游离脂肪酸对畜禽肠道内分泌细胞作用的研究进展
2021-11-09陈文静杨晓静
陈文静,杨晓静
(南京农业大学动物医学院,江苏南京 210095)
肠道是机体最重要的消化器官,也是与外界物质接触最为密切的部位。营养物质包括脂肪酸在胃肠道的消化与吸收影响着胃肠运动和分泌,进而影响机体代谢等生理过程。这些重要的生理功能都离不开肠道上皮层不同类型的细胞,如吸收性肠上皮细胞、杯状细胞、内分泌细胞、潘氏细胞和M 细胞等。其中,肠内分泌细胞(Enteroendocrine Cells,EECs)所分泌的多种激素参与调节肠道自身的运动和分泌功能、食欲和摄食行为、胃肠化学感应、营养物质代谢等过程[1]。因此,脂肪酸和肠道内分泌之间相互作用,在调控机体营养吸收和能量代谢等方面具有重要作用。本文综述了脂肪酸对畜禽肠道EECs 的作用,为其调控畜禽肠道健康、生长发育以及相关疾病的防治提供参考。
1 脂肪酸
脂肪酸是一类由碳、氢、氧3 种元素组成的化合物。不同链长的脂肪酸有不同的代谢途径,脂肪酸链长是脂肪消化和吸收的重要决定因素。根据碳链长度可将其分为短链脂肪酸(碳链上碳原子小于6,Short-Chain Fatty Acids,SCFAs)、中链脂肪酸(碳链上碳原子为6~12,Medium-Chain Fatty Acids,MCFAs)、长链脂肪酸(碳链上碳原子数大于12,Long-Chain Fatty Acids,LCFAs)。
1.1 SCFAs 及其在肠道的功能 SCFAs 又称为挥发性脂肪酸,主要包括乙酸、丙酸、丁酸、异丁酸和异戊酸等。大多数SCFAs 是由肠道微生物(如乳酸杆菌、双歧杆菌等)发酵分解不易消化的碳水化合物(包括抗性淀粉和可溶性低聚糖和多糖)而产生的,其他SCFAs 则是由饮食摄入和蛋白质等代谢产生[2]。肠道SCFAs 中乙酸、丙酸和丁酸含量较高,它们的比例约为3:1:1,占总SCFAs 的90%~95%,其余仅占很小一部分[3]。发酵产生的SCFAs 在体内参与不同组织的代谢,它们可被肠上皮细胞吸收并在线粒体内被氧化,为机体提供5%~10%的能量[4]。SCFAs 除了是一种瞬时能源外,还被认为是局部和远端生理功能中的关键信号分子,如跨上皮阴离子分泌、肠道运动和全身能量代谢等[5]。
SCFAs 主要通过抑制组蛋白去乙酰基酶(HDACs)和激活SCFAs 受体2 种方式对细胞产生影响进而促进肠道稳态。SCFAs 作为配体诱导HDACs 抑制作用,抑制NF-κB 途径,下调促炎细胞因子的表达,进而调控免疫反应[6]。目前已经发现的SCFAs 受体有FFAR2(也被称为GPR43)、FFAR3(GPR41)以及GPR109A。这些受体分布广泛,不仅大量分布于肠上皮细胞,在肠内分泌细胞、免疫细胞、肝细胞、脂肪细胞以及神经细胞中也都有分布[7]。
