3种荒漠植物光合及叶绿素荧光对干旱胁迫的响应及抗旱性评价
2021-11-09王方琳柴成武唐卫东付贵全胥宝一
王方琳,柴成武,赵 鹏,唐卫东,付贵全,孙 涛,胥宝一
(1 甘肃省治沙研究所,兰州 730070;2 甘肃省荒漠化与风沙灾害防治国家重点实验室,甘肃武威 733000)
全球气候与环境变化,加上降水季节、地域分布不均匀,土壤有效含水量逐年减少,干旱已成为全球范围内影响植物个体生长、发育和繁殖的主要环境因素之一[1]。光合作用是干旱影响植物生长和代谢的第一环节[2]。荒漠区常年干旱使土壤水分严重亏缺,直接(光合原料减少) 或间接(气孔关闭、酶失活等) 地导致荒漠植物光合作用效率下降,成为植物生长的最大限制因素[3]。在长期的自然选择和协同进化过程中,荒漠植物在叶片光合结构方面形成了许多对水分亏缺的响应机制和适应策略。因此,在全球气候变暖及干旱胁迫逐渐加剧的形势下,开展荒漠植物抗旱模拟试验,分析其叶片光合作用及叶绿素荧光参数对干旱胁迫的响应特征,研究其抗旱机理,对生态环境建设及荒漠化防治过程中耐旱植物的选育具有重要意义。
梭梭(Haloxylonammodendron)、白刺(Nitrariatangutorum)、沙蒿(Artemisiadesertorum)是石羊河下游荒漠区主要固沙先锋植物,由于长期生长在长年干旱缺水、夏季炎热高温的恶劣环境中,3种植物在外部形态结构方面发生了一系列变化。梭梭叶片退化为同化枝进行光合作用,以减小受光面积降低蒸腾[4];白刺、沙蒿叶片形成浓密的表皮毛和较厚的角质层,对强烈的太阳辐射具有反射作用,以减少叶片吸收的光能,使叶肉组织免受高温灼伤;以上特征都是植物通过减少水分散失来抵御炎热,从而应对干旱环境的策略;同时这些植物在内部光合生理功能方面也发生了变化,深入了解其光合作用对土壤水分变化的响应具有重要意义[5-6]。
目前关于这3种植物光合生理方面的报道以对梭梭、白刺在不同逆境胁迫下光合作用及叶绿素荧光参数变化的研究较多。苏培玺等[7]测定了3种水分条件下梭梭的光合作用特征,并提出避免植物出现光抑制、提高光能利用率的有效方法;吉小敏等[8]采用小区梯度控水处理,比较4种灌水量对梭梭光合生理的影响;鞠强等[9]通过对干旱胁迫过程中梭梭光合日变化研究,提出其对干旱环境的适应主要是通过气孔的部分关闭实现的;赵长明等[10]通过对梭梭、白刺气体交换特征和叶绿素荧光参数日变化研究,得出白刺对荒漠生境的适应性比梭梭更强这一重要结论。以上研究对解释干旱胁迫条件下梭梭、白刺光合生理变化奠定了良好的基础,但这些研究多采用几项指标来反映基于少数几个土壤干旱胁迫梯度下两种植物的光合日变化特征,缺乏对土壤自然干旱过程中3种植物叶片光合色素、气体交换参数及叶绿素荧光参数响应特征方面的综合探究。而干旱环境抑制植物光合作用的机理比较复杂,涉及到诸多生理生化及生态因子。因此,必须在具体的试验条件下同时进行气体交换及叶绿素荧光参数测定,才可能对土壤干旱逆境下植物光合作用的抑制机理有更加深入系统、客观准确的认识[11]。此外,关于3种植物在干旱胁迫下光合作用参数的变化过程、机制及其与土壤水分的定量关系等诸多生理学问题还不清楚。
本试验参考吴芹等[12]的方法,采用盆栽控水试验结合土壤自然干旱处理获得多级胁迫梯度,研究梭梭、白刺和沙蒿3种荒漠植物光合色素、气体交换参数及叶绿素荧光光响应曲线对土壤自然干旱胁迫的响应特征;探讨3种植物光合作用对土壤干旱胁迫的响应机制及其与土壤水分的定量关系。为指导石羊河下游干旱荒漠区的物种选育提供参考。
