浸种对西蒙得木种子萌发阶段贮藏物质动员的影响
2021-11-24李明蔓邓祥胜刘世男
李明蔓, 邓祥胜, 杨 梅, 刘世男, 程 飞
(广西大学林学院, 南宁 530004)
西蒙得木(SimmondsiachinensisLink)是双子叶植物纲西蒙得木科西蒙得木属的常绿沙漠灌木,种子富含45%~50%的液态蜡脂,蜡脂具有天然、纯净、性能稳定、燃点高、沸点低、抗氧化等特性,是化妆品工业不可取代的基本原料,是航空航天及精密仪器专用高级润滑油;医药上,有防治癌症、高血压、冠心病、胃外伤等病的作用,故其开发和经济价值很高,被誉为“液体黄金”[1]。西蒙得木原产于美国和墨西哥的“索诺拉荒漠”地区,现已被广泛引种到以色列、阿根廷等国家,创造了巨大的经济价值,产生了良好的生态效益。
西蒙得木原产地“索诺拉荒漠”地区,地属亚热带,气候干旱少雨,年平均气温21.4~23 ℃,年降雨量210~634 mm。诸远章[1]通过对西蒙得木试种点与原产地生态因子和植被进行对比分析认为,影响西蒙得木生长发育的主要生态因子是气温、雨量和土壤,在年平均气温19.4~23.9 ℃、年降雨量500~800 mm、冲积沙壤土上西蒙得木生长良好。朱大业等[2]根据西蒙得木生态学特性和气候相似引种理论、模糊识别原理,对西蒙得木原产地和引种地的气候条件进行分析对比,划分出国内适宜引种区、较适宜引种区、可能引种区、不宜引种区,其中广西南宁属于适宜引种区。本课题组于2017年8月在南宁试种了一批西蒙得木,2019年11月首次开花,说明西蒙得木在南宁可由营养生长转向生殖生长,有在南宁引种成功的可能(图1)。

图1 2021年2月西蒙得木雄花(广西南宁)
已有引种研究表明,西蒙得木适宜栽植气候条件为:极端低温(>-5 ℃),年平均气温>18 ℃,>10 ℃的有效积温不少于5 500 ℃,光照时数不少于1 700 h[3]。广西南宁各地年平均气温为20.9~21.9 ℃,气温年较差为15.2~16.6 ℃,≥10 ℃积温约7 527 ℃,年日照时数可达1 705 h,可见广西南宁具备西蒙得木适宜生长的气候条件。然而,南宁各地年降水量为1 013~1 505 mm[4],远高于原产地,略高于四川会东(700~1 100 mm)。因此,作为旱生植物,西蒙得木在南宁能否引种成功,取决于其对水分的适应能力、土壤质地情况和人为土壤改良措施。
当前,国内西蒙得木繁育以种子繁殖为主,还未商业化无性繁殖苗木。尽管西蒙得木不似其他生活在沙漠地区的一年生植物,种子遇适宜降水便迅速完成生活史,但因其种皮薄,对水分具有较高敏感性,短时间浸种(如4 h)或放在湿润的发芽纸上,于恒温恒湿条件下种子也能萌发。短期水分处理下,种子吸水未达到饱和,部分种子种皮仍保持褶皱,不够饱满,但浸种24 h的种子会腐烂且出苗率较低[5]。因此,探究浸种时间对西蒙得木种子萌发的影响具有重要意义。