在SCFAs 中,丁酸在畜禽上的应用最为广泛,但是丁酸易挥发、有浓烈的酸臭味并且较难传输到肠道后段,因此目前生产上常用丁酸盐和丁酸甘油酯。Sun 等[8]研究表明,在生长育肥猪日粮中添加丁酸钠可降低耗料增重比,增加胴体重量并增加猪盲肠中拟杆菌的相对丰度,改善肠道健康。在经产母猪日粮中添加丁酸钠显著提高了仔猪出生窝重、新生仔猪体重和断奶体重[9]。而断奶仔猪日粮中添加丁酸钠显著增加了肠道绒毛高度和绒毛高度/隐窝深度比,抑制了肥大细胞活化和炎性介质产生[10],增强肠黏膜紧密连接蛋白的表达[11],增强空肠和结肠闭锁蛋白的表达,改善细菌群落[12],降低了断奶后腹泻发病率。在肉鸡日粮中添加丁酸盐可以促进肠道发育,诱导抗炎作用以及调节肉鸡肠道微生物群落[13-15],并有效控制了肠道中沙门氏菌的繁殖。刘丽华等[16]研究表明,在14 日龄肉仔鸡饲粮中添加不同丁酸钠可显著提高其采食量和体重,显著降低耗料增重比。
1.2 中、长链脂肪酸及其在肠道的功能 MCFAs 包括己酸(C6)、辛酸(C8)、葵酸(C10)和月桂酸(C12),被甘油脂化后形成中链甘油三酯(Medium Chain Triglycerides,MCT)。MCFAs 主要存在于椰子油和棕榈油等植物油中,在反刍动物的乳脂和椰子油中含量较高,但在黄油中MCFAs 只有不到6%,玉米油中不含任何MCFAs[17]。
LCFAs 又称为高级脂肪酸。在碳原子个数为14~24的LCFAs 中,LCFAs 的碳原子个数多为偶数,因为自然条件下脂肪酸的生物合成总是以2 个碳原子为扩增单位,如豆蔻酸(C14)、软脂酸(C16)、硬脂酸(C18)以及花生酸(C20)等饱和脂肪酸。LCFAs 可根据其碳链的饱和程度分为饱和脂肪酸、单不饱和脂肪酸以及多不饱和脂肪酸。根据不饱和脂肪酸双键所连碳原子的位置不同,LCFAs 又可分为n-3、n-6、n-9 等,包括亚麻酸(LA)、油酸、亚油酸(LOA)、花生四烯酸(AA)和二十二碳六烯酸(DHA)等。LCFAs 在机体的生长发育、保护肠黏膜屏障以及刺激肠道内分泌细胞分泌激素等方面起着重要作用。
MCFAs 和LCFAs 主要通过激活受体发挥作用,从而对细胞产生影响调节肠道稳态。不同研究都表明,FFAR1(GPR40)和FFAR4(GPR120)可同时被MCFAs 和LCFAs 激活[18],但是GPR84 只能被MCFAs激活[19]。FFAR1 和FFAR4 分布广泛,LCFAs 直接和间接诱导了胰岛素分泌证明FFAR1 和FFAR4 在肠道中也大量分布[20-21]。GPR84 在脑、结肠、肝脏和脾脏等都有分布[18]。
仔猪对MCFAs 的消化吸收率很好,除了供能作用,还可提高仔猪断奶后的消化率和饲料效率[22],缓解肠道损伤。研究表明,断奶前饲喂MCFAs 提高了仔猪的生长性能,促进了肠道发育[23],降低了弱仔猪的死亡风险[24]。日粮中添加鱼油(富含ω-3 多不饱和脂肪酸)能够抑制LPS 对断奶仔猪的刺激,缓解LPS 对仔猪的生长抑制[25]。在经产母猪日粮中添加鱼油和大豆油显著提高了哺乳仔猪的生长性能[26]。在肉鸡日粮中添加MCFAs 显著降低了腹部脂肪等,防止脂肪过度堆积,提高饲料利用率和胴体品质[27]。早期研究表明,饲粮中添加鱼油可以有效减少柔嫩艾美耳球虫对雏鸡增重的不利影响以及肠道病变的程度[28]。在肉仔鸡和蛋鸡日粮中添加多不饱和脂肪酸能一定程度改善肉品质[29],产出富含多不饱和脂肪酸的鸡蛋[30]。
