玉米抗感粗缩病重要miRNA及其靶基因的诠释
2021-11-19吴斌崔汉青张眉邵春国姜珊珊郭霞徐德坤王升吉
吴斌 崔汉青 张眉 邵春国 姜珊珊 郭霞 徐德坤 王升吉

摘要:借助高通量测序和生物学分析手段,筛选不同抗性品种间的差异表达miRNA,预测相关靶基因。对玉米抗、感粗缩病不同种质材料高通量测序鉴定出的差异miRNA的靶基因,在Genome、Gene Ontology等生物信息数据库中进行比对,同时做GO(Gene Ontology,简称GO)、京都基因和基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,简称KEGG)富集性分析。从某些相关差异miRNA的靶基因及其作用位点的有关GO ID及其生物学功能注释中,发现一些可能与玉米粗缩病抗性有较大关系的miRNA,如zma-miR156a、zma-miR156k、zma-miR1432等;通过进一步的miRNA靶基因KEGG富集性分析,获得代谢通路ID,根据其生物功能注释,推测也有一定数量的miRNA与玉米种质抗(感)病相关。结合所涉差异miRNA在玉米抗、感种质接毒前后上、下调的表现,对这些重要miRNA靶基因可能涉及粗缩病的致病机制或玉米抗性相关的GO ID、KEGG通路ID所蕴含的功能进行了诠释。研究旨在探明抗病相关miRNA通过靶向作用其靶基因,在抗病过程中可能发挥的重要生物学功能和作用途径,研究结果可为探讨玉米抗粗缩病的抗病机制研究奠定理论基础。
关键词:玉米粗缩病;差异miRNA;高通量测序;靶基因;富集分析
中图分类号:S435.13 文献标志码:A
文章编号:1002-1302(2021)20-0064-06
收稿日期:2021-02-11
基金项目:山东省重点研发计划(编号:2018GSF121029);国家重点研发计划(编号:2018YFD0200603);山东省农业科学院农业科学创新工程(编号:CXGC2016B11-绿色防控)。
作者简介:吴 斌(1986—),男,山东临清人,博士,助理研究员,主要从事植物病毒学研究。E-mail:wubin228@126.com。
通信作者:王升吉,硕士,研究员,主要从事植物病毒学研究。E-mail:shjiw@163.com。
玉米粗缩病(maize rough dwarf virus,简称MRDV)是玉米生产中一类重要的病害,严重影响了玉米生产[1]。该病由带毒灰飞虱传播病毒,在山东省,其主要毒原为水稻黑条矮缩病病毒(rice black-streaked dwarf virus,简称RBSDV)[2-3]。前期研究发现,种植抗病品种是防治该病的重要措施,因此,发掘玉米粗缩病抗性相关基因、微小核糖核酸(miRNA)或反式作用干扰小RNA(ta-siRNA)等,分析其功能進而探究其抗病机制,可为抗病品种的遗传改良提供理论依据和技术支持。
miRNA是真核生物体内的一类负调控因子,在植物生长发育调节和病害防御中具有重要作用[4]。病毒在侵染寄主植物的过程中能够诱导大量miRNA产生,这些miRNA在病毒侵染时上调或下调表达,通过抑制防卫反应中的负调控因子和促进防卫反应中的正调控因子来行使功能。Wu等发现,水稻条纹病毒侵染水稻后,miR168与AGO18互作,调控水稻体内AGO1的表达,进而表现出抗病[5]。
