雅丽 蒋日进 张亚洲 冯春雷
环境因子对浙江沿岸春季日本鲐()幼鱼生长、分布的影响*
朱文斌1, 2, 3, 4崔国辰2胡翠林1, 2, 3, 4朱海晨2王雅丽2蒋日进1, 2, 3, 4张亚洲1, 2, 3, 4冯春雷5①
(1. 浙江省海洋水产研究所 舟山 316021; 2. 浙江海洋大学海洋与渔业研究所 舟山 316021; 3. 农业农村部重点渔场渔业资源科学观测实验站 舟山 316021; 4. 浙江省海洋渔业资源可持续利用技术研究重点实验室 舟山 316021; 5. 中国水产科学研究院东海水产研究所 上海 200090)
浙江沿岸海域是日本鲐()的重要产卵场。当前针对日本鲐幼鱼生长发育的研究较少, 而环境因子会对其生长、分布造成较大影响。根据2019年5月的科学观察员取样数据和环境数据, 对其群体组成进行研究, 运用Pearson相关性分析、广义加性模型(GAM)探究其分布及个体大小的影响因子, 利用克里金插值法定量分析溶解氧对日本鲐幼鱼分布影响。结果表明: 海水表层盐度(Sea Surface Salinity, SSS)是影响日本鲐幼鱼分布的主要环境因子, 幼鱼集中分布于27.5—27.8盐度海域; 浙江沿岸南部海域日本鲐幼鱼叉长、体重均值分别为37.6 mm、0.659 g, 中部海域叉长、体重均值分别为64.2 mm、3.305 g, 南部、中部海域间鲐幼鱼大小存在显著性差异(<0.05); 日本鲐幼鱼个体大小主要受溶解氧和纬度影响, 个体大小与溶解氧呈多波峰的非线性关系, 随纬度增加呈先减小再增大趋势。研究表明: 溶解氧是影响浙江沿岸日本鲐幼鱼生长、分布的关键环境因子; 结合幼鱼分布特征及个体大小差异, 推测浙江沿岸海域日本鲐幼鱼来自不同产卵批次。
日本鲐; 幼鱼; 广义加性模型; 分布; 个体大小; 环境因子
日本鲐()属中上层鱼类, 2018年全国日本鲐捕捞量为43.25×104t (农业农村部渔业渔政管理局等, 2019), 其主要捕捞方式有灯光围敷网、疏目浮拖网, 由于资源量丰富且易于集群捕捞, 成为我国重要经济鱼类。日本鲐是东、黄海最重要的经济鱼类之一(郑元甲等, 2003), 严利平等(2010)利用体长结构VPA评估东海鲐类资源量为39.55×104t。浙江沿岸水系分布复杂, 营养物质丰富, 是多种渔业生物的产卵场和索饵场, 有关该海域的渔业研究多集中于鱼虾蟹类的空间生态位及群落结构分析(胡翠林等, 2018; 丁朋朋等, 2019; 严润玄等, 2020), 针对单一物种分布、生长的研究较少。
幼鱼是指未达初次性成熟时的鱼, 其生长发育是鱼类种群再生能力的基础, 故幼鱼的生长及数量变化对其资源变动具有重要影响。然而, 幼鱼的分布及生长对栖息环境的变化敏感度高(Kim, 2002), 栖息海域的冷水团季节性变化、溶解氧浓度、食物条件等均是重要的影响因素(李洁, 2011; 胡艳, 2015; 朱庆环等, 2020; 武瑞等, 2020)。截止目前, 关于日本鲐的研究主要集中在生物学特性、栖息地变动与环境关系、分子地理学等(严利平等, 2012; 李建生等, 2014; 陈爽等, 2019), 由于传统的底拖网调查较难获取日本鲐幼鱼样本, 因此针对日本鲐幼鱼分布及生长的研究报道较少。找出影响日本鲐幼鱼分布及生长的关键环境因子, 定量分析各因子的作用, 可为日本鲐资源养护提供基础。
2017年开始, 我国的伏季休渔开始时间从6月1日提前到5月1日, 因此, 利用专项特许渔业采集5月份近海日本鲐幼鱼数据, 研究春季日本鲐幼鱼生长、分布具有较高价值。本文基于2019年5月浙江沿岸海上科学观察员取样数据, 结合同期水质仪采集环境数据, 对日本鲐幼鱼资源分布、个体大小与环境关系进行研究, 以期为该海域日本鲐资源保护积累科学素材。