浸种可使种子加速吸水,充分膨胀,软化种皮,提高水解酶活性,促进贮藏物质水解,达到有利于种子萌发的状态[6-7],从而使播种后出苗快而整齐。但浸种时间太短时,种子吸水不足,种胚内部酶类等大分子物质和细胞器活化较慢,影响种子萌发[8];浸种时间过长时,种子吸水过多,无氧呼吸严重,贮藏的可溶性养分外渗,易受病原菌感染,影响种子生活力[9]。实际上,不论浸种时间长短,总有部分西蒙得木种子萌发迟缓,导致发芽期延长,出苗不整齐,因此,种子萌发还依赖于其内部的生理变化。具有活力的种子胚乳或子叶中贮藏着大量营养物质,通过一定时间的休眠或解除休眠后给予适当的萌发条件,就能正常萌发成为植株,种子萌发所需要的养料与能量主要来自蛋白质、淀粉和脂肪等贮藏物质的转化与利用[10]。在浸种处理下西蒙得木种子贮藏物质的动员规律及其转化程度对种子萌发可能起到了关键作用,还需要更加深入的探讨。
傅里叶变换红外光谱具有鉴别物质可靠性强、分析快速、样品用量少等优点。从样品的红外光谱可知样品分子结构信息,从样品红外光谱中吸收峰的位置和强度,可知样品分子中可能含有的基团和基团数量,从而对样品组分进行定性和定量分析[11]。目前已应用于快速检测种子老化过程中或农作物种子发芽时水分、蛋白质、脂肪等物质的变化[12-14],本试验利用傅里叶变换红外光谱揭示不同浸种时间不同发芽阶段的西蒙得木种子贮藏物质动员情况,可为今后西蒙得木的引种繁育提供科学参考。
1 材料与方法
1.1 材 料
试验所用西蒙得木(SimmondsiachinensisLink)种子购自四川会东,为2019上半年所采种子,种子在试验前一直密封干燥低温保存。挑选完整饱满、大小均匀、健康的种子作为试验材料。
1.2 方 法
1.2.1西蒙得木种子吸水规律测定
随机抽取40粒西蒙得木种子,称其干重后,置于塑料盆(上口直径19 cm,高6.5 cm)中加蒸馏水1 L进行浸种,每隔2 h取出,用滤纸吸干表面水分,准确称量,连续测定36 h,设置3个重复。采用质量法测定种子吸水进程,计算种子的吸水率[15],并绘制种子吸水率曲线。
吸水率(%)=[(B-A)/A]×100%,式中:A为吸水前重量,B为吸水后重量。
1.2.2西蒙得木种子红外光谱测定
1) 浸种处理
挑选完整饱满、大小均匀、健康的种子360粒浸泡于0.5%(W∶V)KMnO4溶液30 min,取出用蒸馏水冲洗3次。取30粒消毒后的种子于塑料盆(上口直径19 cm,高6.5 cm)中,加蒸馏水1 L进行浸种。设置4个浸种时间,分别为4 h、10 h、16 h和24 h,每个处理3次重复。采用纸上发芽床法,将浸种后的种子均匀散放在垫有一张湿润发芽纸的发芽盆(32 cm×25 cm×11 cm)上,置于通风、无光照的30 ℃恒温培养箱中发芽,以种子着床当天为第1天,保持纸床水分充足3~7 d,每天更换发芽纸床,记录种子发芽情况。