1 材料和方法
1.1 试验材料
参试材料为2年生的梭梭、白刺、沙蒿盆栽实生苗。于2020年5月初选择大小一致的每种植物30株移栽至底径15 cm、口径21 cm、高33 cm的塑料花盆,每盆1株,栽培基质为取自民勤治沙综合试验站的纯沙土(模仿植物自然条件下的生长基质),栽植后常规管理。沙土的全钾、全磷、全氮、有机质和全盐含量分别为2.18%、0.028%、0.009%、0.314%和1.003%,pH为7.881,其黏粒(0.01~2 μm)、粉粒(2~20 μm)、细砂粒(20~200 μm)、粗砂粒(200~2 000 μm)分别占1.639%、4.239%、45.849%和48.271%。
1.2 试验设计
7月31日将花盆移入高2.5 m、上部采用透明聚乙烯材料遮盖的简易挡雨棚中,浇足一次透水后断水进行自然干旱胁迫试验。选取长势相近的每种植物5盆并挂牌标记,每隔6 d取样测定相关指标1次,试验共进行30 d,期间每天使用EM50水分探头每间隔1 h测量土壤含水率(测定结果为体积含水率)。
1.3 测定指标及方法
1.3.1 叶片水分和光合色素含量于上午8: 00~9: 00进行叶片取样,之后将材料立即带回实验室采用烘干法测定叶片相对含水量(RWC)及最大水分亏缺(RWD)。光合色素(叶绿素a、叶绿素b、类胡萝卜素)含量采用分光光度法测定,剪取3种植物的当年生向阳枝数个,蒸馏水清洗后用滤纸吸干表面水分,称取0.4~0.5 g鲜叶,剪碎后立即浸入20 mL内含丙酮、乙醇、水按45∶45∶10配成的混合液中,盖紧瓶盖并置于低温避光处浸提至叶片无色[13],每种植物设5次重复。
1.3.2 光合气体交换参数取每种植物当年生叶片,于上午9∶00~10∶00使用Li-6400光合仪(LI-COR,USA) 活体原位测定植物的光合气体交换参数,并将每种植物净光合速率(Pn)开始降为负值时的土壤含水率确定为该植物的最大水分阈值。采用Li-6400-02B 红蓝光源提供光合有效辐射强度(PAR,μmol·m-2·s-1),开放式气路,在PAR为1 200 μmol·m-2·s-1的光强下活体测定3种植物的叶片Pn(μmol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)、气孔导度(Gs,mmol·m-2·s-1) 和蒸腾速率(Tr,mmol·m-2·s-1),期间设定叶温为25 ℃,CO2浓度为380 μmol·mol-1,空气流速为400 μmol·s-1,每种植物随机选取5张叶片进行测定。最后依据测定参数分别计算3种植物的瞬时水分利用效率(WUE)、气孔限制值(Ls)。其中,WUE=Pn/Tr,Ls= 1-Ci/Ca,Ca为大气CO2浓度(400 μmol·mol-1)[14]。
1.3.3 叶绿素荧光光响应曲线叶绿素荧光光响应曲线采用CF-imager叶绿素荧光图像分析仪(Chlorophyll Fluorescence Imager,Technologica Ltd,UK)进行离体测定。剪取每种植物当年生向阳枝条2~3枝,放入保鲜箱后立即带回实验室,将叶片从基部剪切后依次摆放在湿润垫片上并固定,每种植物均选取5~6张叶片。依据实验目的运行预置程序,测定暗适应及不同稳态作用光强时的叶绿素荧光图像,并进行解析。具体测定程序如下:参照 Baker和Rosenqvist 的方法[15],3种植物的叶片经准确成像并聚焦后暗适应30 min,测定PSⅡ反应中心关闭时的最小荧光(Fo)和开放状态的最大荧光(Fm);开启光响应曲线测定程序,设光强梯度为0、25、50、100、150、200、300、500、800、1 200、1 500、1 800 μmol·m-2·s-1;根据基础荧光产率Fs的变化趋势,0~150 μmol·m-2·s-1低光强下平衡时间设3 min,光强大于200 μmol·m-2·s-1平衡时间设2 min;测定Fm和Fm′的饱和脉冲光为6 840 μmol·m-2·s-1。