1 材料与方法
1.1 数据来源及处理
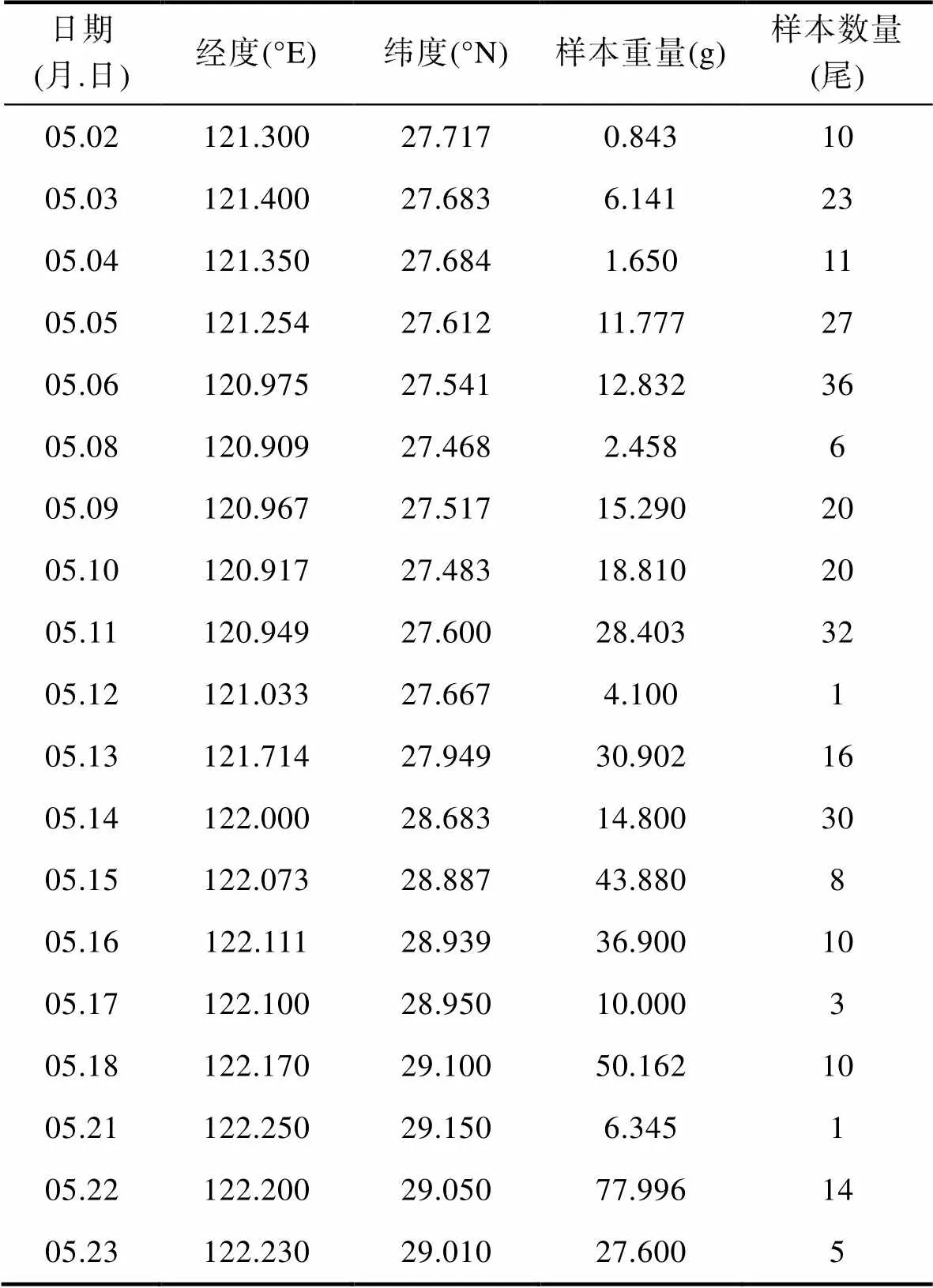
渔业数据来源于2019年5月2—23日期间伏季休渔专项特许捕捞的“华盛渔加2号”的渔捞日志及配套科学观察员记录, 包括作业日期、作业位置(经纬度)、日本鲐幼鱼尾数、体重等(表1)。捕捞渔具为双船有囊围网, 网衣上下纲长300 m, 网口网目1.8 m, 囊网长度40 m, 渔船两艘为1组, 共8组, 每天作业时长为12 h。生产期间每天上午、下午各进行一次采样, 每次随机选取1 kg渔获物进行生物学分析, 并采集渔获物冰鲜带回实验室统计体重(精确到0.001 g)。根据取样样品日本鲐重量以及每天捕捞总量可以计算出每天日本鲐产量。
作业点环境数据来源于多功能水质监测仪(JFE-AAQ171), 包括水温、水深、叶绿素(chlorophyll-, chl)浓度、盐度、溶解氧等。日本鲐作为中上层暖水性鱼类, 主要受表层环境影响(李建生等, 2015), 因此选择0.5 m水层所测得的水温、盐度、chl、溶解氧为海洋环境数据。本文研究区域为: 27°00′N—29°30′N, 120°30′E—122°30′E, 因浙江沿岸春季调查海域范围广, 不同日期间调查船位存在明显差异, 将2—13日日本鲐幼鱼分为南部组, 14—23日分为中部组。
1.2 分析方法
1.2.1 叉长-体重关系拟合 日本鲐幼鱼叉长-体重关系采用幂函数拟合(费鸿年等, 1990), 表达式为:
=aL, (1)
其中,为体重,表示叉长,、为参数。为条件因子, 常表征鱼群生存环境优越程度;为异速生长因子, 当=3时, 表示鱼类个体等速生长;>3时, 表示鱼体呈正异速增长;<3时, 鱼体呈负异速增长(Froese, 2006)。
表1 浙江沿岸5月日本鲐幼鱼生产数据
Tab.1 The production data of S. japonicus juvenile in May in the area of the coast of Zhejiang
1.2.2 鲐幼鱼丰度 日本鲐幼鱼名义CPUE以单位时间内单船捕捞渔获量[kg/(h·艘)]表示, 其计算公式为:

其中, CPUE为单位捕捞努力量渔获量,为日本鲐渔获量,为作业时间,为作业船数。
1.2.3 不同因子对日本鲐幼鱼分布、生长的影响 本研究选择温度、chl、溶解氧以及盐度作为环境因子, 经度、纬度作为空间因子。建立模型之前利用相关性分析对影响因子进行筛选, 具有显著相关性的两个因子保留一个(李迎冬等, 2021)。本研究以日本鲐幼鱼CPUE、平均叉长分别作为响应变量, 利用GAM模型对日本鲐幼鱼分布与影响因子、个体大小与影响因子分别进行回归分析。GAM模型的一般形式为(徐国强等, 2018):

式中,()为联结函数,0为截距,f(x)是各环境因子的非参数函数,是服从正态分布的随机误差项。利用AIC准则选择最优模型, 往基础模型中不断加入影响因子, AIC值最小时对应的模型为最优模型, 最终利用检验评估因子的显著性。
本研究采用SPSS 17.0软件进行Pearson相关性分析, Microsoft Excel 2016软件计算幼鱼CPUE, Arcgis 10.2软件绘制溶解氧分布图, R语言mgcv包进行GAM模型拟合与绘图。
2 研究结果
2.1 群体组成及CPUE变化
