长茎葡萄蕨藻在模拟工厂化循环水养殖环境中的脱氮研究
2021-11-13李华田道贺刘青松段亚飞张家松
李华, 田道贺, 刘青松, 段亚飞, 张家松
长茎葡萄蕨藻在模拟工厂化循环水养殖环境中的脱氮研究
李华, 田道贺, 刘青松, 段亚飞, 张家松*
中国水产科学研究院南海水产研究所, 农业农村部南海渔业资源开发利用重点实验室, 广东省渔业生态环境重点实验室, 广东广州 510300
为优化工厂化循环水养殖尾水处理效果, 探究长茎葡萄蕨藻在工厂化循环水养殖条件下对含氮污染物的去除效果。采用模拟养殖尾水, 考察了长茎葡萄蕨藻对氨氮、亚硝酸盐氮、硝酸盐氮的吸收效果, 进而讨论了不同水力停留时间对长茎葡萄蕨藻吸收去除三氮的影响。结果表明, 长茎葡萄蕨藻在单一含氮污染物中硝酸盐氮吸收速率最快; 在混合含氮污染物中, 氨氮吸收速率最快, 吸收去除效率最高, 其次为硝酸盐氮。长茎葡萄蕨藻在实验条件下养殖密度为16 g·L-1时生长较快, 氨氮、亚硝酸盐氮和硝酸盐氮的去除率为92.63%、61.91%和66.08%。在水流动状态下, 水力停留时间越短, 对三氮的综合去除效果较好, 水力停留时间为4 h时氨氮、亚硝酸盐氮、硝酸盐氮去除率为55.79%、56.37%、58.50%。
长茎葡萄蕨藻; 工厂化循环水养殖; 脱氮; 水力停留时间
0 前言
工厂化循环水养殖因具有节能、节水、节地、减排、安全、高效、不受季节限制等优点正处于快速发展阶段, 其核心技术是养殖尾水的净化循环利用[1-2]。如何除去水体中对养殖生物有害的氨氮和亚硝酸盐氮是养殖尾水脱氮、循环利用的关键。目前多数工厂化循环水养殖系统选择利用好氧硝化细菌将氨氮和亚硝酸盐氮转化成对养殖生物毒性较弱的硝酸盐氮[3-4], 因此在高密度循环水养殖系统中硝酸盐氮积累迅速, 其浓度能够达到200 mg·L-1或更高, 积累的高浓度硝酸盐一方面对养殖生物存在不利影响, 排放后会造成水域环境恶化, 不利于环境保护和水产养殖长期健康的发展[5]。
近年来, 利用大型海藻净化富营养化水体成为研究热点, 且有研究表明大型藻类在海水养殖尾水处理中能够发挥重要作用[6]。王翔宇等[7]比较了6种大型海藻对氮、磷营养盐吸收效果, 结果发现日本马泽藻和孔石莼在72 h时吸收了培养水体中98.0%和97.1%的氨氮、76.9%和82.0%的硝氮、90.8%和86.9%的活性磷, 另外4种蠕枝藻、红毛菜、扇形拟伊藻和鼠尾藻对营养盐的去除效果依次减弱。Luo等[8]在一定温度和光照强度下对比营养成分对石莼生长影响, 发现高浓度的硝态氮和铵态氮可促进石莼快速生长, 分别为13.1%·d-1、16.9%·d-1, 可有效消耗水体中营养盐, 净化废水。高桦楠等[9]在马氏珠母贝稚贝循环水养殖系统中放入针叶蕨藻、麒麟菜、长茎葡萄蕨藻混养, 养殖过程中可保证较好的水质状况, 稚贝存活率提高; 长茎葡萄蕨藻混养实验组对养殖水体的氨氮、硝酸盐氮处理效果最优。
以此可见, 在工厂化循环水水处理系统中引入大型海藻吸收养殖水体中的氨氮、亚硝酸盐氮和硝酸盐氮, 既能改善养殖水体水质, 又能增加养殖经济效益, 具有较好的应用前景[10-12]。长茎葡萄蕨藻()是一种可食用海藻, 又称为“海葡萄”或“绿色鱼子酱”[13], 隶属于绿藻门()、蕨藻科()、蕨藻属()。长茎葡萄蕨藻藻体含有粗多糖、粗纤维、蛋白质和维生素等多种营养成分[14-16], 具有抗癌、抗肿瘤、抗氧化、提高免疫、活化细胞等作用[17-18], 有较高的营养价值和经济价值。有报道长茎葡萄蕨藻对硝酸盐氮吸收净化效果较好, 其最适生长的硝酸盐氮浓度可达到15 mg·L-1, 活性磷酸盐浓度在0–5 mg·L-1[19]。
因此, 为提高工厂化循环水养殖系统对氨氮、亚硝酸盐氮和硝酸盐氮的去除效果。本研究选择长茎葡萄蕨藻为研究对象, 分析了其对氨氮、亚硝酸盐氮和硝酸盐氮的吸收速率; 在模拟养殖尾水中筛选出较优的养殖密度; 以及不同水力停留时间下长茎葡萄蕨藻对氨氮、亚硝酸盐氮和硝酸盐氮的去除效率。最终确定长茎葡萄蕨藻在工厂化循环水养殖系统中较优的运行参数, 以期为长茎葡萄蕨藻在工厂化循环水养殖系统中的深度脱氮提供基础数据和理论参考。
1 材料与方法