其中,PSⅡ最大光化学效率(Fv/Fm)以准确暗适应30 min时的Fo和Fm计算,Fv=Fm-Fo;稳态作用光下PSⅡ有效光量子产量(Fv′/Fm′)依照Genty等[16]的公式计算,即Fv′/Fm′=Fm′-Fo′ /Fm′。PSⅡ非光化学猝灭系数(NPQ) 参照Baker和Rosenqvist 的公式计算:NPQ=Fm/Fm′-1[15]。以上参数中的Fo′以Oxborough和Baker 经验公式估算:Fo′ =Fo/(Fv/Fm)-Fo/Fm′,以上各项参数测定均进行5次重复[17]。测定期间室内气温(22.0±2.0)℃,空气湿度相对恒定。
1.4 数据处理
通过WPS及SPSS19.0进行试验数据处理及作图,试验结果采用 ANOVA 进行单因素(one-way ANOVA) 方差分析,利用Duncan法进行多重比较(α=0.05),图表中数据为平均值±标准差。应用模糊数学中的隶属函数法[18]对测定的16项指标进行计算,最后根据平均隶属函数值的大小综合评价3种植物的抗旱性。
计算公式为:R(Xi)=(Xi-Xmin)/(Xmax-Xmin)
若某一指标与耐旱能力呈负相关,则用反隶属函数公式计算:R(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)
式中:Xi为试验测定值;Xmin、Xmax分别为所有参试材料某一指标的最小值和最大值。
2 结果与分析
2.1 干旱胁迫期间3种荒漠植物土壤含水率和叶片水分含量的变化特征
首先,在自然干旱胁迫处理期间,梭梭、白刺、沙蒿3种荒漠植物的土壤含水率均随胁迫天数增加而逐渐降低,并且在胁迫0~18 d期间降低幅度较大,24~30 d间降幅较小,逐渐趋于稳定(图1,A)。其中,梭梭、白刺、沙蒿3种荒漠植物土壤含水率在干旱胁迫30 d时分别达到3.18%、3.04%和2.66%,分别比干旱胁迫前(0 d)降低了83.54%、84.08%和84.81%,即此时土壤含水率降幅以梭梭较小,沙蒿相对较大。
同时,3种荒漠植物叶片相对含水量(RWC)也均随干旱胁迫天数增加而逐渐降低(图1,B)。其中,在胁迫0~6 d期间,RWC以梭梭、沙蒿降幅较大并均达到显著水平,而白刺降低幅度较小,且白刺的RWC明显较高,而沙蒿明显较低;在干旱胁迫12~18 d期间,梭梭、沙蒿RWC趋于稳定,而白刺则显著降低,且降幅较大,但白刺的RWC仍明显高于梭梭、沙蒿;胁迫18~24 d期间,梭梭、白刺RWC均保持稳定,沙蒿RWC则显著降低并接近至最低水平;在胁迫30 d,梭梭、白刺RWC再次显著降低,且白刺的降幅更大,沙蒿RWC保持稳定,此时梭梭、白刺、沙蒿RWC均降至最小值,分别达到40.37%、25.49%和13.05%,它们比处理开始时(0 d)分别降低39.00%、63.73%和73.24%。干旱胁迫期间,白刺和梭梭叶片的RWC始终高于同期沙蒿;干旱胁迫结束时,3种植物叶片相对含水量及其降幅具有明显差异,梭梭RWC最高、变化幅度最小,沙蒿RWC最低、变化幅度最大。