本研究对RBSDV侵染不同抗性玉米品种所获叶片、茎秆组织进行miRNA高通量测序,对鉴定出的重要差异表达miRNA的靶基因开展GO、KEGG富集性分析,对其在玉米抗粗缩病中的生物学功能进行阐述和讨论,以期探究其在抗病过程中发挥的重要生物学功能和作用途径,进而为研究玉米抗粗缩病的抗病机制奠定理论基础。
1 材料与方法
1.1 试验材料
主要材料如下:玉米抗、感粗缩病品种分别为农大108、郑单958;感染水稻黑条矮缩病病毒(RBSDV)的水稻(或小麦)病株;人工养殖2~3龄的灰飞虱无毒若虫;小麦易感病品种;健康水稻幼苗。
1.2 试验方法
1.2.1 测序用玉米病株、健康株的获得 该工作是通过人工接RBSDV方法完成的,主要包括以下几个步骤:人工养殖继代灰飞虱;2个玉米品种的盆栽播种;2、3龄灰飞虱若虫从RBSDV病株上饲毒,并将度过循回期的带毒灰飞虱接于1~2叶期的玉米幼苗上传毒;将获毒后的玉米幼苗移栽于大田中并生长至喇叭口期,具体参见任春梅等的方法[6-7]。同期播种的玉米苗,保留足够数量不接毒的幼苗,肥、水等生长栽培管理同人工接毒苗。
1.2.2 miRNA高通量测序 待郑单958玉米接毒株充分发病后,依照测序样品分类信息(表1)进行样品采集,用液氮速冻、干冰保存等方式送至杭州联川生物技术股份有限公司进行miRNA的文库构建和高通量测序。
1.2.3 miRNA测序原始数据统计分析 测序获得原始序列(raw reads)后,首先去除3′端接头序列和垃圾序列,例如序列中有80%A/C/G/T、3N(不一定是连续的)序列,获得净化的序列(clean reads);然后进行长度筛选,对于植物,保留碱基为18~25 nt 的序列;接着将剩余的序列与mRNA、RFam(含rRNA、tRNA、snRNA等)和Repbase数据库进行比对,并进行过滤,获得有效序列(valid reads),去除重复序列,获得单一原始序列和单一有效序列;对miRNA的长度分布情况进行统计。
1.2.4 miRNA差异表达分析 统计测序结果中各样品miRNA的表达量,并进行2个品种相同组织在接种病毒、未接种病毒处理间的比对,筛选差异表达miRNA的标准为差异倍数|log2 (fold change)|>2、P<0.05。
1.2.5 差异表达miRNA靶基因的预测及富集分析 借助Target Finder对差异表达miRNA的靶基因进行预测,通过靶基因注释对玉米miRNA靶基因在生物学功能上行使的生物学作用进行分析。候选靶基因的注释借助于Gene Ontology(简称GO,http://www.geneontology.org/)和Genome中的KEGG(http://www.genome.jp/kegg/)数据库。
2 结果与分析
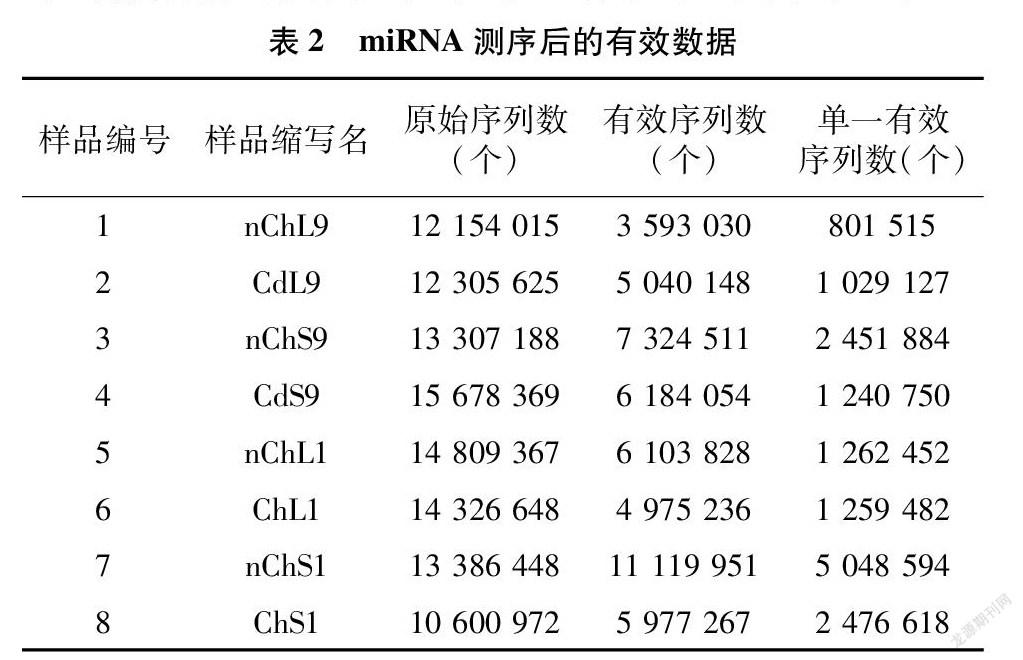
2.1 miRNA测序有效数据产出统计