1.1 实验材料
实验开展于中国水产科学研究院南海水产研究所深圳实验基地6号车间, 车间顶棚半透光, 布有遮阳网, 控制自然光照强度。实验选用10 L收纳箱作为养殖容器, 所用长茎葡萄蕨藻为深圳大鹏湾野生长径葡萄蕨藻, 经选育选取质地较硬、颜色深绿的长茎葡萄蕨藻全株为实验材料。实验前将长茎葡萄蕨藻置于未投加营养盐的天然海水中进行暂养。实验配水所用试剂为氯化铵, 亚硝酸钠, 硝酸钠和磷酸二氢钾, 均为分析纯购自国药化学试剂有限公司。
1.2 实验方法
(1) 长茎葡萄蕨藻对单一含氮污染物的吸收速率
根据前期工厂化循环水养殖初期水质数据[20], 采用天然海水(未灭菌)分别配置含单一氨氮、亚硝酸盐氮和硝酸盐氮的养殖尾水。氨氮、亚硝酸盐氮、硝酸盐氮实测浓度分别为1.45 mg·L-1、0.96 mg·L-1、1.90 mg·L-1, 活性磷酸盐浓度均为0.10 mg·L-1。盐度为33, pH值为8.10±0.10, 水温约为25—28℃, 采用自然光照并进行遮光处理, 避免强光直射。实验组收纳箱中盛放5 L配置海水, 长茎葡萄蕨藻按照10 g·L-1的养殖密度静置于配置尾水中, 未进行曝气, 设置三个重复; 对照组中未放入长茎葡萄蕨藻, 其他条件与实验组一致。实验开始在第0、2、4、6、8、10、12、24 小时取水样过45 μm滤膜后分别测定氨氮、亚硝酸盐氮、硝酸盐氮浓度及活性磷酸盐浓度。
(2) 静态实验:
采用天然海水(未灭菌)配置含氨氮、亚硝酸盐氮和硝酸盐氮的养殖尾水。氨氮、亚硝酸盐氮、硝酸盐氮浓度分别为1.58 mg·L-1、0.58 mg·L-1、1.12 mg·L-1, 活性磷酸盐浓度为0.20 mg·L-1。收纳箱盛放5 L养殖尾水, 长茎葡萄蕨藻按照10 g·L-1、16 g·L-1、22 g·L-1的养殖密度静置于配置尾水中, 未进行曝气, 长茎葡萄蕨藻质量分别为50 g、80 g、110 g, 每个养殖密度设置三个重复。实验在自然光照下进行(遮光, 避免阳光直射), 水温在25℃—28℃范围内, 水体pH值在7.5—8.1之间; 对照组未放入长茎葡萄蕨藻, 其他条件与实验组一致。实验开始在第0、3、6、9、12、24、36、48、60、72 小时取水样过45 μm滤膜后分别测定氨氮、亚硝酸盐氮、硝酸盐氮浓度及活性磷酸盐浓度。
实验开始前和实验过程中, 取长茎葡萄蕨藻用吸水纸吸去长茎葡萄蕨藻表面水分, 用电子天平称取相应养殖密度的长茎葡萄蕨藻质量, 计算特定生长率; 计算长茎葡萄蕨藻对氨氮、亚硝酸盐氮、硝酸盐氮在0—12 h及整个实验过程中的吸收速率。
(3) 动态实验
根据静态实验结果, 选取较优长茎葡萄蕨藻养殖密度, 考察4 h、8 h和12 h三种不同水力停留时间对养殖尾水的净化效率。动态实验采用与静态实验相同模拟尾水浓度及养殖条件, 未进行曝气, 进水方式采用蠕动泵以一定流速将配置养殖尾水以左进右出、下进上出流经长茎葡萄蕨藻培养收纳箱。4 h水力停留时间每隔1 h从出水管中取水样, 8 h和12 h水力停留时间每间隔2 h取水样测定水质变化。水样过45 μm滤膜后测定氨氮、亚硝酸盐氮和硝酸盐氮浓度。
(4) 数据测定方法
光照强度采用标智BENETECH精度数字光照强度测定仪测定; 水温和pH值采用WTW (Multi 3620, Germany)检测, 氨氮(次溴酸盐氧化法)、亚硝酸盐氮(萘乙二胺分光光度法)、硝酸盐氮(锌镉还原法)和活性磷酸盐(抗坏血酸还原磷钼蓝法)均参照《海洋监测规范—海水分析》(GB 17378.4—2007)进行测定。
(5) 数据处理及统计方法
所用计算公式如下:
注: RSG表示特定生长率; W0、Wn分别表示实验前、后长茎葡萄蕨藻重量; t表示实验时间; P表示吸收速率; C0、Cn分别表示实验前、后养殖水体的营养盐浓度; V表示水体体积; m表示长茎葡萄蕨藻的养殖质量。
实验数据采用SPSS 22.0进行单因素方差分析, 如果差异显著(<0.05), 则用Duncan’s多重比较进行分析, 使用Origin 8.5进行图表处理。
2 结果
2.1 长茎葡萄蕨藻对不同含氮污染物的吸收速率