另外,梭梭叶片相对水分亏缺(RWD)随胁迫天数增加而逐渐显著升高,并在胁迫12~18 d、24~30 d保持相对稳定,而白刺、沙蒿均呈“N”形波动式升高趋势,两者分别在6~12 d、6~18 d保持相对稳定;与处理0 d相比,3种植物RWD在胁迫30 d时分别显著升高46.31%、66.10%、42.81%(图1,C)。在土壤干旱胁迫期间,梭梭和白刺的RWD始终明显高于同期的沙蒿,两者最终升幅也高于沙蒿。
2.2 干旱胁迫对3种植物光合色素含量的影响
图2显示,整个干旱胁迫处理期间,梭梭叶绿素a(Chla)含量均无显著变化,各处理天数间无显著差异(P>0.05);白刺、沙蒿的Chla含量均随胁迫天数增加而降低,白刺的Chla含量在胁迫6 d时就显著降低(P<0.05),此后再无显著变化,沙蒿Chla含量在胁迫12 d时开始显著降低,直至胁迫24 d均一直无显著变化,随后在胁迫30 d时再次显著降低。与Chla含量不同,3种植物叶绿素b(Chlb)含量的变化不尽相同;梭梭Chlb含量随处理天数增加而显著降低,但在胁迫6~30 d间无显著降低;白刺、沙蒿Chlb含量呈“M”形波动式降低趋势,但白刺Chlb含量在胁迫6~24 d均显著上升,在胁迫30 d时又稍低于胁迫前水平,而沙蒿Chlb含量除胁迫12 d外均比胁迫前显著降低,在胁迫30 d时降至最低。梭梭、白刺、沙蒿总叶绿素(Chl)含量均随胁迫天数增加而降低,均在胁迫30 d时降低至最小值;梭梭和沙蒿Chl 含量在胁迫期间始终显著低于胁迫前,白刺Chl 含量仅在12和30 d比胁迫前显著降低,其余时间无显著变化。梭梭的类胡萝卜素含量随着胁迫时间先升后降,并在6 d时达到最大值,其在6~12 d时显著高于胁迫前,在24~30 d时显著低于胁迫前;白刺的类胡萝卜素含量随胁迫天数增加而逐渐降低,表现为0 d>6~12 d >18~24 d>30 d,沙蒿的类胡萝卜素含量的变化趋势与白刺相反,表现为0 d<6~12 d<18~24 d<30 d,且各时间段之间均有显著差异。

SS.梭梭;BC.白刺;SH.沙蒿;同一物种内不同小写字母表示处理时期之间差异显著(P<0.05); 下同图1 自然干旱胁迫过程中3种植物土壤含水率及叶片相对含水量和相对水分亏缺变化特征SS. Haloxylon ammodendron; BC. Nitraria tangutorum; SH. Artemisia desertorum; Different normal letters in the same species meant significant difference at 0.05 level. The same as belowFig.1 Changes of soil moisture content, leaf relative water content and leaf relative water deficit of three species during drought stress
2.3 干旱胁迫对3种植物气体交换参数及水分利用效率的影响