2 EECs
EECs 是由肠道干细胞分化而来,是肠上皮细胞中重要的一员,虽然在整个胃肠腔上皮细胞中EECs 仅占不到1%,但是其数量超过了机体其他器官所有内分泌细胞的总和,并且分泌超过20 种激素,因此EECs 被认为是机体内最大、最复杂的内分泌器官[31]。EECs 散布在肠管黏膜的上皮组织中,大多为“开放型”,其顶端表面覆盖着一层微绒毛,与管腔成分直接接触,感受管腔食物刺激和肠道pH 变化等;还有很大比例的EECs 为“封闭型”,包括肠嗜铬细胞(EC 细胞)、肠嗜铬样细胞(ECL 细胞)、X/A 细胞和D 细胞,它们不直接接触管腔,通过旁分泌感受内容物等变化来调节其内分泌状态[32]。
2.1 EECs 的分类 EECs 通常根据其产生的主要激素来进行分类(表1)[33]。在十二指肠中,EECs 中的K 细胞分泌葡萄糖依赖性促胰岛素多肽(Glucose-Dependent Insulinotropic Polypeptide,GIP),具有抑制胃酸分泌、促进胰岛素释放的作用[34]。I 细胞分泌胆囊收缩素(Cholecystokinin,CCK),具有抑制采食、促进胰酶分泌和胆囊收缩的作用[35]。S 细胞分泌肠促胰液素(Secretin,SCT),可以抑制胃酸分泌、促进胰腺分泌[33]。位于小肠远端和结肠的L 细胞产生胰高血糖素样肽1 和2(GLP-1 和GLP-2),也产生肽YY(PYY),具有促进胰岛素分泌、抑制采食等作用[33]。EC 细胞分布在肠道各处,存储了胃肠道中90%以上的5-羟色胺(5-Hydroxytryptamine,5-HT),分泌的5-HT可以刺激肠道平滑肌收缩,调节胃肠道运动[36]。
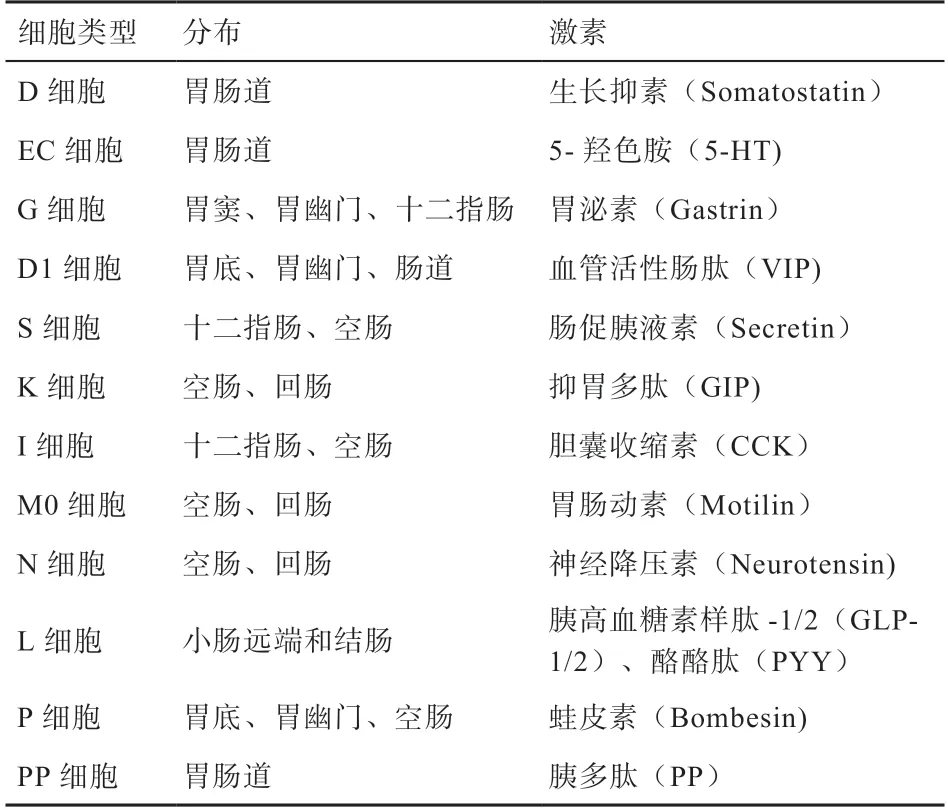
表1 主要肠道内分泌细胞及其分泌的激素[32]
2.2 肠内分泌细胞的分布 EECs 的分布存在物种间差异,比如小鼠的L 细胞主要分布于回肠和结肠,猪的L细胞主要分布于十二指肠[37],而鸡的L 细胞则沿着小肠分布[38]。在猪的肠黏膜中,S 细胞和I 细胞主要分布在十二指肠黏膜[39]。同时EECs 的分布还受到外界环境的影响,当饮食中含有高脂饲料时,小鼠肠道的L 细胞和K 细胞的数量均显著减少,且表达量也显著下降[40]。
EECs 释放的激素大多数通过内分泌的方式进入血液循环作用于靶细胞;有一些激素通过旁分泌进入细胞间隙,扩散到邻近靶细胞发挥作用;还有一部分释放的激素通过自分泌的方式作用于靶细胞膜上。EECs 释放的激素与靶细胞上的受体结合发挥相应的生理作用,如调节采食活动和胃酸分泌、调节胰岛素分泌、胆囊收缩和胃肠蠕动等。
3 脂肪酸对EECs 作用的影响