待8个玉米miRNA文库通过高通量测序后,对每个文库内的全部小RNA片段进行测序,获得相应的原始序列数。然后进行长度筛选,过滤掉碱基长度小于18 nt、大于25 nt的序列,去除重复的miRNA序列,分别获得有效序列、单一有效序列等,详见表2。
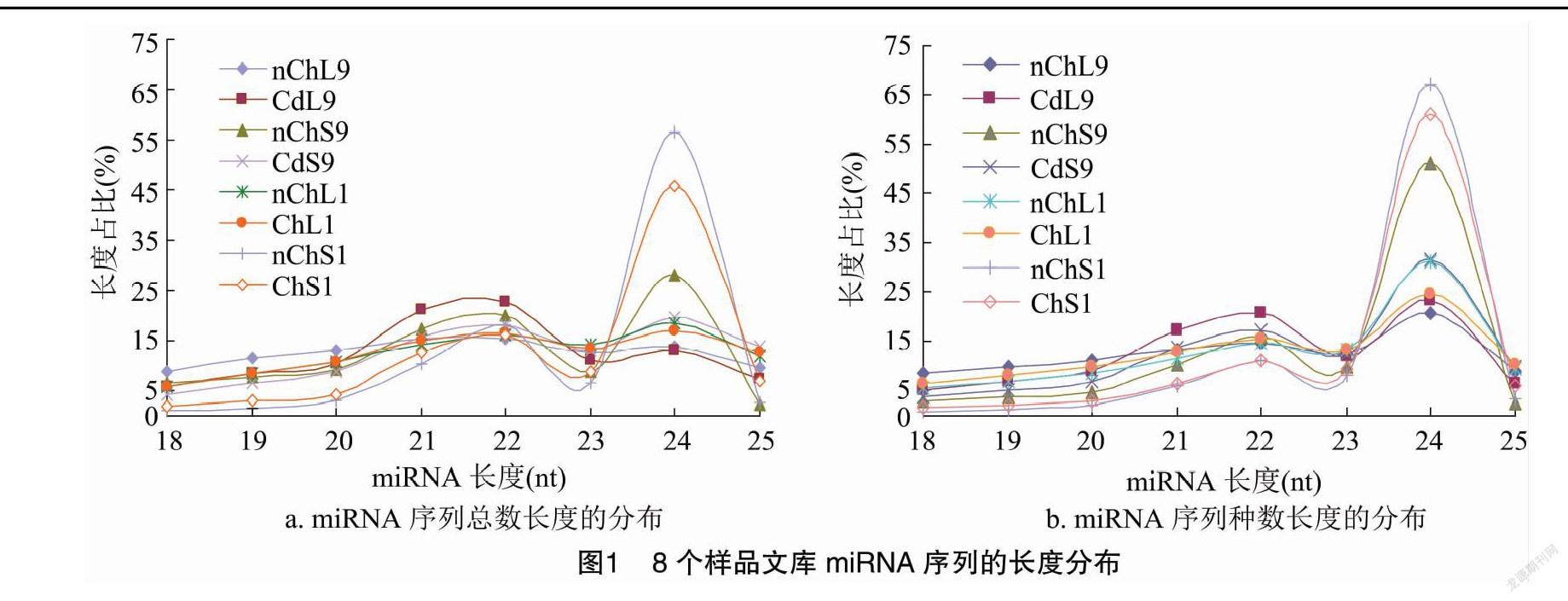
2.2 miRNA的长度分布
在对测序的原始数据进行分析统计的基础上,对8个样品文库中过滤去重后的有效(valid)序列总数(total)以及种数(unique)进行长度分布统计,发现其丰度主要集中于18~25 nt,且在24 nt处的序列丰度最大(图1)。
2.3 差异miRNA的上下调统计分析
通过对抗、感粗缩病2个品种的相同组织在接种病毒、未接种病毒处理间进行比对,并用TPM算法进行归一化处理,获得几个样品比较组有上调、下调差异性表达的miRNA情况。从图2可以看出,抗病品种农大108接毒后,健康植株茎秆、叶片材料较接毒前健康植株的比较组(ChL1 vs nChL1,ChL1 vs nChL1)的差异miRNA下调数明显较上调数多,而感病品种郑单958接毒后病株miRNA较接毒前健康植株(CdL9 vs nChL9,CdS9 vs nChS9)的比较组则相反,其差异miRNA的上调数较下调数多,尤其在叶片材料中,这可能是玉米抗病品种抗病机制的一种表现。
2.4 差异表达miRNA的筛选
通过对8个文库中的miRNA进行归一化分析,确认相关miRNA的表达差异。由表3可以看出,zma-miR156a-5p、zma-miR1432-5p、zma-miR390a-5p、zma-miR156k-5p、zma-miR396f-3p、zma-miR166a-3p、zma-miR164f-5p、zma-miR167e-5p、zma-miR394a-5p等9个miRNA在感病品种郑单958和耐病品种农大108的叶片、茎秆组织中接毒前后表达趋势相反,因此选择这9个miRNA作为重要miRNA展开研究。
2.5 差异表达miRNA靶基因的GO分析
对筛选出的9个差异miRNA的靶基因与Genome、Gene Ontology等生物信息数据库中的数据进行比对,进行GO富集性分析,发现差异miRNA的部分靶基因及其作用位点的生物学功能与玉米粗缩病抗性密切相关。其中,zma-miR156a-5p的部分靶基因与寄主对病原物的防御性反应相关,zma-miR1432-5p的部分靶基因与病毒的加工、装配有关,zma-miR390a-5p的部分靶基因与病毒粒子在寄主内细胞间的传输有关,其他miRNA的靶基因也多参与寄主的细胞信号传导、免疫应答的防卫性反应等(表4)。
2.6 差异表达miRNA靶基因KEGG分析
对筛选出的差异表达miRNA靶基因进行KEGG富集性分析发现,部分通路可能与玉米种质抗病(粗缩病)性相关。其中多个miRNA涉及的ko04626-植物病毒与植物寄主与病原间的相互作用、ko00130-泛醌及其他萜类醌的生物合成、ko04075-植物激素信号传导、ko00280-泛素介导的蛋白水解作用相关(表5)。对ko04626通路分析发现,靶基因通过调控RAR1、SGT1、HSP90等关键基因参与玉米与病原物的互作(图3)。