将长茎葡萄蕨藻放入单一含氮污染物的水体中发现, 氨氮、亚硝酸盐氮、硝酸盐氮均能够被长茎葡萄蕨藻吸收去除。实验组三氮变化曲线与对照组具有明显差别。如图1所示, 实验期间对照组氨氮、亚硝酸盐氮和硝酸盐氮浓度无明显变化, 实验组氨氮和硝酸盐氮浓度在24 h内下降明显, 去除率达到82.70%、90.73%, 亚硝酸盐氮去除率较低24 h去除率为23.98%。
其中, 0—12 h长茎葡萄蕨藻对氨氮和硝酸盐氮的吸收速率分别达到419.16 μg·(h·kg)-1、618.33 μg·(h·kg)-1; 亚硝酸盐氮吸收速率较低为44.17 μg·(h·kg)-1。12 h后光照减弱, 吸收速率减慢, 最终氨氮、亚硝酸盐氮和硝酸盐氮24 h平均吸收速率分别为250.42 μg·(h·kg)-1、47.92 μg·(h·kg)-1、358.13 μg·(h·kg)-1。
此外, 氮源种类不同, 长茎葡萄蕨藻对活性磷酸盐的吸收利用也不同。从表1中可以看出, 以氨氮为氮源时, 长茎葡萄蕨藻每吸收1.00 mg氨氮同时利用0.02 mg活性磷酸盐, 活性磷酸盐去除率为23.82%; 以亚硝酸盐氮为氮源时, 每吸收1.00 mg亚硝酸盐氮同时利用0.26 mg活性磷酸盐, 活性磷酸盐去除率为66.23%; 以硝酸盐氮为氮源时, 每吸收1.00 mg硝酸盐氮同时利用0.04 mg活性磷酸盐, 活性磷酸盐去除率为57.08%。

表1 长茎葡萄蕨藻24 h去除含氮污染物与消耗活性磷酸盐的关系
2.2 静态实验
2.2.1 水质变化
采用10 g·L-1、16 g·L-1、22 g·L-1三种长茎葡萄蕨藻养殖密度吸收去除同时含氨氮、亚硝酸盐氮和硝酸盐氮的模拟养殖尾水。实验发现三种含氮污染物同时存在时, 长茎葡萄蕨藻对含氮污染物的吸收顺序及去除率存在明显差异。从图2中可以看出, 长茎葡萄蕨藻对氨氮和硝酸盐氮在0—12 h去除较快, 亚硝酸盐氮和活性磷酸盐的吸收去除整体较平缓。此外, 吸收去除率均随养殖密度的增大逐渐增加, 但养殖密度增加对氨氮的促进吸收优势不明显。
此外, 实验发现在混合含氮污染物中, 长茎葡萄蕨藻会优先吸收去除氨氮, 0—12 h养殖密度10 g·L-1、16 g·L-1、22 g·L-1的长茎葡萄蕨藻对氨氮去除率即可达到62.13%、76.00%、80.52%, 吸收速率达到398.33 μg·(h·kg)-1、316.41 μg·(h·kg)-1、242.23 μg·(h·kg)-1。12—72 h氨氮浓度下降趋势减缓, 最终总去除率达到88.04%、92.63%、85.96%, 总平均吸收速率分别为94.16 μg·(h·kg)-1、64.24 μg·(h·kg)-1、43.24 μg·(h·kg)-1, 吸收速率与养殖密度负相关。
长茎葡萄蕨藻对亚硝酸盐氮吸收率较低, 去除速率较为稳定, 10 g·L-1、16 g·L-1、22 g·L-1三种长茎葡萄蕨藻养殖密度下亚硝酸盐氮的总去除率分别为25.33%、61.91%、54.61%。在0—12 h长茎葡萄蕨藻对亚硝酸盐氮吸收速率分别为15.00 μg·(h·kg)-1、21.61 μg·(h·kg)-1、24.62 μg·(h·kg)-1, 总平均吸收速率分别为10.07 μg·(h·kg)-1、15.58 μg·(h·kg)-1、9.88 μg·(h·kg)-1, 较氨氮吸收速率变化较小。
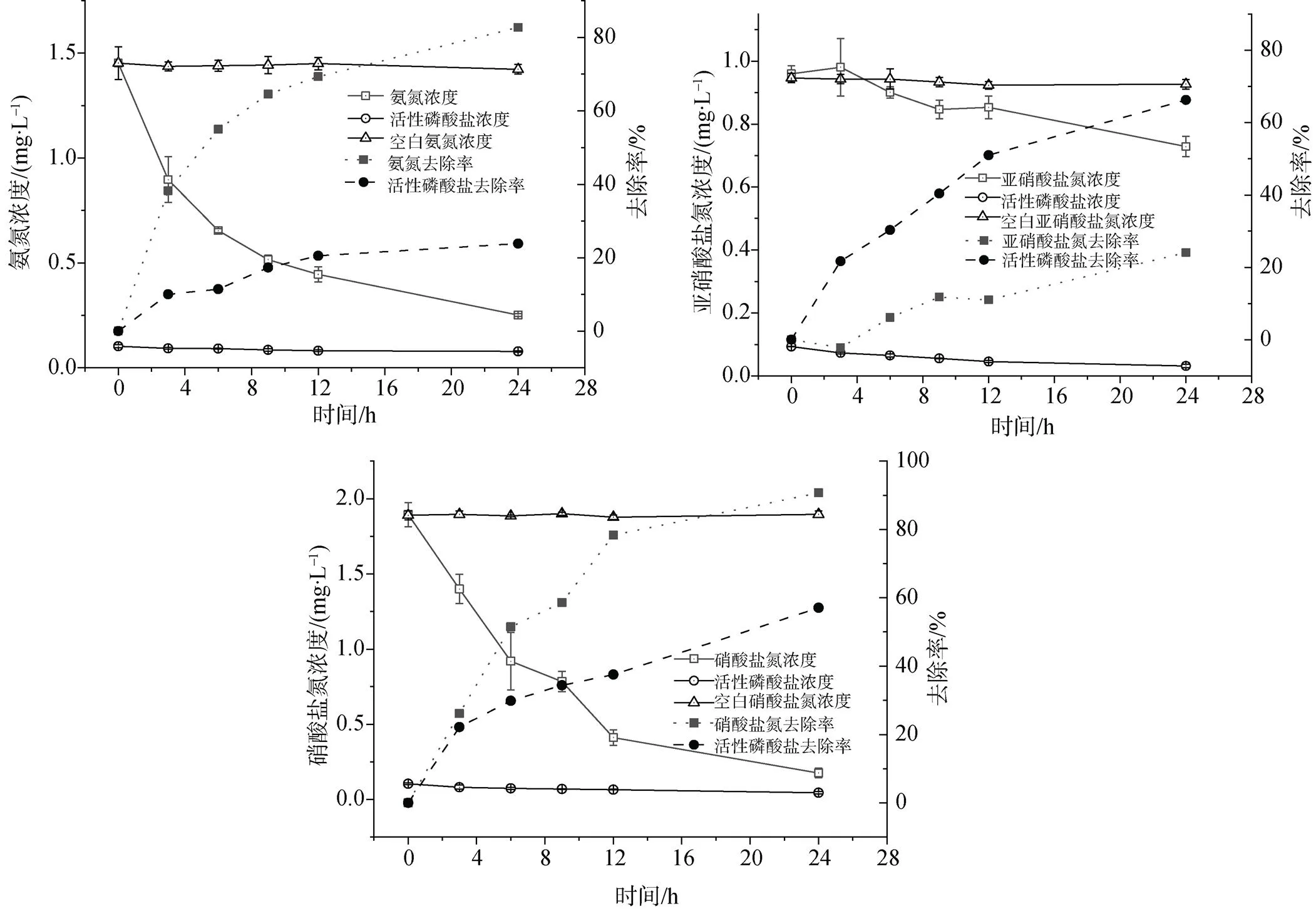
图1 含氮污染物、活性磷酸盐浓度变化及其去除率
Figure 1 Changes of nitrogen pollutants and active phosphate concentrations and their removal rates
硝酸盐氮浓度在实验期间也快速降低, 三种养殖密度下最终硝酸盐氮总去除率分别达到73.42%、66.08%、67.41%, 在0—12 h硝酸盐氮吸收速率分别为100 μg·(h·kg)-1、93.49 μg·(h·kg)-1、77.67 μg·(h·kg)-1。总平均吸收速率达到52.64 μg·(h·kg)-1、30.64 μg·(h·kg)-1、23.36 μg·(h·kg)-1, 略小于氨氮的吸收速率。