南部、中部组叉长范围分别为12—77 mm、31—95 mm, 平均叉长为37.6 mm、64.2 mm, 优势叉长范围为20—60 mm (95.54%)、30—90 mm (98.77%) (图1a); 体重范围分别为0.040—4.100 g、0.200— 8.880 g, 平均体重为0.659 g、3.305 g, 优势体重范围为0—2 g (98.02%)、0—7.5 g (96.30%) (图1b)。对南部、中部组进行独立样本检验, 检验结果表明两组叉长数据存在显著性差异(= –11.778, d=104.399,<0.05), 即中部海域日本鲐幼鱼叉长显著高于南部海域; 两组体重数据同样存在显著性差异(= –9.334, d=83.873,<0.05), 即中部海域日本鲐幼鱼体重显著高于南部海域。
日本鲐幼鱼CPUE随日期变化整体呈波浪式上升趋势, 变化范围为0.05—12.65 kg/(h·艘)。2—12日CPUE波动较小, 13—23日波动剧烈, 高低CPUE交替出现(图2)。

图1 日本鲐幼鱼叉长、体重频率分布

图2 浙江沿岸海域日本鲐幼鱼CPUE日变化
2.2 叉长-体重间关系
日本鲐幼鱼叉长与体重的拟合结果如图3, 幼鱼总体间叉长-体重的关系式为:=9×10–6×3.016 4(2=0.958 6,=283) (图3a)。经ANCOVA检验, 日本鲐幼鱼南部、中部组间叉长-体重关系存在显著性差异(<0.05), 故将两组幼鱼叉长体重数据分开分析, 得出南部组叉长-体重关系式为:=10–5×2.907(2=0.933 9,=202) (图3b); 中部组叉长-体重关系式为:=6×10–7×3.647 2(2=0.988 4,=81) (图3c)。
2.3 幼鱼分布与影响因子关系
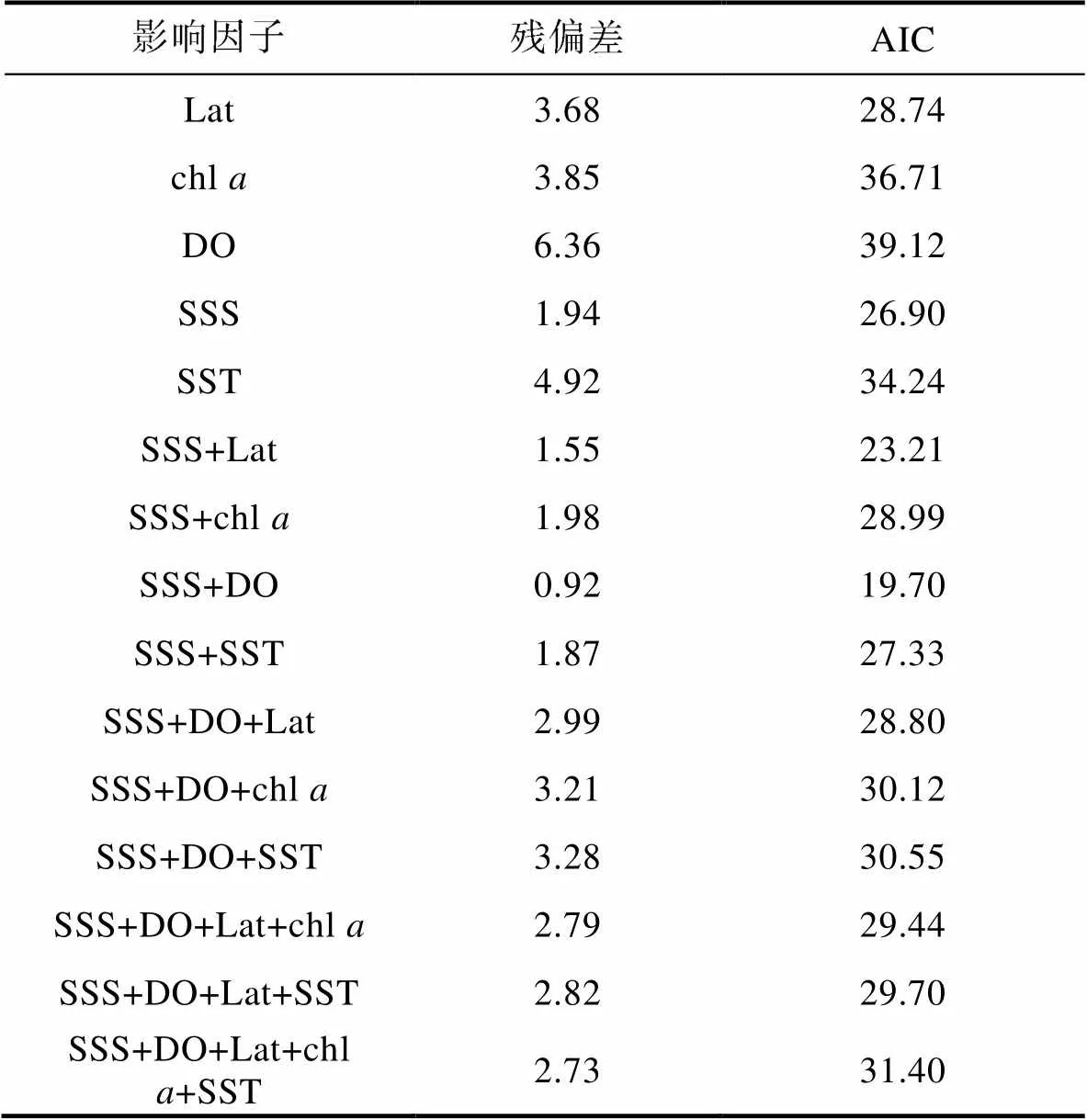
利用AIC准则筛选因子, 筛选过程如表2。根据AIC最小原则得到最优GAM模型, 模型表达式为:
lg(CPUE+1) =0+(SSS) +(DO) +, (4)
式中, CPUE表示日本鲐幼鱼单位捕捞努力量渔获量,0为截距,(SSS)为盐度效应,(DO)为溶解氧效应,是服从正态分布的随机误差项。GAM模型偏差分析显示, 所选建模因子的累计偏差解释率为86.2%, 盐度对日本鲐幼鱼分布影响显著(<0.05) (表3)。
盐度、溶解氧对于日本鲐幼鱼分布的影响如图4。本次调查的日本鲐幼鱼盐度范围在27.0—29.3之间, 集中分布于27.5—27.8盐度海域, CPUE随盐度增加呈先下降后上升再下降趋势。
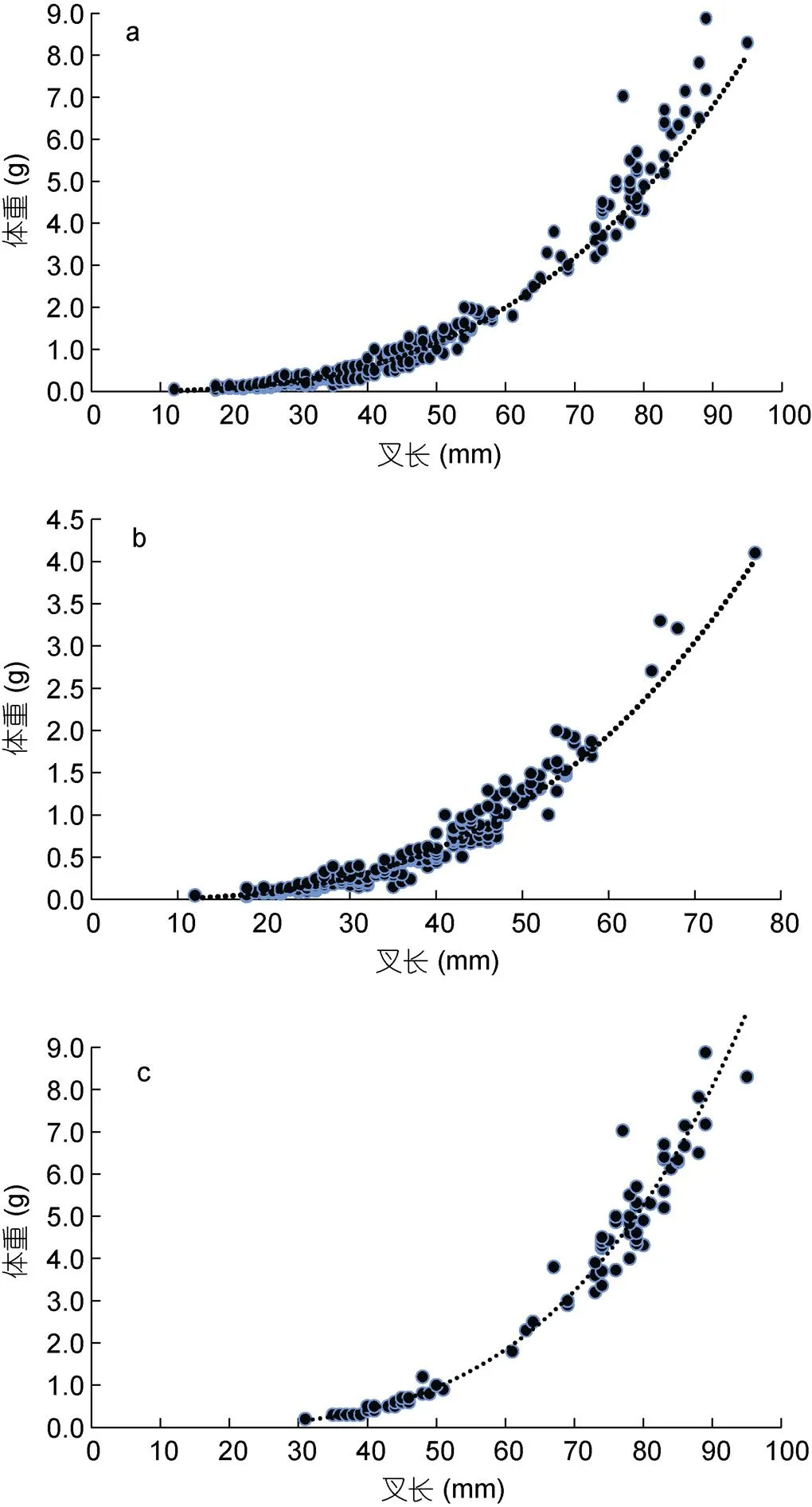
图3 浙江沿岸春季日本鲐幼鱼叉长-体重关系
注: a. 总体; b. 南部; c. 中部
表2 GAM模型变量筛选及影响因子的参数分析
Tab.2 The variable screening process for GAM and parameters analysis
注: Lat表示纬度; chl表示叶绿素; DO表示溶解氧; SSS表示盐度; SST表示水温
表3 CPUE与盐度、溶解氧的GAM模型拟合检验表

Tab.3 GAM model fitting test sheet for CPUE and salinity and dissolved oxygen
注: *表示0.05水平上显著相关(<0.05)
2.4 不同因子对日本鲐幼鱼叉长的影响
利用AIC准则筛选因子, 筛选过程如表4所示。

图4 溶解氧、盐度对浙江沿岸日本鲐幼鱼CPUE的影响
表4 GAM模型变量筛选及影响因子的参数分析
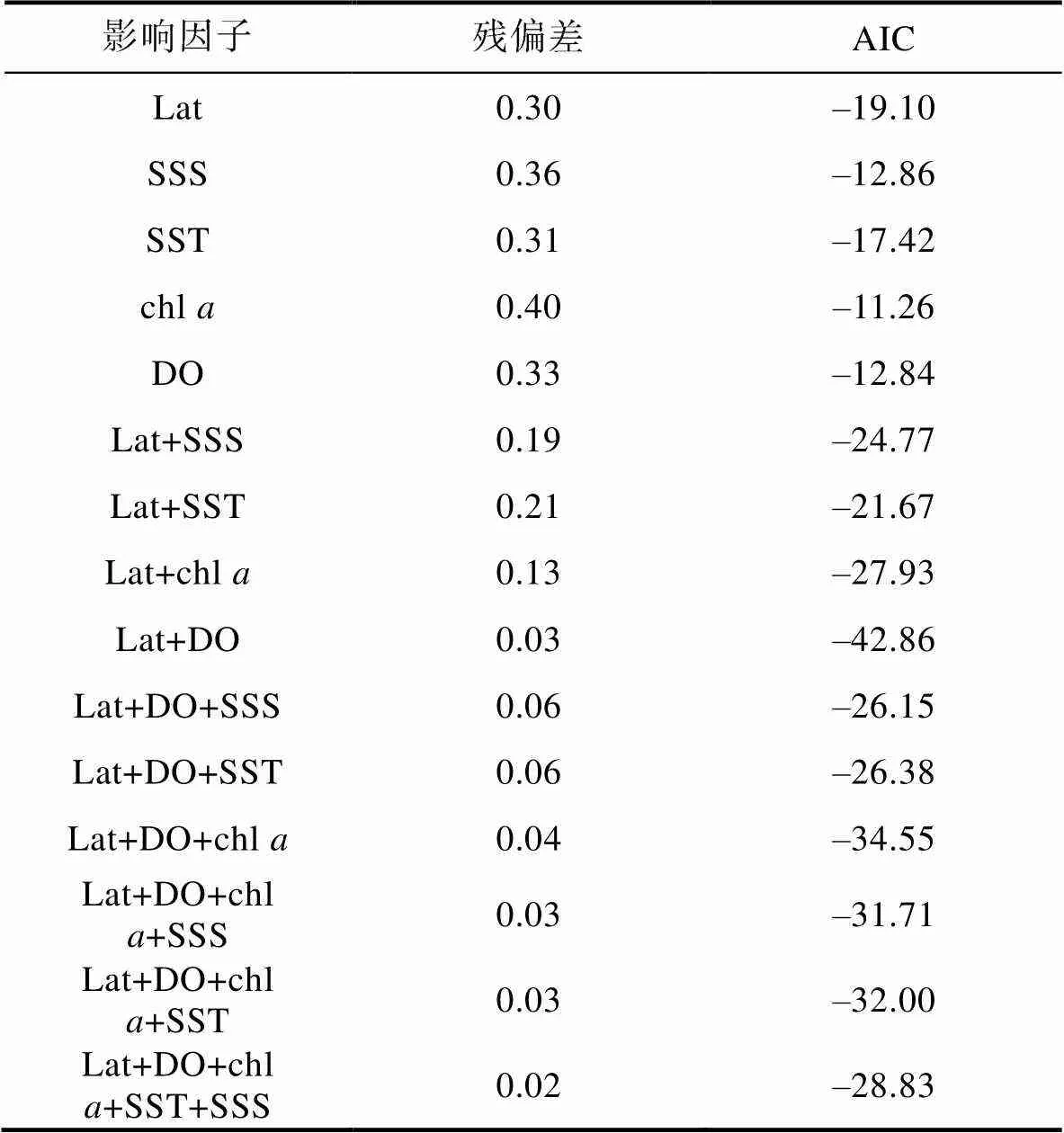
Tab.4 The variable screening process for GAM and parameters analysis
注: Lat表示纬度; SSS表示盐度; SST表示水温; chl表示叶绿素; DO表示溶解氧