据前期预实验观察结果可知,相比短时间浸种,浸种10 h的种子发芽率更高,而浸种时间过长则容易污染,因此以浸种10 h的处理为标准,当其发芽率达到50%时终止所有发芽实验。种子萌发过程可分为吸胀、萌动、发芽3个阶段[16]。本研究从每个发芽盆中挑选出吸胀、萌动和萌发3个阶段的种子用于后续红外光谱测定,种子的吸胀程度与浸种时间有关;萌动种子表现为种皮开裂,露出胚根,即露白;萌发种子表现为胚根长度达到与种子长轴等长[17]。
2) 红外光谱分析
将每个浸种处理中吸胀、萌动、萌发的种子和未浸泡种子(ck)子叶取出,于鼓风干燥箱中60 ℃下烘干后粉碎,从每粒种子子叶碎屑中分别称取0.1 g样品按不同发芽阶段分别混匀,将混合样品于60 ℃下烘干至恒重,在红外灯下取少量样品与溴化钾(KBr)按1∶100的比例混合,于玛瑙研钵中充分研磨混匀,用YP-2压片机模具在25%相对湿度下压片,压力为15~20 MPa,持续压片20 s,压片至透明,厚度约为0.5 mm,采用PerkinElmer公司Frontier傅立叶变换红外光谱仪测定光谱特征(扫描范围4 000~400 cm-1,分辨率4 cm-1,扫描次数128次),所有光谱扣除溴化钾背景,最终得到样品的原始光谱图。为防止样品从环境中吸收其他物质(主要是水和二氧化碳),系统使用干氮进行净化。在相同条件下,每个样品做3次平行实验,取平均值并保存数据。
1.3 数据处理
使用红外光谱数据处理软件(OMNIC 8.0)对所采集的原始红外光谱图进行基线校正和自动平滑,利用Origin 8.5软件进行光谱数据处理,导出蛋白质、脂肪和碳水化合物相关特征峰的峰高及峰面积。表1列出供分析植物所含脂肪、蛋白质和碳水化合物的主要吸收带。其中,3 000~2 800 cm-1一般为脂肪酸甲基、亚甲基的基团振动吸收;1 670~1 500 cm-1一般为蛋白质中酰胺基团的振动;1 500~900 cm-1主要为多糖,也包含蛋白质和核酸的振动吸收[11,18-25]。

表1 生物体在4 000~400 cm-1范围内主要大分子特征性峰位及基团归属
红外光谱曲线是通过峰位和峰值(或峰面积)来确定样品内含物的种类和相对含量,不同光波上出现的峰值(或峰面积)代表不同化合物,而相同波数上峰值大小的不同则代表着相对含量的不同[26]。因红外吸收光谱的峰面积受样品因素和仪器因素的影响比峰高更小,所以使用吸收峰峰面积进行定量计算比使用吸收峰峰高更准确[27]。吸收峰面积通过对吸收峰进行积分计算可得,即将吸收峰波数范围内谱带上的数据点平均值乘以波数范围。谱带面积基本上不受谱带形状变化的影响,谱带面积与样品中基团总数成正比[11]。本研究选择峰面积相对比值来表征西蒙得木种子贮藏物质(脂肪、蛋白质和碳水化合物)的相对含量。