表1显示,梭梭叶片净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE) 均随干旱胁迫天数增加呈先升高后降低的变化趋势;梭梭叶片Pn、Tr均在干旱胁迫12 d(土壤含水率为8.04%)时达到最大值,WUE则在干旱胁迫6 d(土壤含水率为15.25%)时达到最大值,且均与其他处理时间存在显著差异(P<0.05)。说明过高或过低的土壤水分均会引起梭梭叶片光合作用水平的降低,维持叶片较高水平Pn、Tr的土壤含水率范围为8.04%~19.33%,维持较高水平WUE的土壤含水率范围为15.25%~19.33%。同时,梭梭气孔导度(Gs)和气孔限制值(Ls)均随胁迫天数增加而降低,胞间二氧化碳浓度(Ci)则呈逐渐升高趋势,干旱胁迫6 d时就均与处理前差异显著;在干旱胁迫24 d(土壤含水率为3.49%) 时,梭梭Ls降至0.0169,同时也伴随着Pn剧烈降低(0.7624),说明此时植物气孔基本上失去了调节作用,Pn下降的主要原因是发生了由气孔因素向非气孔因素的转变。
白刺气体交换参数变化特征与梭梭略有差异。随着干旱胁迫天数增加,白刺叶片Pn、Gs、Tr均呈逐渐降低的变化趋势,Pn、Gs在干旱6 d以及Tr在干旱12 d时降幅就达到显著水平,Pn在干旱30 d时降低为负值。同时,WUE和Ls均随胁迫天数增加呈先缓慢升高然后剧烈降低的趋势,其Ci则呈先缓慢降低然后剧烈升高的趋势,干旱处理12 d(土壤含水率为8.37%)时,前两者均达到最大值,而后者降至最低值,说明维持白刺WUE处于较高水平的土壤含水率范围为4.17%~19.10%。在干旱30 d时(土壤含水率为3.04%),WUE和Ls均降至最低值,而Ci则达到最大值,Pn也降低为负值,说明此时白刺叶片气孔已失去调节作用。
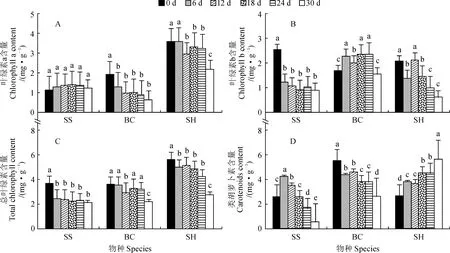
图2 干旱胁迫过程中3种植物光合色素含量变化特征Fig.2 Changes of photosynthetic pigment contents of three species during drought stress
沙蒿气体交换参数变化特征与白刺相似,除Ci随胁迫天数增加呈现稳定升高外,其他各参数均随胁迫天数增加而逐渐降低,并均在干旱6 d或者12 d时升降幅度就达到显著水平,说明土壤含水率变化容易直接引起沙蒿光合作用水平的降低。在干旱胁迫18 d时,沙蒿叶片Pn降至0.056 2,同时Gs、Tr,、WUE也相应降至较低的拐点,Ls已为负值。以上结果说明,干旱胁迫0 d~12 d(土壤含水率为6.48%~17.51%)时,沙蒿可正常生长;当胁迫18 d时,土壤含水率(4.14%)已达到影响其正常生长的水分阈值;胁迫24 d~30 d(土壤含水率为2.66%~3.07%) 时,Pn、WUE等参数均为负值,此时沙蒿叶片光合作用能力完全丧失,植物发生生理性死亡。
2.4 干旱胁迫对3种植物叶绿素荧光光响应曲线的影响
从图3可知,3种荒漠植物叶片PSⅡ最大光化学效率(Fv/Fm)均随光合有效辐射的增强而逐渐降低;干旱胁迫30 d(土壤含水率为3.04%)时,梭梭和白刺的Fv/Fm均降至较低值,此时相同光强下各处理天数间差异不显著(P>0.05);沙蒿Fv/Fm在干旱胁迫0 d~18 d各胁迫天数间无显著差异(P>0.05),在干旱胁迫24 d~30 d(土壤含水率为2.66%~3.07%)时降至最低。
同时,3种植物PSⅡ有效光量子产量(Fv′/Fm′)均在光强小于300 μmol·m-2·s-1时降低幅度均较大,在光强大于300 μmol·m-2·s-1时基本维持稳定;在中、高光强时,白刺Fv′/Fm′ 均保持在0.4~0.6之间,胁迫24~30 d时略有降低;梭梭与沙蒿Fv′/Fm′变化趋势相似,在胁迫30 d时均降为0.2左右,并与其他处理天数间差异显著(P<0.05)。