游离脂肪酸不仅是必需的营养元素,还参与调节许多细胞功能。游离脂肪酸通过EECs 表面的G 蛋白偶联受体(GPCRs)对细胞产生影响,进而影响肠道功能。许多研究表明,GPCRs 作为营养物质传感器,其在整个胃肠道中都有表达[41]。Symonds 等[41]研究表明,短链脂肪酸受体FFAR2 在结肠近端高表达,FFAR3 在空肠远端表达最强。中链脂肪酸受体GPR84 和中、长链脂肪酸受体FFAR1 在回肠中表达最高,而FFAR4 在结肠近端中表达水平最高[41]。这种分布与其生物学效应有关,中、长链脂肪酸来源于饮食,相比之下,肠道中SCFAs 主要由微生物发酵产生,因此,FFAR2 和FFAR3 在肠道前段表达较低,主要存在于肠道后段。
3.1 SCFAs 对EECs 分泌的影响 在SCFAs 中,FFAR2和FFAR3 都可以识别SCFAs,但其优先识别的SCFAs不同,FFAR2 优先识别乙酸和丙酸,而FFAR3 优先感应丙酸、丁酸和其他SCFAs[42]。肠腔内SCFAs 浓度增加可诱导L 细胞和EC 细胞增殖,乙酸和丙酸通过Gq/11 而不是Gi 增加细胞内钙水平,从而诱导L 细胞分泌GLP-1[5]。相反,在一项在体研究中,丁酸(而不是乙酸和丙酸)增加了野生型小鼠的GLP-1 和PYY水平,而在FFAR3-/-鼠中未观察到这一现象,并且丁酸和丙酸对K 细胞分泌GIP 的刺激作用也不依赖于FFAR3[43]。Moquet 等[44]研究表明,在幼龄肉仔鸡日粮中添加丁酸改变了消化过程和氨基酸的生物利用度,可能与肠道内分泌L 细胞分泌活动的改变有关。
3.2 LCFAs 对EECs 分泌的影响 研究表明,FFAR1 敲除鼠在灌胃LCFAs 后,CCK、GLP-1 和GIP 分泌减少[45]。在肠内分泌细胞K 细胞中,FFAR1 在小肠下部K 细胞中高表达,FFAR4 在小肠上部的K 细胞中高表达,当FFAR4 被敲除后,K 细胞的GIP 分泌量减少,FFAR4 在脂肪酸诱导的GIP 分泌中起着重要作用,有利于在高脂引发的肥胖中诱导胰岛素分泌维持正常血糖水平[34]。FFAR4 可以与α-亚麻酸(ALA)和二十二碳六烯酸(DHA)结合,使胞外Ca2+通过L 型通道进入刺激STC-1 细胞株释放CCK、GLP-1 和PYY[46]。虽然LCFAs 的这些作用与FFAR1 和FFAR4 的激活有着密切关系,但这些效应可能还涉及其他机制。有研究表明,ALA 仍然能刺激FFAR1/4 双敲除鼠建立的混合结肠上皮细胞分泌GLP-1[47]。ALA 和DHA 也可以刺激L细胞上的FFAR1 产生GLP-2,从而改善DSS 诱导的小鼠结肠炎[48]。
除了FFAR1 和FFAR4 外,CD36 和脂肪酸转运蛋白(FATP4)与LCFAs 直接结合作用,促进肠腔脂肪酸的吸收[45,49]。橄榄油灌胃后,与野生型小鼠相比,CD36 敲除小鼠CCK 和促胰液素分泌减少;在体外,脂肪酸促进表达CD36 的细胞释放促胰液素和CCK,对脂肪消化和内环境稳定具有重要意义[49]。
4 EECs 中脂肪酸受体下游信号通路
LCFAs 受体FFAR1 和FFAR4 均与Gq偶联,通过激活三磷酸肌醇受体和内质网Ca2+的释放,激活胞吐作用的其他相关信号分子[40]。SCFAs 受体FFAR2 和FFAR3 中,FFAR2 被证实可以通过Gq和Gi/o参与调节cAMP 途径,而FFAR3 对Gs具有选择性[50],cAMP 和下游效应蛋白Epac2 和PKA 都可能参与胞吐机制。