整个实验期间活性磷酸盐浓度平稳下降, 三种长茎葡萄蕨藻养殖密度下活性磷酸盐的浓度分别从0.19 mg·L-1、0.19 mg·L-1、0.21 mg·L-1降低至0.11 mg·L-1、0.08±0.01 mg·L-1、0.06 mg·L-1。总去除率达到42.11%、57.89%、71.43%, 平均吸收速率分别为6.11 μg·(h·kg)-1、4.69 μg·(h·kg)-1、4.77 μg·(h·kg)-1。鉴于长茎葡萄蕨藻在0—12 h对三氮的吸收速率较快, 后续实验设定的水力停留时间在0—12 h范围选取。
2.2.2 长茎葡萄蕨藻生长状况
不同密度的长茎葡萄蕨藻在实验期间生长状况不同, 如图3, 10 g·L-1与16 g·L-1养殖密度的长茎葡萄蕨藻重量不断增加, 22 g·L-1养殖密度的长茎葡萄蕨藻重量则持续降低。其中16 g·L-1养殖密度的长茎葡萄蕨藻实验期间重量增加6.83±0.45 g, 特定生长率最高; 10 g·L-1养殖密度长茎葡萄蕨藻增重4.37± 0.72 g; 而22 g·L-1养殖密度的长茎葡萄蕨藻在整个实验期间重量不断减少, 期间减重6.01±3.08 g, 这可能是由于养殖密度过高, 长茎葡萄蕨藻堆叠较厚, 底层藻体接收不到阳光照射逐渐发白、变软、腐烂造成藻体减产(如图4-a)。因此, 在设计长茎葡萄蕨藻养殖池的时候应选择高度较小, 面积较大的养殖池; 或者将藻种采用藻盘平铺可防止底部藻种接受不到阳光的现象。
图4为实验期间长茎葡萄蕨藻生长图片, 实验初期有部分长茎葡萄蕨藻部分出现白化、腐烂现象(图4-a)。在逐渐适应实验条件后进入快速生长阶段, 生长出大量纤细假根, 攀附在其他藻体或是培养箱璧上, 实现固着生长(图4-b)。其中, 单个直立茎藻体(无匍匐茎)在自身增长的同时, 会增殖生长出匍匐茎及假根用于固着攀附; 匍匐茎藻体除自身长度增加外, 分出较多数量的直立茎, 且数量和长度不断增加(如图4-c d)。

图2 不同密度长茎葡萄蕨藻对混合氮污染物及活性磷酸盐的吸收去除效果
Figure 2 Effects of different culture densities ofon nitrogen pollutants and active phosphate remove
综和水质数据和长茎葡萄蕨藻生长数据, 在实验条件下长茎葡萄蕨藻养殖密度为16 g·L-1生长速度最快, 氨氮、亚硝酸盐氮和硝酸盐氮的去除率可达到92.63%、61.91%和66.08%, 因此在后续动态实验中将采用该养殖密度进行实验。

图3 不同密度长茎葡萄蕨藻在混合氮污染物水体的特定生长率曲线
Figure 3 Curves of specific growth rate ofwith different densities in nitrogen pollutants water

注: 图b中标注为长茎葡萄蕨藻出现腐烂现象; 图c中标注为长茎葡萄蕨藻藻体出现发白现象; 图d中标注为新增直立茎。
Figure 4 Growth ofalgae
2.3 动态实验
如图5、6、7所示为养殖密度为16 g·L-1的长茎葡萄蕨藻在4 h、8 h和12 h水力停留时间对养殖尾水中三氮的吸收去除效果。在设定的水力停留时间内三氮的出水浓度随时间增加呈现先快速上升后趋于平缓的趋势。三个对照组在设定的水力停留时间最终出水浓度相差较小, 实验组中三氮出水浓度则在低于对照组的浓度范围缓慢增加。长茎葡萄蕨藻在一定水流条件下展现出对含氮污染物较好的吸收去除效果。
表2所示为不同水力停留时间下养殖尾水出水水质指标, 在4 h与8 h水力停留时间, 氨氮出水浓度相差较小(>0.05), 去除率分别为55.79%、53.15%(图5)。在12 h水力停留时间, 氨氮出水浓度较4 h和8 h降低明显(<0.05), 氨氮去除率达到65.84%, 较其他水力停留时间高, 停留时间长氨氮吸收去除效果较好。但随水力停留时间的增加, 亚硝酸盐氮出水浓度逐渐增加(<0.05), 从静态实验结果可知, 长茎葡萄蕨藻对亚硝酸盐氮吸收速率较低, 停留时间增加对亚硝酸盐的吸收去除未体现优势。在4 h和12 h水力停留时间下, 出水硝酸盐氮浓度差异不明显(>0.05), 这是因为长茎葡萄蕨藻对硝酸盐氮的吸收速率较稳定, 因此在实验设置的较低硝酸盐氮浓度条件下其对硝酸盐氮的去除率出现与水流速度无关的现象。