此外,3种植物qP在各胁迫天数处理时均随光强增大而逐渐降低。其中,梭梭qP值随光强增加呈明显的“哑铃”形变化,即在低光强和高光强时各胁迫天数间变化无显著差异(P>0.05),在光强为500~1 200 μmol·m-2·s-1时差异较大;梭梭qP在胁迫18 d时达到最大值,而在胁迫0 d和30 d时最小。沙蒿qP值随光强变化也接近“哑铃”形,在胁迫6 d~18 d时具有较大值,在胁迫0 d、24 d、30 d时具有较小值。白刺qP值在各光强下表现出相似的趋势,胁迫30 d时始终最高,胁迫0 d和6 d时始终最低,其余胁迫时间居中。

表1 干旱胁迫过程中3种植物气体交换参数的变化特征

图3 干旱胁迫过程中3种植物叶绿素荧光光响应曲线变化特征Fig.3 Characteristics of chlorophyll fluorescence response curves of three species under drought stress
另外,3种植物NPQ在各干旱胁迫时间下均随光强增大而逐渐升高,且在光强0~500 μmol·m-2·s-1时变化剧烈,而在500~1 800 μmol·m-2·s-1时较为稳定。其中,白刺NPQ分别在胁迫18 d和0 d时分别具有最大值和最小值,其他各处理天数间差异不显著(P>0.05)。梭梭、沙蒿NPQ均在处理0 d、光强500~1 800 μmol·m-2·s-1时具有最大值;胁迫6 d~24 d时,梭梭和沙蒿NPQ均显著降低,但梭梭NPQ在各处理间差异不显著(P>0.05),沙蒿NPQ各处理天数间差异明显,表现为6 d>12 d>18 d、24 d;胁迫30 d时,两植物NPQ在各光强下均明显低于其余处理天数。两植物NPQ在胁迫30 d时各光强下均接近0,其他处理天数在光强500~1 800 μmol·m-2·s-1范围内处于0.5~1.5之间。沙蒿、梭梭NPQ变化趋势与处理天数间明显相关,即存在明显的水分效应;在胁迫18 d~30 d时NPQ降至0~0.5之间,并与其他胁迫天数间存在显著差异(P<0.05)。

表2 干旱胁迫下3种荒漠植物各测定指标的隶属函数值
2.5 3种植物抗旱性综合评价
模糊隶属函数法是一种通过多种指标计算进行植物抗旱性综合评价的有效方法,其平均值越大抗旱性越强,这种方法克服了采用单一指标评价植物耐旱能力的片面性[19]。本研究应用叶片水分、光合及荧光参数共16种指标对3种植物抗旱性进行综合评价(表2),结果表明,3种植物各项指标隶属函数的平均值分别为0.501 2、0.548 9、0.492 9,说明其耐旱能力由高到低依次为白刺、梭梭、沙蒿。
3 讨 论
干旱胁迫引起植物水分亏缺,并通过抑制叶片伸展、影响叶绿体光化学及生物化学活性等途径,延缓或破坏植物正常生长,加快植物个体组织、器官衰老、脱落或导致其死亡[20]。叶片形态的变化是植物受到胁迫时的最初体现,植物叶片相对含水量(RWC)和相对水分亏缺(RWD)是植物体内生理功能旺盛的标志,其值越大说明植物对干旱的适应能力越强[21]。本研究中白刺、沙蒿RWC随干旱胁迫天数增加持续降低,而梭梭RWC在胁迫0 d~18 d时仍较为稳定,植物体仍然能够调节自身体内的水分平衡,说明土壤含水率为5.05%时未达到引起梭梭叶片水分降低的阈值;胁迫24 d~30 d(土壤含水率为3.18%~3.49%)时梭梭叶片RWC显著降低。植物叶片RWD的变化特征也能证明以上观点,干旱胁迫后期,白刺和沙蒿RWD迅速升高,说明这两种植物叶片水分状况对外界环境条件变化较为敏感;可能是由于两植物叶面积较大,干旱胁迫过程中强烈的蒸腾作用导致植物体水分严重缺失,同样也加剧了土壤含水率的快速下降。