根据AIC最小原则得到最优GAM模型, 模型表达式为:
lg(+1) =0+(Lat) +(DO) +, (5)
式中,为日本鲐幼鱼平均叉长,0为截距, Lat为纬度, DO为溶解氧,是服从正态分布的随机误差项。GAM模型检验表显示, 纬度、溶解氧对日本鲐幼鱼个体大小的影响显著(<0.05) (表5)。
表5 平均叉长与纬度、溶解氧的GAM模型拟合检验表
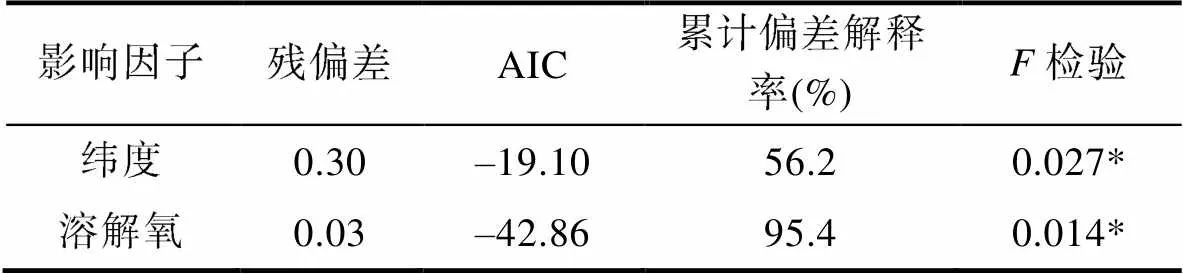
Tab.5 GAM model fitting test sheet for average fork length to latitude and dissolved oxygen
注: *表示0.05水平上显著相关(<0.05)
纬度、溶解氧对于日本鲐幼鱼叉长的影响如图5。日本鲐幼鱼叉长随纬度增加呈先减小再增加趋势; 日本鲐幼鱼分布海域溶解氧范围为8.0—11.5 mg/L, 集中分布于8.0—9.2 mg/L海域。在8.0—10.8 mg/L范围内, 幼鱼叉长随溶解氧增加呈波浪式上升趋势, 之后日本鲐幼鱼叉长随溶解氧增加而不断减小。
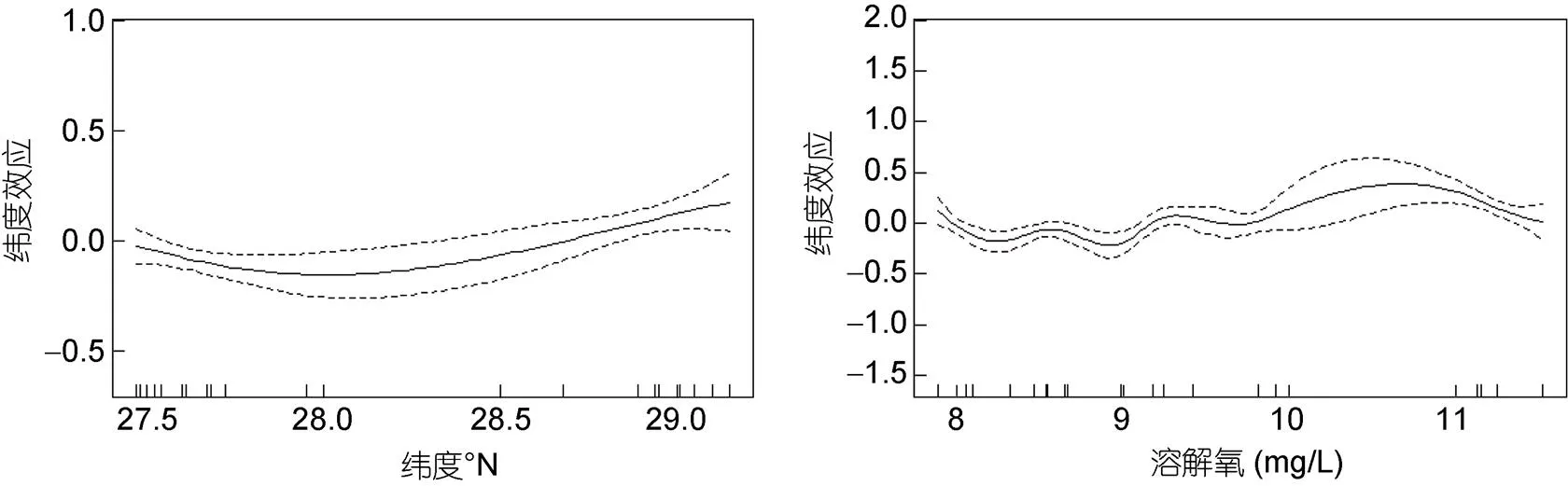
图5 纬度、溶氧含量对日本鲐幼鱼平均叉长的影响
3 讨论与分析
3.1 分布变化及其与环境因子关系
多数具备游泳能力的幼鱼有主动选择生境的能力, 故幼鱼分布范围的变化一定程度上体现了环境的驱动作用(李曰嵩等, 2013)。本研究GAM模型拟合结果表明, 日本鲐幼鱼分布主要受盐度、溶解氧影响。
日本鲐幼鱼的资源分布与海域间盐度范围差异有关, 盐度会直接影响鱼体内外渗透压, 从而对幼鱼的分布、生长造成影响(Pielou, 2011)。调查数据显示南部海域盐度为27.06—29.29, 中部为27.07—28.10, 模型结果显示幼鱼适宜盐度范围为27.5—27.8 (图4),因此中部海域盐度更加适宜幼鱼集群。此外日本鲐幼鱼的饵料以桡足类为主, 盐度对于海洋桡足动物的摄食有显著影响, 中华哲水蚤()的摄食率在盐度为28时达到最高(于娟等, 2012), 这也进一步佐证了盐度影响桡足类丰度从而影响日本鲐幼鱼分布。
溶解氧升降会引起鱼类数量和空间分布的变化。Kramer等(1982)对多种热带鱼类进行研究发现低氧条件下许多鱼类会探测并回避低氧水域; 曾嘉维等(2019)研究雷州湾附近海域鱼类群落结构与环境的关系, 发现溶解氧是春、秋、冬季影响鱼类空间分布的重要环境因子。在本研究中, 中部幼鱼分布海域溶解氧含量较南部高(图6)。这是因为中部幼鱼个体较南部海域大, 个体不同的幼鱼耗氧量存在差异。研究证明, 体重是影响水生动物耗氧量的重要因素之一, 通常个体较小的幼鱼耗氧量比个体大的要低(闫茂仓等, 2007)。
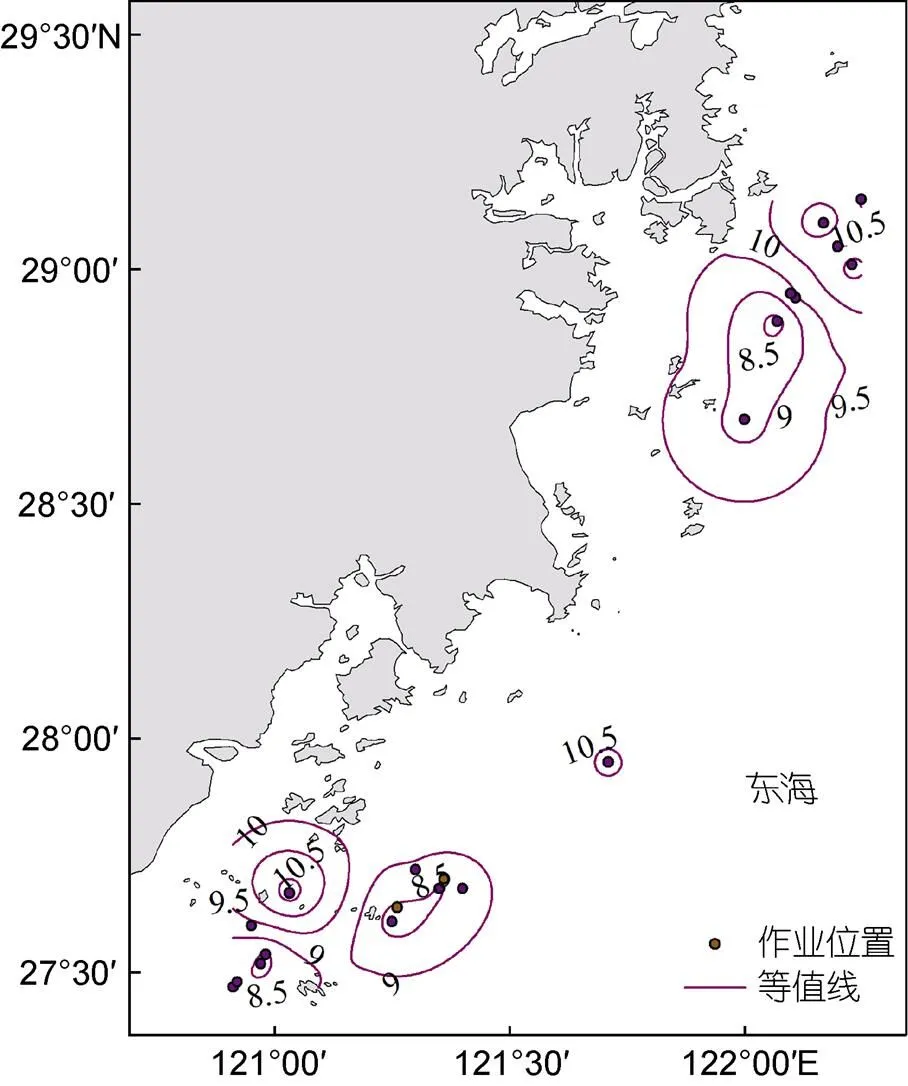
图6 日本鲐幼鱼与溶氧含量分布
苏杭等(2015)研究认为水温与日本鲐时空分布有显著关系, 与本研究结果存在差异。原因可能是日本鲐空间分布主要受洄游机制影响(崔科等, 2007), 而本研究时间、空间跨度小, 海域间水温差异不大, 因此两者关系不明显。此外, 这种现象可能与日本鲐生活史对策有关(王国强等, 2019)。日本鲐成鱼需要完成觅食、繁殖等多种复杂生态过程, 栖息地环境变动较大, 本文研究对象为日本鲐幼鱼, 活动范围有限, 因此温度耐受幅较成鱼小。
除盐度和溶解氧外, 海流也会对日本鲐幼鱼分布产生重要影响, 如李曰嵩等(2018b)发现台湾暖流的强弱影响鲐鱼渔场的位置和渔期。本文因采样条件限制未添加海流数据, 今后还可以结合海流数据进一步研究。