在水流速度一定的情况下, 水力停留时间越长, 所需体积越大, 占地面积越大。结合长茎葡萄蕨藻在不同水力停留时间对三种含氮污染物的去除效果, 可以得出在工厂化循环水养殖系统中养殖长茎葡萄蕨藻, 较短的水力停留时间, 可实现较好的含氮污染物去除效果。
3 讨论
3.1 单一含氮污染物的吸收去除
藻类的生长离不开营养盐的供给, 长茎葡萄蕨藻对不同形态的氮素均可以吸收利用去除, 吸收的亚硝酸盐氮、硝酸盐氮可经过还原酶作用可转化成氨态氮进行同化[21]。本研究中长茎葡萄蕨藻对单一含氮污染物的吸收速率为: 硝酸盐氮>氨氮>亚硝酸盐氮。根据研究报道[22-23]培养长茎葡萄蕨藻最适宜的氮源是硝酸盐氮, 且对硝酸盐氮的吸收大于氨态氮, 与本研究中出现长茎葡萄蕨藻对硝酸盐氮吸收速率较快的现象相吻合。

图5 不同水力停留时间对长茎葡萄蕨藻去除氨氮的影响
Figure 5 Effects of different hydraulic retention time on ammonia nitrogen removal of

图6 不同水力停留时间对长茎葡萄蕨藻去除亚硝酸盐氮的影响
Figure 6 Effects of different hydraulic retention times on nitrite nitrogen removal of
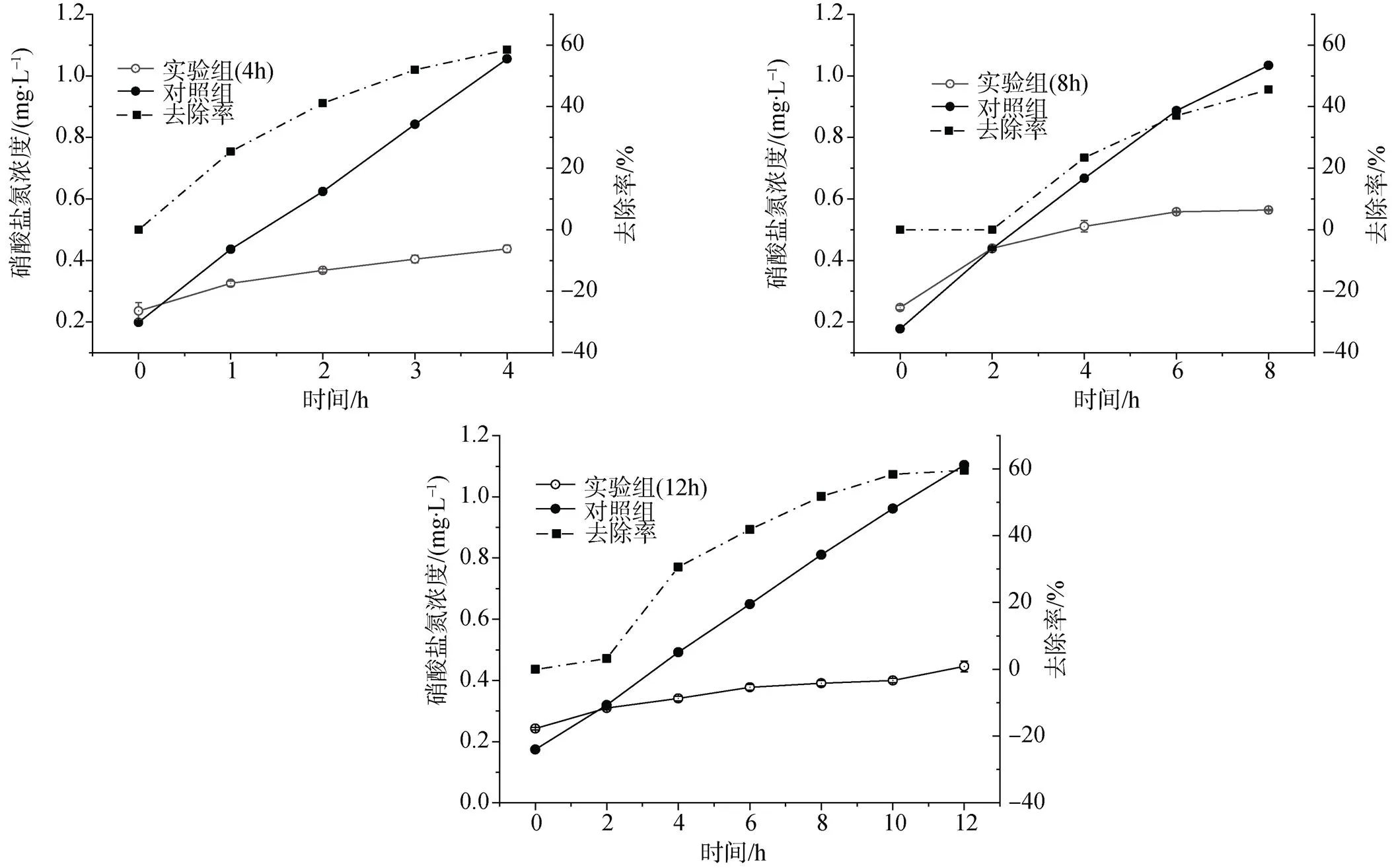
图7 不同水力停留时间对长茎葡萄蕨藻去除硝酸盐氮的影响
Figure 7 Effects of different hydraulic retention times on nitrate nitrogen removal of

表2 不同水力停留时间下出水水质指标
注: 同一列中的不同小写字母表示差异显著(<0.05)
长茎葡萄蕨藻吸收去除单位质量氨氮、硝酸盐氮消耗活性磷无明显差距, 但利用亚硝酸盐氮时, 吸收单位质量亚硝酸盐氮消耗活性磷盐的量分别是吸收单位质量氨氮和硝酸盐氮消耗活性磷酸盐的13倍和6.5倍。在现有关于大型藻类的文献中未发现对该现象的描述与解释。据陆生植物对氮磷的吸收特点, 猜测这可能与氮素中氮存在形态有关, 不同的氮素在吸收、运输、储藏和同化等代谢方面存在较大差异, 在很大程度上影响了长茎葡萄蕨藻的生长和对活性磷的吸收利用[24]。
3.2 混合含氮污染物的吸收去除
在含三种氮素污染物的模拟尾水中, 长茎葡萄蕨藻对不同含氮污染物的吸收速率为: 氨氮>硝酸盐氮>亚硝酸盐氮。有研究表明, 海藻对氨氮的吸收速率常数远大于硝酸态盐的, 藻类会优先利用污水中的氨氮和其它还原态的氮[25-26]。何洁等研究孔石莼、条浒苔()和巨角叉菜()吸收营养盐的动力学结果也表明氨态盐的被吸收率远高于硝酸态盐[27]。因此, 实验开始时处于“氮饥饿”状态的长茎葡萄蕨藻对氨氮有快速、大量吸收的现象。
对比单一含氮污染物中的吸收速率, 长茎葡萄蕨藻在混合含氮污染物中对3种氮素的吸收速率均较小。且出现对氨氮的吸收速率大于硝酸盐氮的吸收速率, 与在单一含氮污染物中的结果相反。Flynn[28]指出海藻对氨氮和硝酸盐氮吸收的相互作用复杂多变, 与生长率、氮源种类以及碳代谢都有关系, 主要可分为优先选择吸收氨态氮、氨态氮抑制硝酸态氮的吸收以及相互抑制等3种情况。在本研究中当模拟尾水中含三种氮素污染物时, 彼此间出现了相互抑制的现象, 造成了吸收速率小于在单一含氮污染物中的吸收速率, 且氨氮的吸收占优势。在工厂化循环水处理系统中, 将长茎葡萄蕨藻养殖池设置于生物过滤池后, 此时氨氮与亚硝酸盐氮已转化为硝酸盐氮, 可充分利用长茎葡萄蕨藻对硝酸盐氮的吸收速率优势实现养殖尾水的深度脱氮。