光合色素是植物进行光合作用的基础,干旱胁迫导致叶绿体膜系统的紊乱或叶绿体结构整体性的破坏,从而影响光合色素的合成[22-23]。研究表明,光合色素含量的高低并不能完全反映叶片光合能力的强弱,但其含量减少直接导致叶片衰老和光合速率的不可逆降低[24]。本研究中,梭梭Chla、Chlb及Chl含量只在胁迫0 d~6 d时显著降低,之后保持稳定,可能是由于胁迫初期土壤含水率的迅速降低,促使其叶绿素降解酶大量合成,而之后持续干旱胁迫时,3种光合色素反应较不敏感。白刺Chla、Chlb、Chl及胡萝卜素含量均在胁迫24 d~30 d时显著降低,说明3.04%~3.69%的土壤含水率已对其叶绿体细胞的整体性造成极大破坏,使叶肉细胞光合活性和吸光强度降低,从而影响叶片的光合作用;沙蒿光合色素含量在胁迫18 d~30 d、土壤含水率为2.66%~4.14%时急剧降低,但其Chla和Chl含量均显著高于其他两种植物,这与师生波等[25]对民勤沙生植物园4种云杉属(Picea)植物的研究结果类似,可能是沙蒿在相同生长期间维系自身枝干较高生长所致。此外,梭梭、白刺类胡萝卜素含量随干旱胁迫天数增加而降低,而沙蒿则呈逐渐上升趋势,这种现象可能与野外深秋时节观测到的幼苗叶色由嫩绿变为紫红的现象有关。
光合气体交换参数也受到干旱胁迫程度和时间的影响。土壤干旱胁迫下植物光合速率的下降通常可分为气孔因素和非气孔因素,前者是指干旱胁迫导致气孔导度下降,CO2的进入受到阻碍从而使光合速率下降,后者是指植物叶肉细胞的光合活性下降;干旱胁迫过程中,大部分植物能通过合理协调碳同化和水分消耗之间的关系,调节叶片WUE的变化,这是植物抗旱策略的重要组成部分[10]。本研究中3种植物Pn、Tr、WUE对土壤水分都表现出明显的阈值响应,梭梭WUE达到最大值时的土壤含水率 (15.25%)明显大于其Pn达到最大值时的土壤含水率(8.04%),即梭梭叶片WUE在水分胁迫会降低,这与赵长明等[10]的研究结果一致;相反,白刺WUE达到最大值时的土壤含水率 (8.37%) 则明显小于其Pn达到最大值时的土壤含水率(19.10%),说明白刺的WUE在适宜的干旱胁迫则会显著提高,这与裴斌等[26]对沙棘(Hippophaerhamnoides)的研究结果相一致;沙蒿WUE和Pn达到最大值的土壤含水率相同(17.51%),其Pn和WUE 均随土壤含水率降低而持续下降,甚至在土壤含水率低于4.17%时均降低为负值。可见,3种植物中沙蒿所受干旱胁迫的影响最大。干旱胁迫过程中梭梭、白刺、沙蒿3种植物维持正常光合生理活动适宜的土壤含水率阈值范围分别为8.04%~19.33%、4.17%~19.10%、6.48%~17.51%,以上阈值能保证植物叶片同时获得较大的Pn和WUE,这种协调作用符合荒漠植物生长的“高效水阈值”或“经济水阈值”,即以高效用水为核心的栽植管理原则[27-28]。