3 结论与讨论
当前对于玉米粗缩病的研究多聚焦于致病机制方面,对于其抗病機制的研究相对较少。水稻黑条矮缩病病毒通过细胞外壁的伤口或昆虫介体刺吸而造成微伤侵染玉米后,会引起寄主防御系统中相关基因表达、蛋白质合成、激素水平及生物途径的变化,使玉米生长受阻、发育不良,最终导致发病甚至死亡[8-10]。在此期间,玉米抗、耐病品种被植物病毒侵染诱导寄主产生大量的miRNA中,相关miRNA即可通过上调或者下调等模式,抑制防卫反应中的负调控因子或促进防卫反应中的正向调控因子,进而行使功能,可能是玉米抗耐病机制的一种表现[11]。本研究中涉及的zma-miR156、zma-miR1432、zma-miR390a-5p、zma-miR164f-5p等多个miRNA,均在受到RBSDV侵染胁迫后,在玉米抗、感病种质材料中呈现相反的表达趋势,且存在2种情况,部分在抗性材料(农大108)中表现为上调,在感病材料(郑单958)中表现为下调,另一种情况恰好相反,分析其原因,可能是miRNA的靶基因在抗病防御性反应中的正、负调控机制差异所致。
此外,在本研究中,抗耐病品种农大108接毒后,与接毒前相比,健康植株茎秆、叶片中差异miRNA下调数明显较上调数多,与感病品种郑单958在接毒后病株与接毒前健康植株比较的情况相反,可能是玉米抗、感品种分别受到粗缩病侵染胁迫时内源miRNA自身反馈调节的一种真实体现。农大108受到粗缩病病原侵染后,差异miRNA下调数多于上调数,可能由于miRNA所起作用的靶基因负调控作用机制相对较多,从而抑制了抗耐病品种植株发病,而感病品种郑单958受到病毒侵染后,差异miRNA的上调数多于下调数,植物病毒在与寄主的互作中占据优势,导致寄主发病。
另有研究者分别对水稻与玉米miRNA及其靶基因对玉米粗缩病病毒的应答通路进行了研究[12-14],结果显示,一些miRNA家族及其靶基因在玉米与病毒互作中起着重要作用[14]。例如,与泛素有关(miR166a-3p等)的靶基因的表达受抑制,泛素合成途径相关基因的表达发生变化,表明泛素途径在玉米抗粗缩病中起着重要作用。黄强的研究结果表明,RBSDV的侵染引起玉米中吲哚-3-乙酸(IAA)含量下降,玉米素核苷(ZR)相对保持不变,细胞分裂素与生长素的比值(C/A)升高,引起玉米顶端优势丧失,并导致细胞分化的紊乱,出现植株矮缩、脉突、心叶无法抽出等生长畸形症状,粗缩病症状迅速表现,导致植株畸形生长[9]。
此外,1种miRNA(例如miR156)可同时参与玉米的生长发育、信号传导、转录调控等多种功能,并在粗缩病胁迫响应过程发挥重要作用[15-16]。分析其原因可能是玉米粗缩病诱导玉米体内miRNA调控其靶基因参与信号传导、抗氧化胁迫、自身反馈调节、抗病相关代谢等途径[17-18],各类抗病途径共同构建了玉米抗粗缩病胁迫的复杂的防御机制。
参考文献:
[1]孙晓民,韩成卫,曾苏明,等. 玉米粗缩病发生规律与防控对策[J]. 吉林农业科学,2010,35(5):31-33.