长茎葡萄蕨藻对三种含氮污染物的吸收速率与养殖密度成负相关, 这可能是由于长茎葡萄蕨藻养殖密度较大, 在有限的水体环境中, 分布较厚, 藻体的相互遮挡影响了它们对光的吸收, 光合作用率下降, 从而影响了对氮磷营养盐的吸收, 所以没有体现出养殖密度的影响优势[29]。由此可以看出,长茎葡萄蕨藻有最适养殖密度, 超过此养殖密度, 吸收速率不仅不会增大, 而且对长茎葡萄蕨藻的生长会产生负面影响, 较好的养殖方式是采用藻盘平铺避免藻种堆叠。
3.3 水力停留时间对三氮污染物的吸收去除
水交换率的控制在工厂化循环水系统养殖过程中是控制养殖成本的重要环节。水交换率过低造成有害物质的积累; 过高则能耗高, 降低养殖效益[30]。水力停留时间越长相对应水交换率越小, 水流越小, 实验中发现停留时间增加并未体现对含氮污染物的吸收去除优势。且综合对比, 长茎葡萄蕨藻在较短停留时间, 对氨氮、亚硝酸盐氮和硝酸盐氮具有较好的吸收去除性能。这主要是因为停留时间短, 水流速度增加能够减小藻体表面扩散边界层厚度从而加速养分和二氧化碳的供应与吸收, 促进其生长[31-32]。因此推测有可能出现更短的停留时间对三氮去除效率更高的现象, 但由于工厂化循环水处理系统生物滤池需要足够的停留时间来进行硝化反应, 且结合前期研究[20], 4 h停留时间既可保证微生物的处理效率, 长茎葡萄蕨藻对含氮污染物也能有较好吸收去除效率, 可实现养殖尾水的有效净化处理, 减少养殖水体氮污染物的积累。
4 结论
长茎葡萄蕨藻在实验条件下, 养殖密度为16 g·L-1时特定生长率高, 在水力停留时间为4 h其对氨氮、亚硝酸盐氮、硝酸盐氮去除率可达到55.79%、56.37%、58.50%。因此, 在工厂化循环水养殖系统中长茎葡萄蕨藻养殖池可设置在生物滤池后, 采用藻盘平铺养殖, 充分利用其对硝酸盐氮快速吸收的优势, 实现养殖尾水的稳定深度脱氮。
[1] BOVENDEUR J, EDING E H, HENKEN A M. Design and performance of a water recirculation system for high-density culture of the African catfish, Clarias gariepinus (Burchell 1822)[J]. Aquaculture, 1987, 63(1): 329–353.
[2] LAWSON T B. Fundamentals of aquacultural engineering[M]. Boston: Kluwer Academic Publishers, 1995.
[3] CHEN S, TIMMONS M B, ANESHANSLEY D J, et al. Suspended solids characteristics from recirculating aquacultural systems and design implications[J]. Aquaculture, 1993, 112(2–3): 143–155.
[4] PFEIFFER T J, OSBORN A, DAVIS M. Particle sieve analysis for determining solids removal efficiency of water treatment components in a recirculating aquaculture system[J]. Aquacultural Engineering, 2008, 39(1): 24–29.
[5] 宋协法, 杨晓晗, 黄志涛. 硝酸盐对鱼类毒性研究进展[J]. 中国海洋大学学报 (自然科学版), 2019, 49(9): 34–41.
[6] MCGLATHERY K J, PEDERSEN M F, BORUM J. Changes in intracellular nitrogen pools and feedback controls on nitrogen uptake in Chaetomopha linum (Chlorophyta)[J]. Journal of Phycology, 2010, 32(3): 393– 401.
[7] 王翔宇, 詹冬梅, 李美真, 等. 大型海藻吸收氮磷营养盐能力的初步研究[J]. 渔业科学进展, 2011, 32(4): 67–71.