叶绿素荧光是研究光合作用机制和探测光合生理状况的一种新兴技术,与反映“表观性”的气体交换指标相比,具有“内在性”的特点,是研究植物光合生理与逆境胁迫的内在探针[25]。叶绿素荧光参数的光响应曲线可快速反映植物光合功能对环境光强变化的应答[29]。本研究中3种植物PSⅡ最大光化学效率(Fv/Fm)的光响应曲线具有非常相似的变化趋势,可能是植物光合机构对民勤干旱荒漠气候长期适应的表现。植物Fv/Fm未受到胁迫时一般在0.80~0.85之间,遭受干旱、强光等逆境胁迫时变小[30-31]。本试验中各胁迫天数处理时,3种植物Fv/Fm均随光强增大而降低,说明干旱胁迫对3种植物PSⅡ反应中心活性均有影响;光强低于500 μmol·m-2·s-1时,梭梭、白刺的Fv/Fm值均较大,但随着光强增大而大幅度降低,以胁迫24 d和30 d时降低较为明显。这说明较长时间的干旱胁迫对沙蒿产生光抑制,可降低或破坏其光合活性;梭梭、白刺Fv/Fm受干旱胁迫的影响较小,胁迫天数增加时均可得到一定程度的恢复。
PSⅡ有效量子产量(Fv′/Fm′)反映实际光能转换效率,随光强增大而减小[31]。本研究除梭梭、白刺胁迫30 d的时段外,其他胁迫天数时3种植物Fv′/Fm′均在光强0~300 μmol·m-2·s-1时较大幅度降低,之后随光强增加趋于稳定状态;整个胁迫过程中白刺Fv′/Fm′值均大于等于0.4,而梭梭、沙蒿均在胁迫30 d时降至0.2左右。说明土壤水分条件较好时,3种植物均可通过自身调节机制平衡叶片对光能的捕获、利用与耗散之间的关系,保持叶片PSⅡ反应中心活性,而土壤含水率的降低特别加剧了沙蒿Fv′/Fm′下降趋势。相比较而言,白刺通过调节机制耗散掉过剩的光能使自身PSⅡ反应中心免受伤害的能力较强。
光化学淬灭系数(qP) 反映了PSⅡ反应中心的开放程度,其值越大说明电子传递活性越高[32]。干旱胁迫时, 植物体内基质类囊体膜上PSⅡ反应中心多呈失活状态,并与光捕获天线色素一起参与对过剩激发能的耗散,从而保护有功能的反应中心免受多余光能伤害[21]。本研究中,当光强小于300 μmol·m-2·s-1时,3种植物qP变化幅度均较小;当光强大于300 μmol·m-2·s-1时,3种植物qP均呈波动式降低趋势,且均随胁迫天数增加降低幅度逐渐增大,说明干旱胁迫可显著抑制3种植物叶片PSⅡ反应中心的开放比例和参与光化学反应的能量。同时,3种植物非光化学淬灭系数(NPQ)的光响应曲线变化趋势与Fv/Fm、Fv′/Fm′及qP相反;整个胁迫过程中白刺NPQ处于较高水平,梭梭、沙蒿呈上升趋势,说明白刺可通过增加热耗散来避免强光对光合系统的伤害,从而减轻自身在干旱胁迫下的光抑制;而在干旱胁迫30 d和18~30 d时,梭梭、沙蒿这种热耗散能力几乎消失,长时间干旱胁迫会引起两种植物光合电子传递受阻,并随着光强的增加,过剩能量以qP形式耗散,导致PSⅡ反应中心受损,表现出光抑制。
4 结 论
土壤自然干旱胁迫过程中,梭梭、白刺、沙蒿3种植物在光合生理特性方面均可通过合理协调碳同化和水分消耗之间的关系,调节叶片气孔、水分利用效率及叶绿素荧光参数等的变化,增强对干旱胁迫的耐受性及适应性,保证自身生理代谢活动的进行与正常生长,但其抗旱途径与机理方面存在差异,即不同植物对干旱胁迫的适应性不同,原因可能与3种植物的生长习性及其抗旱生理特点有关。例如,梭梭、白刺具有肉质叶片,干旱胁迫下不易失水,能最大限度地保持细胞的亲水能力,具有保持较高水平的抗旱生理特点;而沙蒿叶片表面虽有较厚的角质层,但其叶片面积较大,干旱胁迫条件下叶片蒸腾作用强烈,保水力相对较弱。据此,根据整个胁迫过程中植株的生长状态,结合隶属函数法对16项测定指标的综合评价,表明3种植物中梭梭的耐旱能力仅次于白刺,沙蒿耐旱能力稍弱。这一结果与3种植物的生长习性及光合生理特点相吻合。因此,在干旱荒漠区生态恢复过程中,对土壤干旱程度较重的造林地,应尽量首选白刺、梭梭作为先锋物种。