采用Microsoft Excel 2010软件进行数据的整理和作图,运用IBM SPSS Statistics 19.0软件进行数据统计分析。对实验数据进行双因素方差分析、多重比较,结果以平均值±标准差表示,p<0.05表示差异显著。用Pearson相关性分析贮藏物质之间及其与浸种时间、发芽阶段的关系。
2 结果与分析
2.1 西蒙得木种子吸水率变化
西蒙得木种子的吸水率随着浸种时间的延长呈抛物线型(图2),0~10 h的斜率最大,吸水速率最快,为急剧吸水期;10~24 h吸水速率逐渐变缓,为缓慢吸水期;24 h后吸水速率变化不明显,种子重量基本趋于恒定,进入吸水饱和期。由此可知,西蒙得木种子吸水吸胀主要集中在24 h内。

图2 西蒙得木种子吸水率变化趋势
2.2 原始红外光谱特征分析


图3 不同浸种时间不同发芽阶段西蒙得木种子的傅立叶变换红外光谱
脂肪特征峰2 926、2 855、1 465、1 167、1 740 cm-1吸收强,无干扰,选作脂肪分析峰,其峰面积范围为2 991.05~2 881.13 cm-1、2 879.20~2 819.42 cm-1、1 774.19~1 714.41 cm-1、1 482.99~1 427.07 cm-1、1 205.29~1 139.72 cm-1;蛋白质的酰胺Ⅰ 1 646 cm-1、酰胺Ⅱ 1 547 cm-1、酰胺Ⅲ 1 246 cm-1特征峰作为分析峰,其峰面积范围为1 714.63~1 492.62 cm-1和1 292.07~1 209.15 cm-1;1 075 cm-1特征峰作为碳水化合物分析峰,其峰面积范围1 141.65~931.449 cm-1[28]。计算样品脂肪特征峰面积、蛋白质特征峰面积、碳水化合物特征峰面积与特征峰面积之和的比值RF、RP、RC,分析西蒙得木种子贮藏物质的动员情况[28]。
2.3 不同浸种时间不同发芽阶段贮藏物质红外光谱分析
2.3.1RF的变化
双因素方差分析(表2)显示,浸种时间对RF有极显著影响(p=0.010),而吸胀和萌动的种子不同浸种时间之间差异不显著,因此显著影响来源于萌发阶段的种子。由图4 A可知,萌发阶段的种子RF随浸种时间总体呈逐渐增加的趋势,浸种16 h和24 h的萌发种子RF均显著高于对照组(未浸种)和浸种时间短(4 h)的种子,浸种24 h萌发种子的RF达到最高,吸胀的种子RF也有类似随浸种时间逐渐增加的趋势。不同发芽阶段种子的RF之间差异不显著(p=0.338),除4 h和24 h外,总体为随发芽进行RF呈现逐渐降低的趋势(图4 B)。浸种时间×发芽阶段交互效应对RF的影响不显著(p=0.220)。

注:同组小写字母相异表示差异显著(p<0.05),字母相同表示差异不显著(p>0.05)。下同。