[8] LUO Minbo, LIU Feng, XU Zhaoli. Growth and nutrient uptake capacity of two co-occurring species, Ulva prolifera and Ulva linza[J]. Aquatic Botany, 2012, 100: 0–24.
[9] 高桦楠, 李乐, 郑兴, 等. 循环水系统内马氏珠母贝与3种大型藻类的混养状况及水质分析[J]. 热带生物学报, 2016, 7(02): 147–152.
[10] XU Yongjian, FANG Jianguang, WEI Wei. Application of Gracilaria lichenoides (Rhodophyta) for alleviating excess nutrients in aquaculture[J]. Journal of Applied Phycology, 2008, 20(2): 199–203.
[11] 包杰, 田相利, 董双林, 等. 对虾、青蛤和江蓠混养的能量收支及转化效率研究[J]. 中国海洋大学学报(自然科学版), 2006, 36(S1): 27–32.
[12] 牛化欣, 马甡, 田相利, 等. 菊花心江蓠对中国明对虾养殖环境净化作用的研究[J]. 中国海洋大学学报(自然科学版), 2006, 36(s1): 45–48.
[13] KUDAKA JUN, ITOKAZU K TAIRA, et al. Investigation and Culture of Microbial Contaminants of Caulerpa lentillifera (Sea Grape)[J]. Journal of the Food Hygienic Society of Japan, 2008, 49(1): 11–15.
[14] MATANJUN P, MUHAMMAD K. Nutrient content of tropical edible seaweeds, Eucheuma cottonii, Caulerpa lentillifera and Sargassum polycystum[J]. Journal of Applied Phycology, 2009. 21(1): 75–80.
[15] SAITO H, XUE C, YAMASHIRO R, et al. High polyunsaturated fatty acid levels in two subtropical macroalgae, cladosiphon okamuranus and caulerpa lentillifera1[J]. Journal of Phycology, 2010, 46(4): 665– 673.
[16] 姜芳燕, 宋文明, 杨宁, 等. 海南长茎葡萄蕨藻的营养成分分析及评价[J]. 食品工业科技, 2014, 35(24): 356–359.
[17] MAEDA R , IDA T , IHARA H , et al. Immunostimulatory Activity of Polysaccharides Isolated from Caulerpa lentillifera on Macrophage Cells[J]. Bioscience Biotechnology and Biochemistry, 2012, 76(3): 501–505.
[18] BHESH, RAJ, SHARMA, et al. Anti-diabetic effects of Caulerpa lentillifera: stimulation of insulin secretion in pancreatic β-cells and enhancement of glucose uptake in adipocytes[J]. Asian Pacific Journal of Tropical Biomedicine, 2014, (7): 575–580.