3.2 个体大小差异及其与影响因子关系
值表示鱼的体质量增加系数与叉长增长系数之比(黄真理等, 1999), 其大小与营养条件和生活阶段的不同有关(刘勇等, 2006)。南部日本鲐幼鱼值小于3, 主要是幼鱼其早期阶段进行骨骼化生长导致个体偏细长(李曰嵩等, 2018a)。南部至中部值变大, 说明幼鱼体重的增速大于叉长的增速, 因此南部、中部海域间日本鲐幼鱼值差异的原因可能是发育阶段不同。有研究证明幼鱼耗氧量与鱼体大小呈正相关关系(崔前进等, 2018), 本文南部、中部幼鱼分布海域溶氧量存在差异, 南部海域溶解氧集中于8.5—9.0 mg/L, 中部集中于9.5—10.5 mg/L (图6), 这与个体大的幼鱼分布海域溶氧量高结果一致(图6)。
本研究27.5°N—28.0°N范围内日本鲐幼鱼个体叉长随纬度增加而减小, 28.0°N—29.2°N范围内日本鲐幼鱼个体叉长随纬度增加而增大(图5)。刘勇等(2006)认为日本鲐个体随纬度增加而增大很可能与海水温度有关, 本研究日本鲐幼鱼个体大小与纬度并非线性关系, 原因可能是南部、中部海域间日本鲐鱼个体大小不同, 耐受水温范围存在差异, 超过耐受水温范围生长受到抑制(吴欢欢等, 2020)。此外, 纬度是表征温度、chl等各种因子的综合, 不同纬度海域饵料生物会存在差异, 从而影响日本鲐幼鱼个体大小。
中部海域日本鲐幼鱼平均叉长、体重较南部海域均有明显增加。一方面, 幼鱼个体随时间推移不停生长; 另一方面, 随着幼鱼游泳能力增强, 可迁移到更适宜的环境。李曰嵩等(2013)对东海日本鲐游泳能力进行研究, 发现5月末仔幼鱼逐渐有能力游向并滞留适宜海域。李建生等(2020)发现日本鲐产卵群体从东海中南部外海游向浙中南海域的过程中分批产卵, 因此推测南部、中部海域幼鱼来自不同产卵批次, 这可能也是两组幼鱼个体大小存在明显差异的原因。但仅靠一年数据推断两组幼鱼来自不同产卵批次是不够的, 还需多年数据进一步考证。
4 结论
(1) 海水表层盐度是影响浙江沿岸日本鲐幼鱼分布的主要环境因子, 幼鱼集中分布于27.5—27.8盐度海域; 日本鲐幼鱼个体大小主要受溶解氧和纬度影响, 个体大小与溶解氧呈多波峰的非线性关系, 随纬度增加呈先减小再增大趋势; 溶解氧是影响浙江沿岸日本鲐幼鱼生长、分布的关键环境因子。
(2) 结合幼鱼分布特征、个体大小差异以及相关学者研究, 推测此次浙江沿岸日本鲐幼鱼来自不同产卵批次。
丁朋朋, 高春霞, 彭 欣等, 2019. 浙江南部近海主要虾类的时空生态位及种间联结性. 应用生态学报, 30(11): 3942—3950
于 娟, 张 瑜, 杨桂朋等, 2012. 饵料、温度和盐度对两种海洋桡足类摄食与排粪的影响. 中国海洋大学学报, 42(7/8): 45—52
王国强, 夏继刚, 2019. 恒温与变温对不同生活史阶段斑马鱼热耐受性的影响. 生态学杂志, 38(7): 2133—2137
朱庆环, 田永军, 张 弛等, 2020. 基于耳石微结构的黄、渤海蓝点马鲛幼鱼日龄与生长. 海洋学报, 42(2): 87—95
刘 勇, 严利平, 程家骅, 2006. 东海北部和黄海南部日本鲐鱼生长特性及合理利用. 中国水产科学, 13(5): 814—822
闫茂仓, 单乐州, 邵鑫斌等, 2007. 温度及体重对鮸鱼幼鱼耗氧率和排氨率的影响. 热带海洋学报, 26(1): 44—49
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2019. 2019中国渔业统计年鉴. 北京: 中国农业出版社
严利平, 李建生, 凌建忠等, 2010. 应用体长结构VPA评估东海西部日本鲭种群资源量. 渔业科学进展, 31(2): 16—22