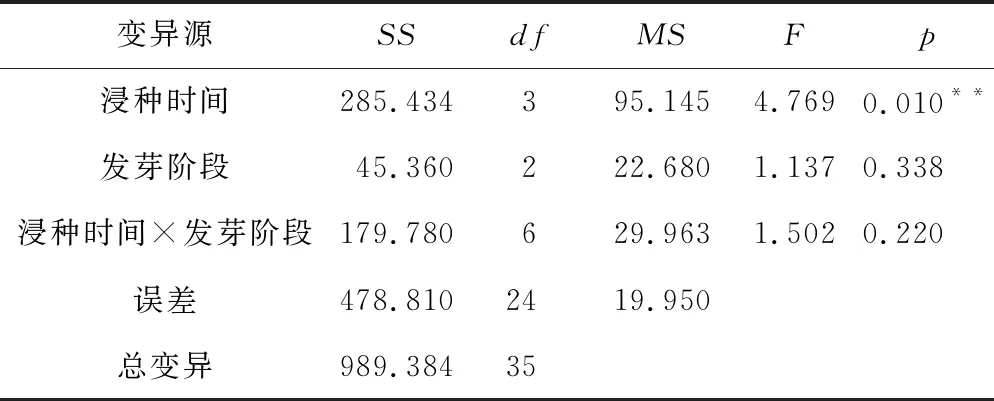
表2 不同浸种时间不同发芽阶段西蒙得木种子RF的双因素方差分析
2.3.2RP的变化
与RF类似,双因素方差分析(表3)显示,浸种时间对RP有显著影响(p=0.024),而吸胀和萌动的种子不同浸种时间之间差异不显著,因此显著影响来源于萌发阶段的种子。由图5 A可知,萌发阶段的种子RP随浸种时间总体呈逐渐降低的趋势,浸种16 h和24 h的萌发种子RP显著低于浸种4 h的,略低于ck组(未浸种)和浸种10 h的种子,但差异不明显,当浸种时间为24 h时,萌发种子的RP达到最低。不同发芽阶段种子的RP之间差异不显著(p=0.112)。浸种时间×发芽阶段交互效应对RP的影响也不显著(p=0.091)。

图5 不同浸种时间不同发芽阶段西蒙得木种子的RP(平均值±标准差,n=3)

表3 不同浸种时间不同发芽阶段西蒙得木种子RP的双因素方差分析
2.3.3RC的变化
双因素方差分析(表4)显示,浸种时间对RC有显著影响(p=0.029),而萌动和萌发的种子不同浸种时间之间差异不显著,显著影响来源于吸胀阶段的种子。由图6 A可知,萌发阶段的种子RC随浸种时间总体呈逐渐降低的趋势,浸种16 h和24 h的萌发种子RC低于对照组(未浸种)、浸种4 h和10 h的种子,但差异不明显,当浸种时间为24 h时,萌发种子的RC达到最低。不同发芽阶段种子的RC之间差异不显著(p=0.668),浸种时间×发芽阶段的交互效应对RC的影响也不显著(p=0.580)。

图6 不同浸种时间下不同发芽阶段西蒙得木种子的RC(平均值±标准差,n=3)

表4 不同浸种时间不同发芽阶段西蒙得木种子RC的双因素方差分析
2.4 贮藏物质之间及其与浸种时间、发芽阶段的关系
由表5可知,吸胀阶段的种子RP、RC与RF、浸种时间均呈显著负相关,RP与RC呈极显著正相关,RF与浸种时间呈极显著正相关;萌动阶段种子的RP、RC与RF均呈显著负相关,RP与RC呈极显著正相关,此时RP、RC与RF三者之间相关性达到最大,而与浸种时间的相关性均达到最低;萌发阶段种子的RP、RC与RF、浸种时间均呈显著负相关,RF与浸种时间呈极显著正相关。由表2~表4可知,发芽阶段对西蒙得木种子RF、RP、RC影响并不显著,而浸种时间对其影响显著。将RF、RP、RC与浸种时间进行相关性分析,可知种子RF与RP和RC呈极显著负相关,与浸种时间呈显著正相关;RP与RC呈极显著正相关;RC与浸种时间呈显著负相关。
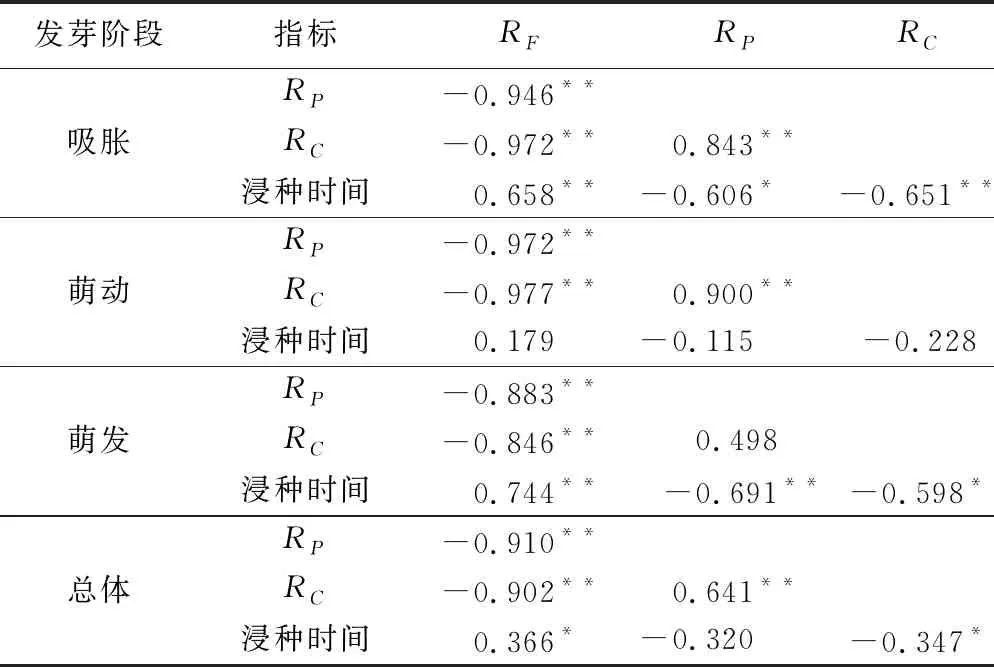
表5 不同发芽阶段西蒙得木种子的RF、RP、RC与浸种时间的相关分析
3 讨 论