[2]陈 佳,朱芹芹,袁从阳,等. 引起玉米粗缩病的水稻黑条矮缩病病毒山东分离物的分子特性[J]. 植物病理学报,2008,38(5):540-543.
[3]王升吉,张恒木,赵玖华,等. 山东玉米粗缩病原S10序列分析[J]. 玉米科学,2013,21(6):26-30.
[4]Carrington J C,Ambros V. Role of microRNAs in plant and animal development[J]. Science,2003,301(5631):336-338.
[5]Wu J G,Yang Z R,Wang Y,et al. Viral-inducible Argonaute18 confers broad-spectrum virus resistance in rice by sequestering a host microRNA[J]. Elife,2015,4:e05733.
[6]任春梅,楊 柳,刘保申,等. 一种高通量玉米粗缩病人工接种鉴定方法的建立及应用[J]. 玉米科学,2018,26(2):142-148.
[7]施 艳,韩晓玉,刘焱昆,等. 河南省玉米粗缩病抗病品种鉴定及病害与产量的相关性分析[J]. 河南科学,2020,38(2):208-211.
[8]Liu H J,Wei C H,Zhong Y W,et al. Rice black-streaked dwarf virus minor core protein P8 is a nuclear dimeric protein and represses transcription in tobacco protoplasts[J]. FEBS Letters,2007,581(13):2534-2540.
[9]黄 强. 玉米粗缩病症与内源激素水平变化的关系研究[D]. 成都:四川农业大学,2010.
[10]杨 青,郝俊杰,王新涛,等. 玉米粗缩病抗病基因定位和抗病育种研究进展[J]. 分子植物育种,2020,18(12):4021-4028.
[11]Peng D,Wu J G,Zhang J Y,et al. Viral infection induces expression of novel phased microRNAs from conserved cellular microRNA precursors[J]. PLoS Pathogens,2011,7(8):e1002176.
[12]Xu D L,Mou G P,Wang K,et al. MicroRNAs responding to southern rice black streaked dwarf virus infection and their target genes associated with symptom development in rice[J]. Virus Research,2014,190(9):60-68.
[13]李荣改,陆艳梅,王月影,等. 玉米粗缩病的分子研究新进展[J]. 植物学报,2017,52(3):375-387.
[14]Zhou Y,Xu Z N,Duan C X,et al. Dual transcriptome analysis reveals insights into the response to rice black-streaked dwarf virus in maize[J]. Journal of Experimental Botany,2016,67(15):4593-4609.
[15]雷凯键,刘 浩. 植物调控枢纽miR156及其靶基因SPL家族研究进展[J]. 生命的化学,2016,36(1):13-20.
[16]Wang J W,Czech B,Weigel D. MiR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana[J]. Cell,2009,138(4):738-749.
[17]朱早兵,虞夏清,翟于菲,等. 黄瓜microRNA171的克隆与功能分析[J]. 园艺学报,2019,46(5):864-876.
[18]Li Z X,Li S G,Zhang L F,et al. Over-expression of miR166a inhibits cotyledon formation in somatic embryos and promotes lateral root development in seedlings of Larix leptolepis[J]. Plant Cell,Tissue and Organ Culture,2016,127(2):461-473.