严利平, 张 辉, 李圣法等, 2012. 东、黄海日本鲭种群鉴定和划分的研究进展. 海洋渔业, 34(2): 217—221
严润玄, 冯 明, 王晓波等, 2020. 浙江北部海域大型底栖动物优势种的时空分布. 海洋与湖沼, 51(5): 1162—1174
苏 杭, 陈新军, 汪金涛, 2015. 海表水温变动对东、黄海日本鲐鱼栖息地分布的影响. 海洋学报, 37(6): 88—96
李 洁, 2011. 限制溶解氧供应对褐牙鲆幼鱼生长的影响及其机制的实验研究. 青岛: 中国海洋大学硕士学位论文
李曰嵩, 白松麟, 潘灵芝等, 2018a. 基于个体模型水温变动对东海鲐鱼补充量影响模拟研究. 海洋湖沼通报, (6): 118—124
李曰嵩, 潘灵芝, 官文江等, 2018b. 基于个体的东海鲐鱼补充量年际变动模拟研究. 海洋学报, 40(1): 87—95
李曰嵩, 潘灵芝, 陈新军等, 2013. 东海鲐鱼()仔幼鱼游泳行为对输运和补充量的影响. 海洋与湖沼, 44(2): 318—325
李迎冬, 张崇良, 纪毓鹏等, 2021. 山东半岛南部海域小黄鱼时空分布及其与环境因子的关系. 中国水产科学, 28(4): 442—450
李建生, 严利平, 胡 芬, 2020. 基于鱼卵仔鱼数据的东海中南部日本鲭产卵场分析. 海洋渔业, 42(1): 10—19
李建生, 严利平, 胡 芬等, 2015. 温台渔场日本鲭的繁殖生物学特征. 中国水产科学, 22(1): 99—105
李建生, 胡 芬, 严利平, 2014. 台湾海峡中部日本鲭产卵群体生物学特征的初步研究. 应用海洋学学报, 33(2): 198—203
吴欢欢, 张运海, 陈 文等, 2020. 水温对前鳍高原鳅幼鱼生长和存活的影响. 淡水渔业, 50(5): 25—30
陈 爽, 郭 爱, 陈新军, 2019. 海表温度变化时近海日本鲭栖息地时空分布预测. 水产学报, 43(3): 593—604
武 瑞, 李建超, 叶振江等, 2020. 黄海大头鳕幼鱼的生长和分布特征. 中国海洋大学学报, 50(7): 63—73
郑元甲, 陈雪忠, 程家骅等, 2003. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 348—357
胡 艳, 2015. 长江口近岸水域幼鱼时空分布及代表种资源评估. 上海: 上海海洋大学硕士学位论文
胡翠林, 张亚洲, 李德伟等, 2018. 浙江沿岸产卵场春、夏季鱼类资源密度和群落多样性研究. 水生生物学报, 42(5): 984—995
费鸿年, 张诗全, 1990. 水产资源学. 北京: 中国科学技术出版社, 1—21
徐国强, 朱文斌, 张洪亮等, 2018. 基于GAM模型分析印度洋大眼金枪鱼和黄鳍金枪鱼渔场分布与不同环境因子关系. 海洋学报, 40(12): 68—80
黄真理, 常剑波, 1999. 鱼类体长与体重关系中的分形特征. 水生生物学报, 23(4): 330—336
崔 科, 陈新军, 2007. 东黄海鲐鱼资源丰度与表温关系. 南方水产, 3(4): 20—25
崔前进, 尚胜男, 蔡忠璐等, 2018. 盐度、温度和体质量对钝吻黄盖鲽幼鱼排氨率和耗氧率的影响. 上海海洋大学学报, 27(1): 64—72
曾嘉维, 林 坤, 王学锋等, 2019. 雷州湾附近海域鱼类群落结构及其与环境因子的关系. 中国水产科学, 26(1): 108—117
Froese R, 2006. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations. Journal of Applied Ichthyology, 22(4): 241—253