种子萌发是植物生命周期的起始环节,种子自身特性与环境因子是影响种子萌发的主要因素。由吸水曲线可知西蒙得木种皮透水性良好,对种子吸水没有限制,外种皮不是阻碍其萌发的原因,与赵叶等[29]的研究结果一致。西蒙得木种子具有短时间内迅速吸水的特点,该特点与其长期适应沙漠环境而进化的生存策略有关,这与张卫华等[30]研究中沙芥种子吸水进程一致。西蒙得木种子吸胀前,内部大分子的淀粉粒和蛋白质等处于凝胶状态。吸胀时,种子膨胀、软化,水分子会迅速以扩散作用或毛细管作用等形式进入凝胶内部,具有极性的水分子与亲水凝胶结合起来,并使种子膨胀[31]。吸胀结束后,种子细胞原生质从凝胶状态转变为溶胶状态,各种酶开始活化,呼吸和代谢作用急剧增强[32]。与此同时,细胞迅速分裂和生长,吸水量迅速增加,胚开始生长[16],西蒙得木种子内贮藏的营养物质开始大量消耗。西蒙得木胚根突破种皮而外露,接着长出胚芽。
种子内贮藏的营养物质主要包括蛋白质、糖类和脂肪三大类,蛋白质可分解成小分子氨基酸,为种子萌发提供氮源,脂肪和糖类物质为种子解除休眠与萌发阶段的呼吸作用提供基础物质[33]。本研究中,西蒙得木种子贮藏物质在不同发芽阶段之间没有显著的差异,而主要影响来源于浸种时间,正如双因素方差分析(表2~表4)所示。多重比较进一步表明,萌发阶段的种子受浸种时间的影响显著。然而,Pearson相关性表明西蒙得木种子的RP和RC之间呈显著正相关关系,且均与RF表现出强烈且显著的负相关关系,本结果与张瑞等[34]的“芝麻种子萌发过程中总糖含量与粗脂肪含量呈极显著负相关”结论相似,说明本研究中贮藏物质之间是普遍存在的关系。RP和RC二者之间的正相关关系,蛋白质和碳水化合物在西蒙得木种子发芽过程中存在协同作用(图5 B和图6 B)。西蒙得木种子在萌发前期,水解蛋白酶活性逐渐增强,贮藏蛋白被水解为氨基酸等小分子,用于合成新蛋白质或转化合成糖类和脂肪[10],蛋白质相对含量降低;种子内的淀粉酶等水解酶会将自身贮藏淀粉等糖原分解为葡萄糖[17],呼吸增强,为种子生命活动供能,糖类的相对含量降低;西蒙得木种子在萌发初始阶段,子叶中的脂肪尚未动员或转化很少,胚根突破种皮后脂肪开始水解转化为碳水化合物(主要是糖),这同韩克杰等[35]在研究欧洲榛子时发现的种子胚乳中脂肪动员情况相似;随着浸种时间的增加,吸胀程度越高,蛋白质和碳水化合物的损失就越快,因此与浸种时间呈负相关关系,而随着蛋白质和碳水化合物的减少,脂肪相对累积,与浸种时间呈正相关关系。
4 结 论
1) 西蒙得木种子吸水吸胀主要集中在前24 h内,其中在0~10 h吸水速率最快,10~24 h吸水速率逐渐变缓,24 h以后达到饱和状态。
2) 西蒙得木种子贮藏物质在不同发芽阶段之间没有显著差异,主要受到浸种时间的影响,其中萌发阶段的种子受到浸种时间显著影响。
3) 蛋白质和碳水化合物在西蒙得木种子发芽过程中存在协同作用,随浸种时间延长而减少,脂肪随浸种时间延长而相对增加。