[19] 黄建辉. 氮磷浓度及培养方式对长茎葡萄蕨藻(Caulerpalentillifera)生长的影响[J]. 渔业研究, 2012, 34(5): 416–419.
[20] 田道贺. 间歇式双循环工厂化养殖系统构建及应用[D]. 舟山: 浙江海洋大学, 2019.
[21] HOCKIN N L, MOCK T, MULHOLLAND F, et al. The response of diatom central carbon metabolism to nitrogen starvation is dillerent from that of green algae and higher plants[J]. Plant Physiology, 2012, 158(1): 299–312.
[22] 王海阳, 唐贤明, 金月梅, 等. 长茎葡萄蕨藻培养条件的研究[J]. 海洋湖沼通报, 2017, (6): 129–136.
[23] GUO Hui, YAO Jianting, SUN Zhongmin, et al. Effects of salinity and nutrients on the growth and chlorophyll fluorescence of Caulerpa lentillifera[J]. Chinese Journal of Oceanology & Limnology, 2015, 33(2): 410–418.
[24] 张平. 氮形态对低磷胁迫下苗期玉米磷素吸收的影响及其机理[D]. 南京: 南京农业大学, 2012.
[25] 程芬, 李乐, 殷小龙, 等. 3种海藻对无机盐和有机物水体中氨态盐净化效果比较[J]. 基因组学与应用生物学, 2017, 36(03): 1084–1089.
[26] XU GAO, HIKARU ENDO, YUKIO AGATSUMA. Seasonal changes in photosynthesis, growth, nitrogen accumulation, and salinity tolerance of Chaetomorpha crassa (Cladophorales, Chlorophyceae)[J]. Journal of Applied Phycology, 2018(1): 1–8.
[27] 何洁, 刘瑀, 张立勇,等. 三种大型海藻吸收营养盐的动力学研究[J]. 渔业现代化, 2010, 37(001):1-5..
[28] FLYNN K J. Algal carbon–nitrogen metabolism: a biochemical basis for modelling the interactions between nitrate and ammonium uptake[J]. Journal of Plankton Research, 1991, 13(2): 373–387.
[29] 王萍, 桂福坤, 吴常文. 营养盐因子对孔石莼和繁枝蜈蚣藻氮、磷吸收的影响[J]. 水产科学, 2010, 029(4): 208–211.
[30] DAVIDSON J, GOOD C, WELSH C, et al. The effects of ozone and water exchange rates on water quality and rainbow trout Oncorhynchus mykiss performance in replicated water recirculating systems[J]. Aquacultural Engineering, 2011, 44(3): 80–96
[31] 于琰, WERNBERG Thomas, DE BETTIGNIES Thibaut, 等. 室内三种富营养模拟条件下水流调控对羊栖菜和石莼生长及生物质合成的影响[J]. 浙江农业学报, 2018, 30(8): 1398–1407.
[32] HURD C L. Water motion, marine macroalgal physiology, and production[J]. Journal of Phycology, 2000, 36(3): 453–472.
Nitrogen removal from simulated recirculating aquaculture systemby
LI Hua, TIAN Daohe, LIU Qingsong, DUAN Yafei, ZHANG Jiasong*
Key Lab of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture; Guangdong Provincial Key Lab of Fishery Ecology Environment; South China Sea Fisheries Research Institute, Chinese Academy of Fishery Science, Guangzhou 510300, China
In order to optimize the water treatment effect of recirculating aquaculture system, the nitrogen removal ability ofin simulated recirculating aquaculture system was studied. In artificial aquaculture waste water, the removal ability of ammonia nitrogen, nitrite nitrogen and nitrate nitrogen bywas investigated. And then, the effect of different hydraulic retention time onnitrogen removal ability was examined. The results showed thathad fast absorption removal rate of nitrate nitrogen in waste water containing sole nitrogen pollutant, and had fast absorption removal rate of ammonia nitrogen in waste water containing three nitrogen pollutants, followed by nitrate nitrogen. Under experimental conditions, the culture density ofin 16 g·L-1grew faster and the removal rate of ammonia, nitrite and nitrate nitrogen could reach to 92.63%, 61.91% and 66.08%. In the flow condition,showed a good nitrogen removal ability at short hydraulic retention time. At 4 h hydraulic retention time, the removal rate of ammonia, nitrite and nitrate nitrogen could reach to 55.79%, 56.37% and 58.50%.
; recirculating aquaculture systems; nitrogen removal; hydraulic retention time
10.14108/j.cnki.1008-8873.2021.05.008
X52
A
1008-8873(2021)05-059-10
2020-04-09;
2020-05-25
广东省基础与应用基础研究基金区域联合基金青年基金(2020A1515110834);中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金资助(2021XK01,2021SD19);广东省现代农业产业技术体系创新团队项目(2021KJ150)
李华(1984—), 助理研究员, 主要从事养殖排放水处理技术研究, Email: babybluedaisy@163.com
通信作者:张家松(1971—), 研究员, 主要从事水产养殖研究, Email: jiasongzhang@hotmail.com
李华, 田道贺, 刘青松, 等. 长茎葡萄蕨藻在模拟工厂化循环水养殖环境中的脱氮研究[J]. 生态科学, 2021, 40(5): 59–68.
LI Hua, TIAN Daohe, LIU Qingsong, et al. Nitrogen removal from simulated recirculating aquaculture systemby[J]. Ecological Science, 2021, 40(5): 59–68.