Kim H Y, Sugimoto T, 2002. Transport of larval jack mackerel () estimated from trajectories of satellite-tracked drifters and advective velocity fields obtained from sequential satellite thermal images in the eastern East China Sea. Fisheries Oceanography, 11(6): 329—336
Kramer D L, McClure M, 1982. Aquatic surface respiration, a widespread adaptation to hypoxia in tropical freshwater fishes. Environmental Biology of Fishes, 7(1): 47—55
Pielou E C, 2011. An introduction to mathematical ecology. Bioscience, 24(2): 7—12
EFFECTS OF ENVIRONMENTAL FACTORS ON GROWTH AND DISTRIBUTION OF YOUNG CHUB MACKERELALONG THE COAST OF ZHEJIANG IN SPRING
ZHU Wen-Bin1, 2, 3, 4, CUI Guo-Chen2, HU Cui-Lin1, 2, 3, 4, ZHU Hai-Chen2, WANG Ya-Li2, JIANG Ri-Jin1, 2, 3, 4, ZHANG Ya-Zhou1, 2, 3, 4, FENG Chun-Lei5
(1.Zhejiang Marine Fisheries Research Institute,Zhoushan 316021, China; 2.Marine and Fisheries Institute of Zhejiang Ocean University, Zhoushan 316021, China; 3.Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds,Ministry of Agriculture and Rural Affairs, Zhoushan 316021, China; 4. Key Laboratory of Sustainable Utilization of Technology Research for Fishery Resources of Zhejiang Province, Zhoushan 316021,China; 5. East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences College of Marine Science, Shanghai 200090,China)