LCFAs 与FFAR1 和FFAR4 结合,使得FFAR1 和FFAR4 与Gq/11偶联,激活下游信号通路PLC-β,生成IP3 和DAG,IP3 调节细胞内钙信号,Ca2+分泌增加,而DAG 调节PKC,共同调控GLP-1 等激素的分泌。但是,FFAR1 的激活调节信号Ca2+并不是从细胞的钙库中释放的,而是通过激活TRPC3、膜去极化和电压依赖的Ca2+进入原代L 细胞中的[51]。有研究证实,EC细胞中存在α2-肾上腺素能受体下游的百日咳毒素敏感刺激机制(瞬时受体电位TRPC4 通道的激活)[52]。在STC-1 细胞中,LA 可激活STC-1 细胞的瞬时受体电位通道M5(Trpm5),干扰降低Trpm5的mRNA 表达后,LA 诱导的细胞内钙和CCK 分泌的增加明显减弱,FFAR4 通过Trpm5 诱导STC-1 细胞CCK 分泌[53]。在原代L 细胞和GLUtag 细胞中,TRPA1 激动剂使细胞内Ca2+浓度升高,促进GLP-1 的分泌,当把TRPA1敲除或用TRPA1 抑制剂后,这些作用都显著减弱[54]。FFAR2 优先与2~3 碳的SCFAs 结合,通过Gq-磷脂酰肌醇途径增加细胞内Ca2+浓度,也可通过G 蛋白亚基Gi/o途径减少细胞内cAMP,而FFAR3 只能通过Gi/o途径优先与剩下的SCFAs 结合[55]。
除了FFARs,CD36 在十二指肠和空肠绒毛细胞的刷状膜缘处表达,脂肪酸通过CD36 增加细胞内钙和cAMP,进而调节EECs 的分泌[57]。LCFA 与CD36 结合后引起Src-PTKs 的磷酸化作用,激活PLC 途径后产生IP3,随后Ca2+从内质网中释放出来;同时钙池调控钙离子通道(SOC)打开,细胞外的Ca2+内流。Ca2+浓度增加和cAMP 的增加促进了EECs 的分泌。CD36作为LCFAs 的转运载体,还可能作为GPR120 的共受体,诱使和转运LCFA 至低亲和力受体GPR120[58]。
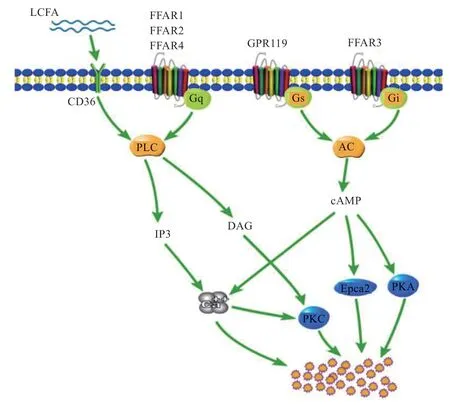
图1 肠内分泌细胞感应脂肪酸机制[56]
5 小 结
游离脂肪酸与EECs 的相互作用形成了稳定的肠道环境。游离脂肪酸作为饲料添加剂常用于提高肉鸡和仔猪的生长性能,但作用方式还不完全清楚。长链脂肪酸受体FFAR1 和FFAR4、短链脂肪酸受体FFAR2和FFAR3 都分布于肠内分泌细胞上,LCFAs 和SCFAs通过激活这些受体刺激EECs 分泌胃肠激素(CCK、GLP-1、5-HT 和GIP 等)。这些激素通过肠道作用于全身,如EC 细胞分泌的5-HT 可以刺激肠道平滑肌收缩,也可以通过脑-肠轴调节机体大脑活动;L 细胞分泌的GLP-1 在调节胰岛素分泌中起着重要作用。因此,了解脂肪酸对畜禽EECs 的作用,深入研究信号途径,有利于全面了解肠道消化吸收功能的过程。通过调整饲粮的脂肪酸成分,调控胃肠道化学感受器,优化肠道激素的分泌,刺激食欲和调节采食量,对改善畜禽肠道健康、调控生长发育具有重要意义。