The coastal waters of Zhejiang Province is one of the important spawning grounds for. Currently, few studies focus on the growth and development of juvenile, and environmental factors have a great influence on their growth and distribution. Based on scientific observer samplings and environmental data collected in May 2019, we analyzed the distribution and growth status ofjuvenile off the coast of Zhejiang Province. We studied the population composition, explored the influencing factors of its distribution and body size using the Pearson correlation analysis and generalized additive model, and analyzed the influence of environmental factors on the distribution of mackerel juvenile quantitatively by using Kriging interpolation. The results show that the distribution of juvenileis concentrated in salinity waters of 27.5—27.8, which implies sea surface salinity is the main environmental factor affecting the abundance distribution. There were significant differences in size between the southern and central coastal waters of Zhejiang (<0.05). The body size of young juvenile was mainly influenced by dissolved oxygen and latitude and had a nonlinear multi-peaks relationship with dissolved oxygen. In detail, the trend of their body size decreased first and then increased with latitude. Combing the distribution characteristics with body size differences, it is concluded that these juvenilemay come from different spawning batches.Key words; juvenile; generalized additive model; distribution; body size; environmental factors
* 国家重点研发计划项目, 2019YFD0901505号; 浙江省重点研发计划项目, 2018C02026号; 浙江省自然科学基金项目, LGN21C190005号; 限额捕捞关键技术研究与制度探索, 农财发[2017]36号。朱文斌, 博士, 高级工程师, E-mail: foolse@126.com
冯春雷, 副研究员, E-mail: fengmaster@126.com
2021-05-17,
2021-07-12
S932
10.11693/hyhz20210500117